
Abstract
Background:
Oral tongue squamous cell carcinoma (OTSCC) is a common type of oral tumor. LncRNAs (long noncoding RNAs) and miRNAs (microRNAs) were identified as regulators in many human cancers. This study aims to explore the molecular basis of HOXA transcript at the distal tip (HOTTIP) in regulating OTSCC progression.
Materials and Methods:
The expression of HOTTIP, miR-124-3p, and high-mobility group AT-hook 2 (HMGA2) was detected by quantitative real-time polymerase chain reaction. Next, the proliferation was evaluated by 3-(4,5-dimethylthiazole-2-y1)-2,5-biphenyl tetrazolium bromide (MTT) assay. The migration and invasion were assessed by transwell assay. Furthermore, dual-luciferase reporter assay was performed to confirm the combination between HOTTIP and miR-124-3p, miR-124-3p, and HMGA2. Protein levels of HMGA2, β-catenin, c-Myc, and E-cadherin were examined by Western blot. The nude mice model was employed to test the tumor growth in vivo.
Results:
HOTTIP was upregulated in OTSCC tissues and cells, and was highly expressed in positive lymph node metastasis and late-stage OTSCC patients. Silencing HOTTIP impeded proliferation, migration, and invasion of OTSCC cells. Moreover, HOTTIP knockdown inhibited proliferation, migration, and invasion of OTSCC cells by targeting miR-124-3p. Besides, miR-124-3p targeted HMGA2 to block proliferation, migration, and invasion. HMGA2 could rescue the inhibitory effects of HOTTIP interference on proliferation, migration, and invasion. In addition, HMGA2 overexpression reversed the downregulation of β-catenin and c-Myc protein levels and upregulation of E-cadherin level affected by HOTTIP silencing. Finally, HOTTIP silencing repressed tumor growth and resulted in a great rise on miR-124-3p and E-cadherin expression and a distinct fall on HMGA2, β-catenin, and c-Myc protein levels.
Conclusions:
HOTTIP knockdown restrained proliferation, migration, and invasion of OTSCC cells by miR-124-3p/HMGA2 axis through Wnt/β-catenin pathway.
Introduction
Oral tongue squamous cell carcinoma (OTSCC) is the main type of oral squamous cell carcinoma, ranking sixth among the most common cancer types in the world. 1 OTSCC frequently occurred in young people, especially in male, 2,3 and has an increasing trend of incidence in recent years. 4 Despite great improvement in therapeutic strategies for OTSCC, the 5-year survival rate has not been obviously improved. 5 OTSCC has been widely reported to feature with the easy recurrence, metastasis, and poor prognosis because of the rapid cell migration and invasion. 6 Metastasis is also the main reason for the high mortality among OTSCC patients. 7 Therefore, how to regulate distant metastasis and tumor growth is the key theme for the treatment of OTSCC patients.
Long noncoding RNAs (lncRNAs), by definition, are a class of noncoding RNAs with length of >200 nucleotides. They have no protein-coding capacity but act as important regulators in various biological and physiological processes such as genes transcription, epigenetic modification, and cell proliferation. HOXA transcript at the distal tip (HOTTIP) was found to locate at 5′ tip of the HOXA gene cluster in chromosome 7p15.2 and played a carcinogenicity role in varieties of human cancers, including OTSCC. 8 –10 Emerging study has demonstrated that HOTTIP knockdown inhibits the growth and invasion of OTSCC cells, suggesting that it may serve as a potential target for OTSCC therapy. 11 Also, the level of HOTTIP was upregulated in TSCC and HOTTIP might be a novel marker of poor prognosis, which implied that HOTTIP could be a therapeutic target for TSCC. Although the function of HOTTIP in the progression of OTSCC cells has been reported, the regulatory network of HOTTIP in OTSCC is still unknown.
MicroRNAs (miRNAs), a kind of small noncoding RNAs with ∼22 nts in length, suppress the level of target mRNAs through base complementary matching principle. Over the past decades, numerous miRNAs have been discovered to involve in diagnosis and therapy of OTSCC. 12 –14 MiR-124-3p, the mature form of human miR-124 precursor, functioned as a vital regulator in many cancers, including OTSCC. Zhang et al. indicated that miR-124 was involved in the regulation of epithelial–mesenchymal transition in TSCC cells together with UCA1/JAG1 axis. 15 However, little research has been done on the mechanism of miR-124-3p regulating the proliferation, migration, and invasion of OTSCC cells.
High-mobility group AT-hook 2 (HMGA2) has been reported to associate with carcinogenesis, in particular metastasis. 16,17 HMGA2 expression was sharply elevated in many cancers, including breast cancer, 18 lung cancer, 19 colorectal cancer, 20 and oral carcinomas. 21 Moreover, Zhao et al. reported that HMGA2 promoted invasion and metastasis of TSCC cells, and could be used as a biomarker for prognosis and therapy of TSCC. 22 Also, Zhang et al. determined that HMGA2 promoted malignant transformation and was an independent prognosis marker for TSCC. 23 Even though plenty of studies have indicated the function of HMGA2 in TSCC, the role of HMGA2 and the molecular mechanism of HOTTIP/miR-124-3p/HMGA2 axis in OTSCC need to be further explored.
In this study, we tested the expression of HOTTIP in OTSCC tissues and cells, and evaluated its effects on proliferation, migration, and invasion of OTSCC cells. Besides, we explored the interaction among HOTTIP, miR-124-3p, and HMGA2. These findings may provide new evidence for better understanding of HOTTIP, and it may be a potential biomarker for diagnosis of OTSCC.
Materials and Methods
Patients and cell culture
OTSCC tumor tissues and adjacent normal tissues were obtained from 60 OTSCC patients, who had enrolled at our hospital in 2012–2017. The tissue samples were immediately stored in liquid nitrogen. None of the patients received any treatment before the samples were collected. A written informed consent was provided by each patient before surgery. All methods and experiments were authorized by the Ethics Committee of Central Hospital of Jingzhou, the Second Clinical Medical College, Yangtze University.
Normal Oral Keratinocytes-Spontaneously Immortalized (NOK-SI) was provided by Dr. Silvio Gutkind (National Institutes of Health, Bethesda, Rockville, MD). OTSCC cell lines (SCC9, CAL27, and SCC25) were purchased from American Tissue Culture Collection (ATCC, Manassas, VA). The UM1 cells were acquired from Japanese Collection of Research Bioresources (JCRB) cell bank (Osaka, Japan). All the cells were incubated in Dulbecco's modified Eagle's medium (Gibco, Carlsbad, CA) supplemented with 10% fetal bovine serum (Invitrogen, Carlsbad, CA) at 37°C, 5% CO2 air atmosphere.
Cell transfection
Small-interfering RNA against HOTTIP (si-HOTTIP#1 and si-HOTTIP#2) and their corresponding negative control (si-NC), short hairpin RNA against HOTTIP (sh-HOTTIP) and its control (sh-NC), the overexpression vector of HOTTIP and miR-124-3p (HOTTIP and miR-124-3p mimics) and their negative controls (pcDNA and mimics-NC), miR-124-3p inhibitor and its matching control (inhibitor-NC) were all purchased from RiboBio (Guangzhou, China). HMGA2 overexpression vector (HMGA2) and its control were synthesized from Sangon Biotech (Sangon, Shanghai, China). The oligonucleotides or vectors were transfected into SCC25 and UM1 cells by Lipofectamine 2000 transfection reagent (Invitrogen).
Quantitative real-time polymerase chain reaction
Total RNA extraction was conducted by TRIzol reagent (Invitrogen) referring to the user manual. The quality and quantity of total RNA were assessed by NanoDrop 2000 (Thermo Fisher Scientific, Rockford, IL). For the lncRNA HOTTIP and HMGA2 mRNA, the reverse transcription was conducted by using EvoScript Universal cDNA Master (Roche, Basel, Switzerland). For miR-124-3p, the strand of cDNA was conducted by miRNA reverse transcription kit (Funeng, Guangzhou, China). Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using SYER Green (Solarbio, Shanghai, China). HOTTIP, HMGA2, and miR-124-3p were normalized by glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and U6 small RNA. The relative expression was calculated using 2−ΔΔCt method. The primers for HOTTIP, miR-124-3p, and HMGA2 were synthesized from Sangon Biotech (Sangon), and the primer sequences were listed as follows: HOTTIP (F, 5′-AAGGAGGAAACGCCTTCTCG-3′; R, 5′-CATCAAGCTAGGGTGGGGTG-3′) miR-124-3p (F, 5′-CGTAAGGCACGCGGTGAA-3′; R, 5′-AGTGCAGGGTCCGAGGTATT-3′), HMGA2 (F, 5′-ACCAACCGGTGAGCCCTCTC-3′; R, 5′-TTGAGCTGCTTTAGAGGGACTCTTG-3′), GAPDH (F, 5′-ATTCCATGGCACCGTCAAGGCTGA-3′; R, 5′-TTCTCCATGGTGGTGAAGACGCCA-3′), U6 (F, 5′-GTAGATACTGCAGTACG-3′; R 5′-ATCGCATGACGTACCTGAGC-3′).
Cell proliferation assay
Cells were inoculated into 96-well plates and cultured in an incubator at 37°C, 5% CO2 for 24 h. After transfection with various vectors, cells were collected at 24, 48, and 72 h. Then, 3-(4,5-dimethylthiazole-2-y1)-2,5-biphenyl tetrazolium bromide (MTT) solution (0.5 mg/mL; Sigma-Aldrich, Louis, MO) was added to cells and cultured for 4 h. Next, an orange colored product (formazan) was observed and dissolved by using dimethyl sulfoxide. The absorbance of cells was observed under a microplate reader (Thermo Fisher Scientific) at 490 nm.
Transwell assay
For cell invasion, the transwell upper chambers (Corning, New York, NY) were coated by Matrigel (BD, Franklin Lakes, NJ), and then OTSCC cells were cultured into the upper chambers. Next, the medium containing serum was added to the lower chambers. While for cell migration, the transfected cells were directly seeded into the upper chamber without any treatment. The chambers were incubated at 37°C. After 24 h, cells that pass through the membranes were collected and stained with 0.5% crystal violet (Sigma) for 5 min. Finally, cell number was evaluated by the Countess automatic cell counter (Invitrogen) with at least three random fields.
Dual-luciferase reporter assay
The putative binding sites between HOTTIP and miR-124-3p were predicted by miRcode tool, and the potential target of miR-124-3p (HMGA2) was verified by starBase2.0 website tool. The wild type of HOTTIP and HMGA2 3′-UTR (WT-HOTTIP and HMGA2 3′-UTR-WT) was synthesized by Tongyong (Shanghai, China). At the same time, the mutant sequences of HOTTIP and HMGA2 3′-UTR (MUT-HOTTIP and HMGA2 3′-UTR-MUT) were designed and synthesized by Sangon. Next, MUT-HOTTIP and HMGA2 3′-UTR were cloned into the luciferase vector. Subsequently, all luciferase vectors along with miR-124-3p mimics or mimics-NC were transfected into SCC25 and UM1 cells. After cultivation in the incubator for 48 h, the activities of luciferase were monitored by the Dual-Luciferase Reporter Assay Kit (Genomeditech, Shanghai, China).
Western blot assay
Proteins were obtained from transfected cells by RIPA Buffer (Beyotime, Shanghai, China). Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gel was freshly prepared to separate proteins according to the molecular weight of different proteins. After electrophoresis for 2 h, proteins were electrotransferred onto polyvinylidene difluoride membranes (Millipore, Bedford, MA). After being blocked in 5% nonfat milk for 2 h, the membranes were incubated with primary antibodies against HMGA2 (1/1000, ab97276; Abcam, Cambridge, MA), β-catenin (1/5000, ab32572; Abcam), c-Myc (1/1000, ab32072; Abcam), E-cadherin (1/50, ab1416; Abcam), or GAPDH (1/2000, ab37168; Abcam) at 4°C. The next day, the membranes were interacted with horseradish peroxidase-conjugated secondary antibody (1/5000, ab6721; Abcam) for another 2 h. The protein signals were observed by ECL reagent (R&S, Shanghai, China) and analyzed using ImageJ analysis software.
Murine xenograft model
Nude mice (n = 6) were purchased from Tengxin Company (Chongqing, China) and randomly divided into two groups (n = 3). All the experiments on the mice model were permitted by the Animal Research Committee of Central Hospital of Jingzhou, the Second Clinical Medical College, Yangtze University, and the methods were in line with the guidelines of the National Animal Care and Ethics Institution. One group was infected with lentivirus harboring short hairpin (sh-NC and sh-HOTTIP). Tumor volume was measured every 3 d starting seventh day after inoculation and figured by the formula: 1/2 × width 2 × length. Tumor tissues were cut off from mice and weighted after cultured for 28 d. The tumors were stored at −80°C for further molecular experiments.
Statistical analysis
GraphPad Prism 7 software (GraphPad, Inc., La Jolla, CA) was used to analyze the data from qRT-PCR, Western blot, and luciferase assays. All data were repeated three times and presented as the mean ± standard deviation. Student's t-test and one-way analysis of variance were performed to evaluate the differences between groups. p-Value <0.05 was considered to be significant.
Results
LncRNA HOTTIP was highly expressed in OTSCC tissues and cells
To begin with, qRT-PCR was conducted to detect the expression of HOTTIP in OTSCC patients. Compared with the normal tissues, the expression of HOTTIP was largely increased in tumor tissues (Fig. 1A). Also, higher expression of HOTTIP was found in patients with positive lymph node metastasis than in negative patients (Fig. 1B). To clarify the role of HOTTIP in the development of OTSCC, the patients were divided into two groups (I + II and III + IV) according to the classification of clinical stage. qRT-PCR assay showed that HOTTIP level was significantly enhanced in stage III + IV relative to stage I + II (Fig. 1C). Furthermore, patients with high HOTTIP expression survived shorter than patients with low HOTTIP expression (p = 0.0096), suggesting that HOTTIP was closely associated with overall survival rate of OTSCC patients (Fig. 1D). In addition, HOTTIP expression was detected in normal (NOK-SI) and OTSCC cell lines (SCC9, CAL27, SCC25, and UM1), and the results indicated that HOTTIP level was prominently elevated in OTSCC cells (Fig. 1E). These data revealed that HOTTIP was involved in OTSCC development.
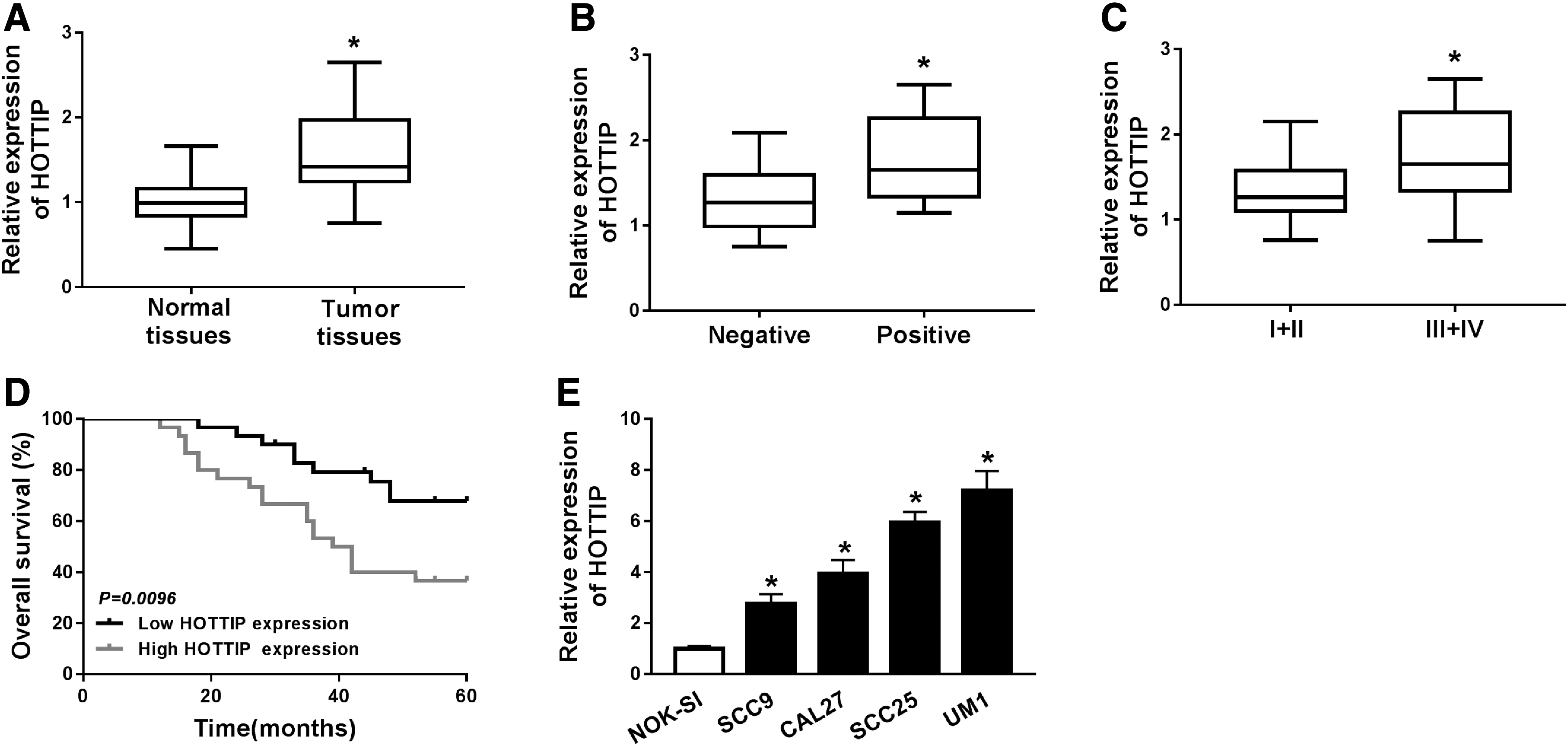
LncRNA HOTTIP expression in OTSCC tissue samples and cells and its correlation with clinical parameters.
Knockdown of HOTTIP inhibited proliferation, migration, and invasion of OTSCC cells
To explore the function of HOTTIP in OTSCC, si-HOTTIP#1 or si-HOTTIP#2 were transfected into SCC25 and UM1 cells. qRT-PCR data showed that HOTTIP expression was significantly decreased in both SCC25 and UM1 cells by transfection with HOTTIP knockdown (Fig. 2A, B). Moreover, the proliferation of SCC25 and UM1 cells was markedly inhibited when transfected with si-HOTTIP#1 or si-HOTTIP#2 (Fig. 2C, D). Meanwhile, silencing HOTTIP drastically hindered migration and invasion of SCC25 and UM1 cells (Fig. 2E–H). Due to the better suppression effect on HOTTIP level, si-HOTTIP#2 was selected for further experiments in our study. All the above data implied that HOTTIP knockdown restrained progression of OTSCC cells.

HOTTIP knockdown inhibited proliferation, migration, and invasion of OTSCC cells.
HOTTIP as a molecular sponge antagonized miR-124-3p availability on proliferation, migration, and invasion of OTSCC cells
To explore the network of HOTTIP in OTSCC, bioinformatics analysis tool (MiRcode) was used to predict the downstream target of HOTTIP. Interestingly, miR-124-3p has the potential binding sites with HOTTIP (Fig. 3A). To confirm the prediction, the mutant type of HOTTIP was designed and cotransfected with miR-124-3p mimics or mimics-NC into SCC25 and UM1 cells. Luciferase assay showed that miR-124-3p overexpression limited the luciferase activities of SCC25 and UM1 cells in WT-HOTTIP group compared with mimics-NC, while no changes of activities were observed in MUT-HOTTIP group (Fig. 3B, C). More interestingly, miR-124-3p expression was remarkably repressed in tumor tissues (Fig. 3D), and there is a negative correlation between HOTTIP and miR-124-3p expression in OTSCC (Fig. 3E). Further, miR-124-3p expression was measured by qRT-PCR in OTSCC cells, and we found that miR-124-3p level was decreased in SCC25 and UM1 cells (Fig. 3F). The expression of miR-124-3p was apparently promoted by downregulation of HOTTIP, and suppressed by overexpression of HOTTIP in SCC25 and UM1 cells (Fig. 3G). MTT assay determined that the proliferation of SCC25 and UM1 cells was greatly impeded by the interference of HOTTIP, which was abated by the introduction of miR-124-3p inhibition (Fig. 3H, I). Also, transwell assay indicated that migration and invasion of SCC25 and UM1 cells were largely blocked by HOTTIP silencing, while miR-124-3p inhibitor attenuated the effects on cell migration and invasion caused by HOTTIP knockdown (Fig. 3J, K). Totally, HOTTIP targeted miR-124-3p and downregulated miR-124-3p expression, and silencing HOTTIP hindered proliferation, migration, and invasion of OTSCC cells.
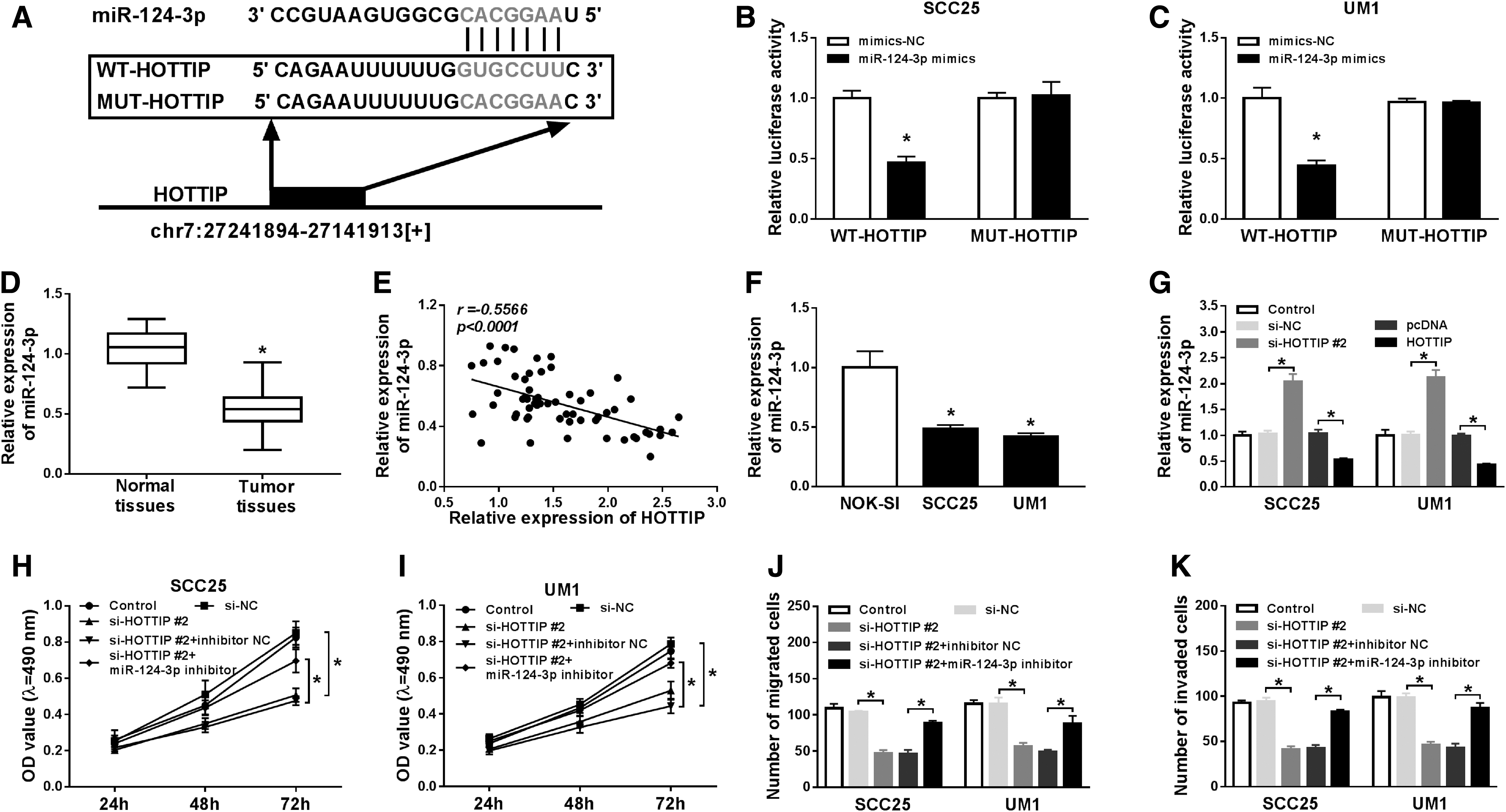
HOTTIP targeted miR-124-3p to regulate proliferation, migration, and invasion of OTSCC cells.
MiR-124-3p was directly interacted with HMGA2 and regulated proliferation, migration, and invasion of OTSCC cells through HMGA2
To illuminate the regulatory mechanism of miR-124-3p in OTSCC, starBase2.0 tool was used to predict its potential targets. The putative binding sites between miR-124-3p and HMGA2 3′-UTR and mutant sequences of HMGA2 3′-UTR-MUT are shown in Figure 4A. Furthermore, this prediction was verified by dual-luciferase reporter assay. Compared with mimics negative control group, luciferase activities of SCC25 and UM1 cells transfected with miR-124-3p upregulation were strikingly inhibited in HMGA2 3′-UTR-WT group, whereas in HMGA2 3′-UTR-MUT group luciferase activities remain unchanged (Fig. 4B, C). Besides, we detected the expression of HMGA2 mRNA and protein by qRT-PCR, and Western blot assay in SCC25 and UM1 cells transfected with miR-124-3p mimics or miR-124-3p inhibitor. The results demonstrated that both mRNA and protein levels of HGMA2 were downregulated by miR-124-3p overexpression and obviously enhanced by transfection with miR-124-3p inhibition (Fig. 4D, E). Notably, the mRNA level of HMGA2 was drastically increased in OTSCC tumor tissues (Fig. 4F), and had a negative correlation with miR-124-3p expression (Fig. 4G). Also, HMGA2 mRNA and protein were evidently promoted in SCC25 and UM1 cells compared with NOK-SI cell line (Fig. 4H, I). Subsequently, MTT and transwell assays were performed to assess the interaction between miR-124-3p and HMGA2 on cell proliferation, migration, and invasion. The results indicated that miR-124-3p upregulation distinctly restrained proliferation of SCC25 and UM1 cells, while HMGA2 overexpression rescued the inhibitory effect on cell proliferation by miR-124-3p (Fig. 4J, K). Similarly, cell migration and invasion were inhibited by miR-124-3p upregulation, which were all abrogated by the overexpression of HMGA2 (Fig. 4L, M). Collectively, HMGA2 was a target of miR-124-3p, and HMGA2 overexpression could weaken miR-124-3p upregulation-mediated inhibitory effect on proliferation, migration, and invasion of OTSCC cells.
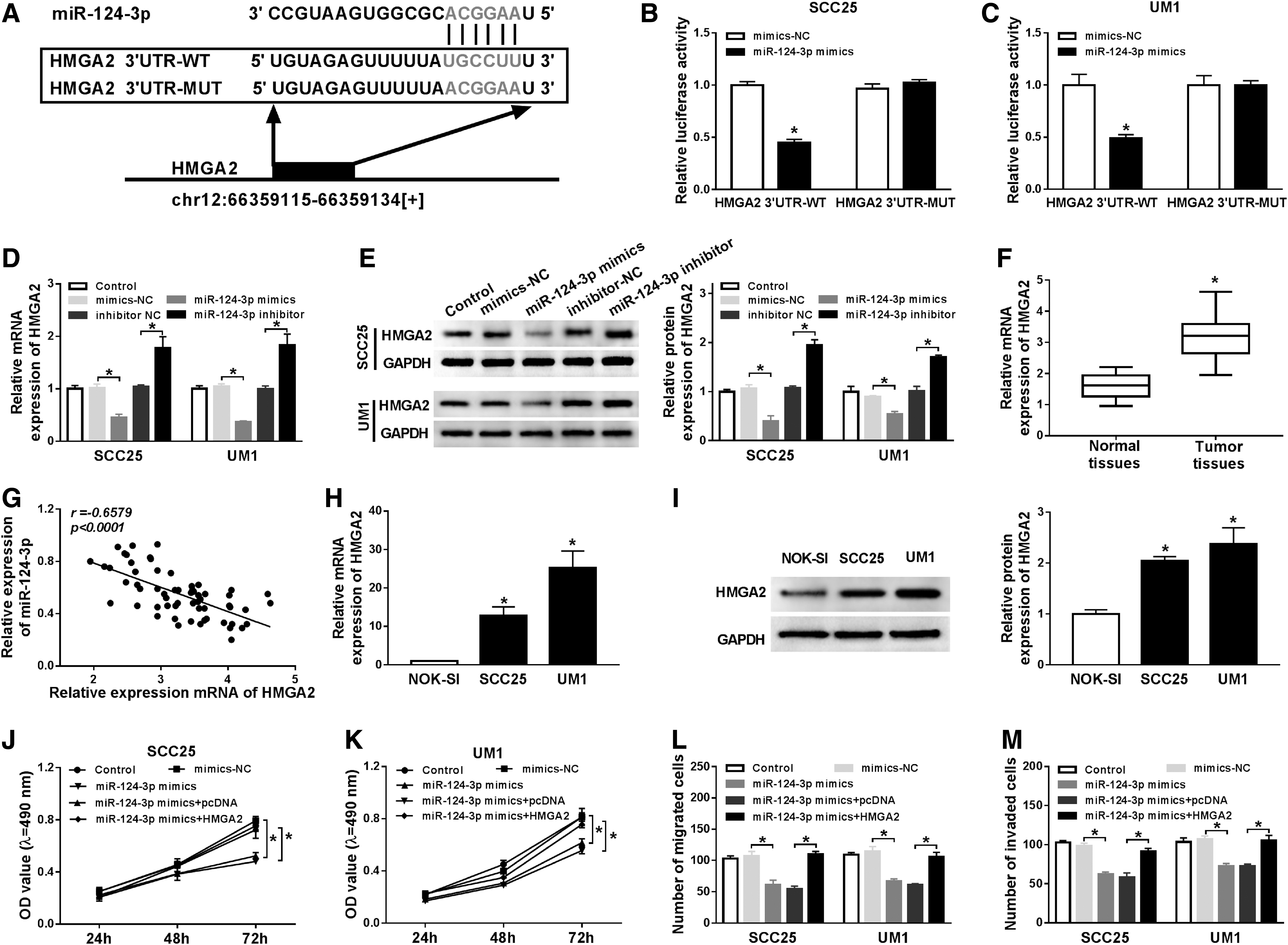
MiR-124-3p interacted with HMGA2 and retarded proliferation, migration, and invasion of OTSCC cells through HMGA2.
HOTTIP targeted miR-124-3p to regulate HMGA2-mediated Wnt/β-catenin pathway
To clarify the relationship among HOTTIP, miR-124-3p, and HMGA2 in OTSCC, qRT-PCR and Western blot assays were conducted to detect HMGA2 expression in OTSCC cells. Compared with the negative control, HOTTIP interference largely reduced HMGA2 mRNA and protein expression in SCC25 and UM1 cells, while the introduction of miR-124-3p inhibition resulted in an obvious increase in HMGA2 expression (Fig. 5A, B). Interestingly, HOTTIP expression was positively correlated with HMGA2 expression (Fig. 5C). Next, cell proliferation was examined by MTT assay, and results exhibited that silenced HOTTIP significantly retarded proliferation of SCC25 and UM1 cells. After cotransfection with HMGA2, however, cell proliferation was partially recovered (Fig. 5D, E). Transwell assay disclosed that HMGA2 mitigated HOTTIP knockdown-mediated antimigration and anti-invasion effect on SCC25 and UM1 cells (Fig. 5F, G). In addition, levels of Wnt/β-catenin pathway-related proteins β-catenin, c-Myc, and E-cadherin were measured by Western blot. The results revealed that β-catenin and c-Myc expression were constrained and E-cadherin was enhanced by HOTTIP silencing, which were all reversed by HMGA2 overexpression (Fig. 5H, I). In total, HOTTIP knockdown hindered proliferation, migration, and invasion through HMGA2-mediated Wnt/β-catenin pathway.
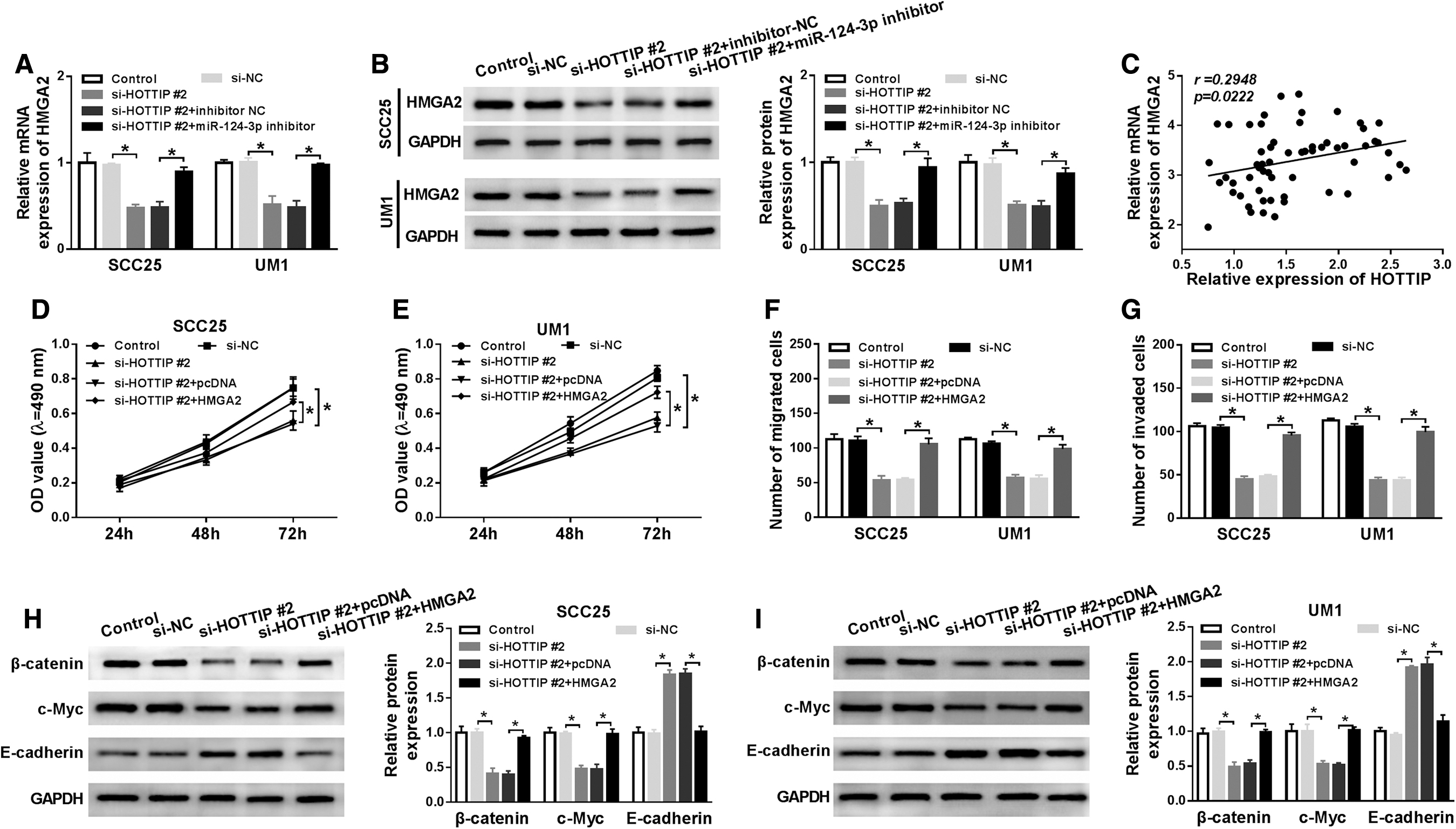
HOTTIP targeted miR-124-3p to regulate HMGA2-mediated Wnt/β-catenin pathway.
Knockdown of HOTTIP inhibited tumor growth in vivo
To further confirm the tumorigenicity of HOTTIP in vivo, six nude mice were injected with transfected cells. At 28 d postinoculation, tumor tissues were cut off for the measurement of volume and weight (Fig. 6A). The volume of tumor was apparently reduced in sh-HOTTIP group compared with those in control group (Fig. 6B). Tumor weight of sh-HOTTIP group was also lighter than that of sh-NC group (Fig. 6C). Furthermore, the tumors were collected for molecular analysis. The levels of HOTTIP and HMGA2 were prominently decreased, and miR-124-3p expression was increased by the transfection with HOTTIP downregulation (Fig. 6D). In addition, the abundance of HMGA2, β-catenin, and c-Myc was strikingly inhibited, and E-cadherin protein level was promoted in tumor tissues transfected with sh-HOTTIP (Fig. 6E). In a word, the interference of HOTTIP blocked tumor growth by suppressing Wnt/β-catenin pathway in vivo.

HOTTIP silencing blocked tumor growth of OTSCC mice through Wnt/β-catenin pathway.
Discussion
OTSCC is a common malignancy of the oral cavity and has been a great threat to the public health. Increasing studies manifested that lncRNAs and miRNAs were the important regulators in the various processes of cancers, such as tumorigenesis, cell proliferation, and autophagy. However, the underlying mechanism remains largely unknown. Our study illuminated a new molecular mechanism of HOTTIP in the development of OTSCC.
Burgess reported that HOTTIP elevated the expression of multiple 5′HOXA genes in many cancers. 24 In TSCC, HOTTIP has the higher expression in TSCC tissue, and its expression was closely related to the overall survival of TSCC patients. 25 In addition, HOTTIP was highly expressed in OTSCC, and its suppression inhibited the proliferation of OTSCC cells. 11 Consistent with above studies, we investigated the expression in OTSCC. The data showed that HOTTIP level was markedly increased in OTSCC tumor tissues, and the higher level of HOTTIP was also found in the positive lymph node metastasis and late-stage patients. Moreover, HOTTIP knockdown blocked the proliferation, migration, and invasion of OTSCC cells. All these data further confirmed the expression and function of HOTTIP in OTSCC, and manifested that HOTTIP might play an oncogenic role in OTSCC.
LncRNAs function as competing endogenous RNAs to regulate miRNAs biological function was extensively reported in recent years. In our study, bioinformatics analysis was performed, and the results disclosed that HOTTIP directly bind to miR-124-3p. Although Zhang et al. have demonstrated that miR-124-3p expression was declined in tongue cancer tissues and miR-124-3p modulated epithelial–mesenchymal transition (EMT) and progression of tongue cancer with lncRNA UCA1, 15 the interaction between HOTTIP and miR-124-3p remains unexplored. We conducted luciferase assay to confirm the combination between them. Furthermore, the expression of miR-124-3p was largely decreased in OTSCC tissues and had a negative correlation with HOTTIP level. Not only that, the suppression of miR-124-3p rescued the inhibition of OTSCC proliferation, migration, and invasion in vitro induced by HOTTIP knockdown. These data suggested that HOTTIP directly bound to miR-124-3p and regulated the progression of OTSCC cells by sponging miR-124-3p.
A previous study showed that HMGA2, a key regulator of tumor metastasis, was upregulated in TSCC tissues and cells. 26 In agreement with this finding, the data in our study showed that HMGA2 expression was promoted in OTSCC tissues, and there was an opposite trend in miR-124-3p expression. Interestingly, HMGA2 was predicted as a target of miR-124-3p, and dual-luciferase reporter assay verified this prediction. Moreover, HMGA2 could recover the inhibitory effect mediated by miR-124-3p on cell proliferation, migration, and invasion upregulation.
Klaus and Birchmeier implicated that Wnt signaling pathway played a vital role in tumor initiation, 27 and Kodai et al. reported that lymph node metastasis was an important predictor of patients' survival. 28 To elucidate the interaction among HOTTIP, Wnt signaling, and lymph node metastasis in OTSCC, proteins related to Wnt/β-catenin signaling pathway and lymph node metastasis were also detected in our study. The results determined that HOTTIP silencing drastically hindered the expression of β-catenin and c-Myc and enhanced E-cadherin level, which were all relieved by the overexpression of HGMA2. Hence, these data indicated that HOTTIP interference inhibited the Wnt/β-catenin signaling pathway and lymph node metastasis of OTSCC by regulating HMGA2 expression. The in vivo experiment indicated the role of HOTTIP in the tumor growth in OTSCC, and the levels of miR-124-3p, HMGA2, β-catenin, c-Myc, and E-cadherin were all regulated by HOTTIP. Collectively, HOTTIP inhibited proliferation, migration, and invasion by miR-124-3p/HMGA2 axis through Wnt/β-catenin signaling pathway in OTSCC.
Conclusions
HOTTIP was upregulated in OTSCC tissues and cells, and HOTTIP knockdown blocked proliferation, migration, and invasion of OTSCC cells. HOTTIP bound to miR-124-3p and downregulated miR-124-3p expression. HOTTIP silencing inhibited proliferation, migration, and invasion through sponging miR-124-3p. Also, HMGA2 was a target of miR-124-3p, and its expression was negatively regulated by miR-124-3p. HGMA2 abolished the inhibition effects induced by miR-124-3p on proliferation, migration, and invasion in OTSCC cells. Furthermore, there was an antagonism between silenced HOTTIP and HMGA2 overexpression on proliferation, migration, invasion, Wnt signaling and lymph node metastasis. Besides, short hairpin HOTTIP remarkably reduced OTSCC tumor growth in vivo. Our results first demonstrated the functional effects and molecular mechanism of HOTTIP/miR-124-3p/HGMA2 axis in the development of OTSCC, which might provide a new light for the diagnosis and therapy of OTSCC.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.