
Abstract
Background:
miR-664b-5p accelerates the development of certain cancers, but the role of miR-664b-5p in hepatocellular carcinoma (HCC) has been less reported. Therefore, the authors aimed to study the role of miR-664b-5p in HCC progression.
Materials and Methods:
miR-664b-5p expression in liver cancer and adjacent tissues, and in HepG2 and SUN-475 cells, was measured by quantitative real-time polymerase chain reaction (qRT-PCR). Relationship between miR-664b-5p and AKT2 was predicted by TargetScan and confirmed by dual-luciferase reporter assay, and gene or protein expressions were determined by performing qRT-PCR and Western blotting. The viability and apoptosis, and the migration and invasion of HepG2 and SUN-475 cells were determined by CCK-8 assay and flow cytometry, and transwell assay, respectively.
Results:
Downregulated miR-664b-5p was observed in hepatocellular cancer tissues. Functional analyses revealed that miR-664b-5p mimic suppressed viability, migration, and invasion, but promoted apoptosis in HepG2 and SUN-475 cells. AKT2 was a target of miR-664b-5p, whose mimics inhibited the expression of AKT2. However, upregulated AKT2 promoted viability, migration, and invasion, but inhibited apoptosis in HepG2 and SUN-475 cells, and such effects were reversed by miR-664b-5p mimics.
Conclusions:
miR-664b-5p acts as a cancer suppressor through negatively regulating AKT2 expression in HepG2 and SUN-475 cells, suggesting that miR-664b-5p could be a protective target for HCC patients.
Introduction
Primary liver cancer, one of the most common malignant tumors, is the third cause of cancer-related death all over the world. 1 Hepatocellular carcinoma (HCC) accounting for 85%–90% of primary liver cancer is a main type of primary liver tumor, and 700,000 cases of HCC are diagnosed annually in the world, 2,3 with more than half of the cases occurring in China. 4 Due to a long subclinical period, the onset and progression of HCC are recessive and often difficult to be detected, but desired prognosis of HCC is particularly relevant to its early detection. 5 However, patients suffering from HCC are often diagnosed at an advanced stage, and treatment at that stage is limitedly available. 6 Furthermore, HCC progression is a complex process, with complex gene networks and changes in signaling pathways, many of them still need to be illuminated. 7 Therefore, it is highly important to understand the potential therapeutic target and molecular mechanism of HCC treatment.
MicroRNAs (miRNAs) are a class of small RNAs of about 22 nucleotides, and exert their effects in posttranscriptional regulation and different biological processes. 8 Accumulated evidence demonstrated that miRNAs play a pivotal role in the development of cancers. 9 For example, Zhang et al. reported that downregulated miR-664 is related to lower overall survival in cervical cancer 10 ; Ding et al. showed that downregulated miR-664 promotes cutaneous malignant melanoma proliferation 11 ; moreover, Song et al. found that miR-664b-5p plays an important role in the progression of breast cancer. 12 However, studies conducted on the effect of miR-664b-5p on HCC development are limitedly available.
Phosphatidylinositol 3-kinase (PI3K)/AKT signaling pathway plays an important role in cancer progression, 13,14 and among the many signal molecules of PI3K/AKT signaling pathway, AKT was the main signal molecule and its activation is often observed in cancers. 15 In mammals, AKT molecules were divided into three subtypes, namely, AKT1, AKT2, and AKT3. It has been reported that AKT1 regulates proliferation, growth, and hypertrophy, while AKT2 regulates glucose uptake, fatty acid transport, and glycogen synthase activity, 16 and AKT3, which is a serine/threonine-specific protein kinase at G2 phase, is required by cancer cells to preserve cell cycle and prevent cell death. 17 Compared with AKT1 and AKT3, AKT2 is most related to cancer cell survival, invasion, and metastasis. 18 It was revealed that AKT2 expression is downregulated by prohibitin-2 to promote prostate cancer cell migration 19 ; AKT2 plays an important role during breast cancer progression through regulating specific downstream proteins. 20 More importantly, studies reported that some miRNAs exerts their functions through the regulation of AKT2. 21 –23 However, the correlation and effects of miR-664b-5p and AKT2 on HCC development are less known.
The current research aimed to explore the effects and mechanism of miR-664b-5p and its target gene AKT2 in HepG2 and SUN-475 cells, hoping to gain a new understanding of molecular target and mechanisms of HCC treatment.
Materials and Methods
Tissue specimens and cell culture
Human HCC tissue specimens and adjacent normal tissues were obtained from 20 patients diagnosed with HCC between October 1, 2017, and July 4, 2019, at Shanxi Provincial People's Hospital. The study was approved by the Research Ethics Committee of the Shanxi Provincial People's Hospital and all patients signed the informed written consent before the study. All tumor tissues were histologically confirmed, and no patients had received radiotherapy or chemotherapy before the surgical resection. 24 The extracted tissue samples were pathologically confirmed according to the standard of the American Joint Committee on Cancer. 25 The extracted tissue samples were stored immediately at −80°C. Two liver cancer cell lines (HepG2 and SUN-475) and normal hepatocytes (THLE-2) were purchased from the American Tissue Culture Collection (ATCC, Manassas, VA). The cell lines were then cultured in Dulbecco's modified Eagle's medium (DMEM, 12100; Solarbio, Beijing, China) containing 10% fetal bovine serum (FBS, 11011-8611; Solarbio) with 5% CO2 in 95% humidity at 37°C.
Cell transfection
HepG2 and SUN-475 cells were cultured in 6-well plates (3 × 105 cells per well) for 24 h to reach cell confluence between 60% and 80%. For miRNA transfection, miR-664b-5p mimics (miR10022271-1-5, 5 nmol; RiboBio, China) and mimics control (MC, miR1N0000002-1-5; 5 nmol, RiboBio) were separately transfected into HepG2 and SUN-475 cells using Lipofectamine® 2000 transfection reagent (11668; Invitrogen) in 5% CO2 atmosphere at 37°C for 48 h. For AKT2 transfection, AKT2 cDNA was amplified by PCR and cloned into the pcDNA3.1 vector (V79020; Invitrogen) to form pcDNA-AKT2 overexpression plasmid. Then, the putative binding sites in the pcDNA-AKT2 were mutant to generate pcDNA-AKT2-mut plasmid. The empty plasmid served as a negative control (NC) of pcDNA-AKT2. According to the manufacturer's protocol, 3 mg plasmids were transfected into HepG2 and SUN-475 cells using Lipofectamine 2000 transfection reagent (11668; Invitrogen) in 5% CO2 atmosphere at 37°C for 48 h.
Targeted relationship prediction and dual-luciferase reporter assay
The potential relationship between miR-664b-5p and AKT2 was predicated by TargetScan. The fragment from AKT2 containing the predicted miR-664b-5p binding site was amplified by PCR and then cloned into a pmirGLO dual-luciferase miRNA target expression vector (E1330; Promega, Madison, WI) to generate AKT2 overexpression reporter vector. The putative binding site of miR-664b-5p in AKT2 was mutated by site-directed mutagenesis kit (Takara, Shiga, Japan), and the sequence of putative binding site was replaced and named as AKT2-mutated type (AKT2-mut). For luciferase reporter assay, AKT2 or AKT2-mut was, respectively, cotransfected with miR-664b-5p mimics or pmirGLO empty vector in HepG2 and SUN-475 cells using Lipofectamine 2000. The Dual-Luciferase® Reporter Assay System (E1910; Promega) was performed to determine luciferase activity. Firefly luciferase was normalized to Renilla luciferase.
Quantitative real-time polymerase chain reaction
Total RNA was isolated from the HepG2 and SUN-475 cell lines, and HCC's tissues and corresponding tissues, using the TRIzol reagent (15596018; Invitrogen) following the manufacturer's protocols. For miRNA analysis, reverse transcription reaction and quantitative real-time polymerase chain reaction (qRT-PCR) of miR-664b-5p were carried out by Hairpin-it TM miRNA and U6 snRNA Normalization RT-PCR Quantitation Kit (E22001-E22010; GenePharma, Shanghai, China). Parameters of qRT-PCR were set at 95°C for 3 min, 40 cycles at 95°C for 12 s, and at 62°C for 40 s in the 7500 Fast Real-Time PCR System (Applied Biosystems).
For analysis of mRNA expression, reverse transcription was performed using the PrimeScript™ RT reagent kit with gDNA Eraser (RR047A; Takara, Japan). qRT-PCR assays were carried out in 7500 Fast Real-Time PCR System (Applied Biosystems) using TB Green® Premix Ex Taq™ II (RR820Q; Takara) under the following conditions: predenaturation at 95°C for 30 min, followed by 40 cycles at 95°C for 5 s and at 60°C for 30 s, followed by melt curve conditions at 95°C for 5 s, at 60°C for 1 s, and annealing at 50°C for 30 s. MiR-664b-5p and AKT2 expression levels were, respectively, normalized to the levels of U6 snRNA (U6, E22001-E22010; GenePharma) and GAPDH by the 2−ΔΔCT method. 26 GAPDH was synthetized by Sangon Biotech. The sequences of primers used in the research are listed in Table 1.
Primer Sequences Used for Quantitative Real-Time Polymerase Chain Reaction
Cell viability assay
The cell viability was determined by CCK-8 assay (CA1210; Solarbio). The cells (2 × 103/well) were seeded into 96-well plates, which were cultured with CO2 in an incubator for 24, 48, and 72 h. Next, the CCK-8 solution (10 μL) was added into the cells and cultured for another 4 h. At last, the plates were put under a microplate reader (SpectraMax iD5; Molecular Devices), and the absorbance was determined at 450 nm.
Flow cytometry analysis
After precipitating and resuspending the cells, HepG2 and SUN-475 cells (0.5 × 104) were stained by the Annexin V-FITC Apoptosis Detection Kit (CA1020; Solarbio) in the dark. The apoptotic rates of HepG2 and SUN-475 cells were quantitated by flow cytometry in flow cytometer Accuri™ C6 (BD Biosciences). The data were analyzed by CellQuest software 3.3 (Becton-Dickinson).
Cell migration and invasion assay
The migration and invasion of HepG2 and SUN-475 cells were determined by transwell chambers (8 μm pores; Corning, Inc., Corning), with chambers precoated by Matrigel (BD Biosciences, San Jose, CA) used for migration assay and un-precoated chambers used for invasion assay. HepG2 and SUN-475 cells (1 × 105/well) were, respectively, cultivated into transwell chambers (Corning, Inc.) in the upper chambers containing 200 μL DMEM, while 500 μL DMEM containing 10% FBS was added into the lower wells as the chemoattractant. After 24 h, HepG2 and SUN-475 cells in the upper chamber were removed by a cotton swab, while the cells adhered to the lower surface were fixed by 4% precold methanol for 15 min and stained by 0.1% crystal violet solution (Sigma-Aldrich) for 20 min at room temperature. Cell number was counted under 10 random fields per filter by a 200 × microscope (Ts2r-FL; Nikon, Japan).
Western blot assay
Lysates of HepG2 and SUN-475 cells were quickly lysed for protein isolation and analysis. Briefly, for the detection of AKT2 expression, HepG2 and SUN-475 cells were washed twice by phosphate-buffered saline, and then, the total proteins were extracted using RIPA buffer (Thermo Fisher Scientific, Rockford, IL) with a complete protease inhibitor cocktail (Roche Diagnostics GmbH, Mannheim, Germany) and reacted on ice for 30 min. Subsequently, the supernatants were collected and the protein concentration of supernatants was determined by the BCA protein assay kit (Solarbio).
After centrifugation and quantification of proteins, the proteins (30 μg/lane) were separated on 6%–24% sodium dodecyl sulfate/polyacrylamide gel electrophoresis gels and transferred onto polyvinylidene difluoride membranes (Millipore Corp., Bedford, MA). After that, the membranes were then blocked by 5% (w/v) nonfat milk in tris-buffered saline containing 0.1% Tween-20 (w/v) buffer for1 h under room temperature before being exposed to specific primary antibodies (anti-AKT2 [mouse, 1:1000, ab175354; Abcam]; anti-GAPDH antibody [mouse, 1:500, ab8245; Abcam]) at 4°C overnight; GAPDH served as the internal control. After that, the membranes were incubated with the horseradish peroxidase-conjugated secondary antibody (goat anti-mouse, 1:2000, ab205719; Abcam) at room temperature for 2 h, and then washed three times for 10 min. Finally, a specific protein signal was shown by an ECL detection kit (ECL; Promega), and target bands were digitized by ImageJ software (ImageJ 1.8.0; National Institute of Health).
Statistical analysis
The data are shown as mean ± standard deviation. Paired t-test was used for paired sample comparison. Comparisons of multiple groups were performed by one-way analysis of variance. p-value of <0.05 was considered statistically significant. Statistical analysis in the study was processed by SPSS 17.0 software (SPSS, Inc., Chicago, IL).
Results
miR-664b-5p was downregulated in human HCC and hepatocellular cells
qRT-PCR was performed to determine the expression of miR-664b-5p in HCC tissues and hepatocellular cells. The results showed that miR-664b-5p was downregulated in HCC tissues compared with the adjacent normal tissues (Fig. 1A, p < 0.01), similarly, miR-664b-5p had a low expression in hepatocellular cells (HepG2 and SUN-475) compared with normal hepatocytes (Fig. 1B, p < 0.01).
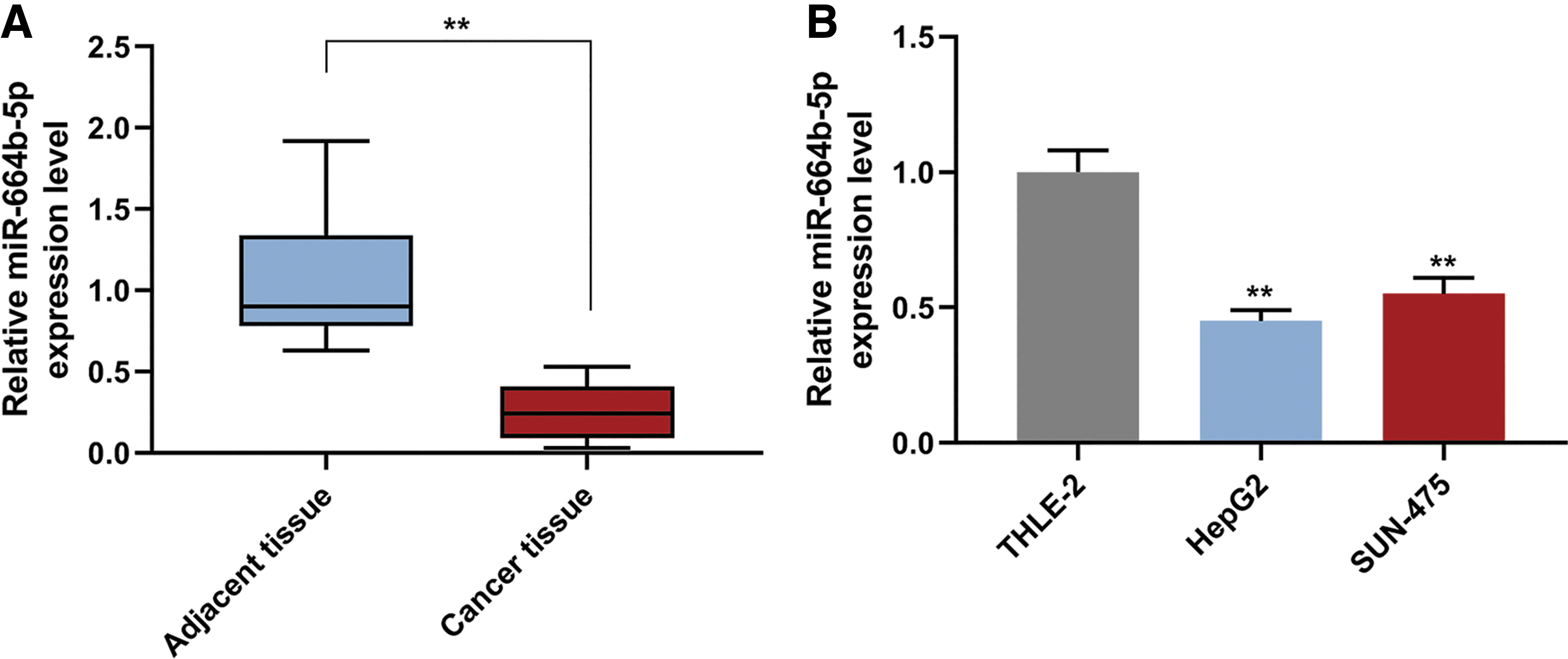
The expression of miR-664b-5p in liver cancer tissue and cells.
Overexpression of miR-664b-5p mitigated viability, promoted apoptosis, inhibited migration and invasion of HepG2 and SUN-475 cells
To investigate the role of miR-664b-5p on HCC progression, the authors successfully transfected the miR-664b-5p mimic into HepG2 and SUN-475 cells (Fig. 2A, p < 0.01), and the cell viabilities were measured at 24, 48, and 72 h. CCK-8 analysis showed that overexpression of miR-664b-5p significantly decreased the cell viability at 48 and 72 h (Fig. 2B). Flow cytometry found that the apoptotic rates of HepG2 and SUN-475 cells transfected with mimics were 15.48% and 22.12%, while the apoptotic rates in their controls were 6.27% and 6.6%, respectively. The data revealed that overexpression of miR-664b-5p greatly elevated apoptotic rates in HepG2 and SUN-475 cells compared with their Control groups (Fig. 2C, p < 0.01),. The migration and invasion assays were performed to determine the effect of miR-664b-5p on the metastatic potential of HCC. From the transwell experiment, the results revealed that overexpression of miR-664b-5p significantly decreased migration and invasion rates in HepG2 and SUN-475 cells (Fig. 2D and E, p < 0.01, scale bar = 50 μm). These data suggested that miR-664b-5p played an important part in regulating viability, apoptosis, and metastasis of HepG2 and SUN-475 cells.

The effect of miR-664b-5p on viability, apoptosis, migration and invasion of HepG2 and SUN-475 cells.
miR-664b-5p directly interacted with AKT2 in HepG2 and SUN-475 cells
Bioinformatic results found that AKT2 had binding sites in miR-664b-5p (Fig. 3A). Dual-luciferase reporter assay was performed to verify whether AKT2 directly targeted miR-664b-5p, and the authors observed that the luciferase activity of miR-196b-5p + AKT2 group was significant lower than control + AKT2 groups and miR-196b-5p + AKT2-mut groups in HepG2 and SUN-475 cells (Fig. 3B, p < 0.01). There were no statistically significant differences found in relative miR-664b-5p expression level between AKT2 and NC groups in HepG2 and SUN-475 cells. In comparison with the AKT2 group, the relative expression level of miR-664b-5p in the AKT2 + Mimic group greatly increased (Fig. 3C, D, p < 0.01). In comparison with the Control and NC groups, relative expression level of miR-664b-5p in the Mimic group increased noticeably in HepG2 and SUN-475 cells (Fig. 3C, D, p < 0.01). qRT-PCR and Western blot were performed to further investigate the interaction between miR-664-5p and AKT2, and the results showed that compared with Control and NC groups, relative levels of AKT2 protein and mRNA expression of the AKT2 group were significantly increased in HepG2 and SUN-475 cells (Fig. 3E–G, p < 0.01), but greatly decreased in Mimic groups (Fig. 3E–J, p < 0.01). Relative levels of AKT2 protein and mRNA expression of AKT2 + Mimic group were significantly decreased compared with AKT2 groups (Fig. 3E–J, p < 0.01), but greatly increased in the AKT2 + Mimic group compared with the Mimic group (Fig. 3E–J, p < 0.01). Meanwhile, AKT2 expression was noticeably higher in cancer tissues than that in adjacent tissues (Fig. 3K). Pearson correlation analysis showed that miR-664b-5p expression level was negatively correlated with AKT2 expression level (Fig. 3L).

AKT2 was a target of miR-664b-5p and the expression of AKT2 was partly reversed by miR-664b-5p mimic
Overexpression of AKT2 partly regulated the effect of miR-664b-5p mimic on cell viability and apoptosis in hepatocellular cells
CCK-8 assay demonstrated that viabilities of AKT2 group increased significantly at 48 and 72 h compared with that of Control group and NC group (Fig. 4A, p < 0.05 or p < 0.01), while viabilities of HeppG2 and SUN-475 cells in Mimic group decreased greatly at 48 and 72 h (Fig. 4A, p < 0.05 or p < 0.01). In addition, the viabilities of HepG2 and SUN-475 cells in AKE2 + miR-664b-5p Mimic group (AKT2 + Mimic) decreased significantly at 48 and 72 h compared with AKT2 group (Fig. 4A, p < 0.05 or p < 0.01), while the viabilities increased significantly compared with Mimic group (Fig. 4A, p < 0.05 or p < 0.01). These results demonstrated that miR-664b-5p mimic mitigated cell viability and promoted apoptosis of HepG2 and SUN-475 cells, and such effects can be partly reversed by AKT2 overexpression.
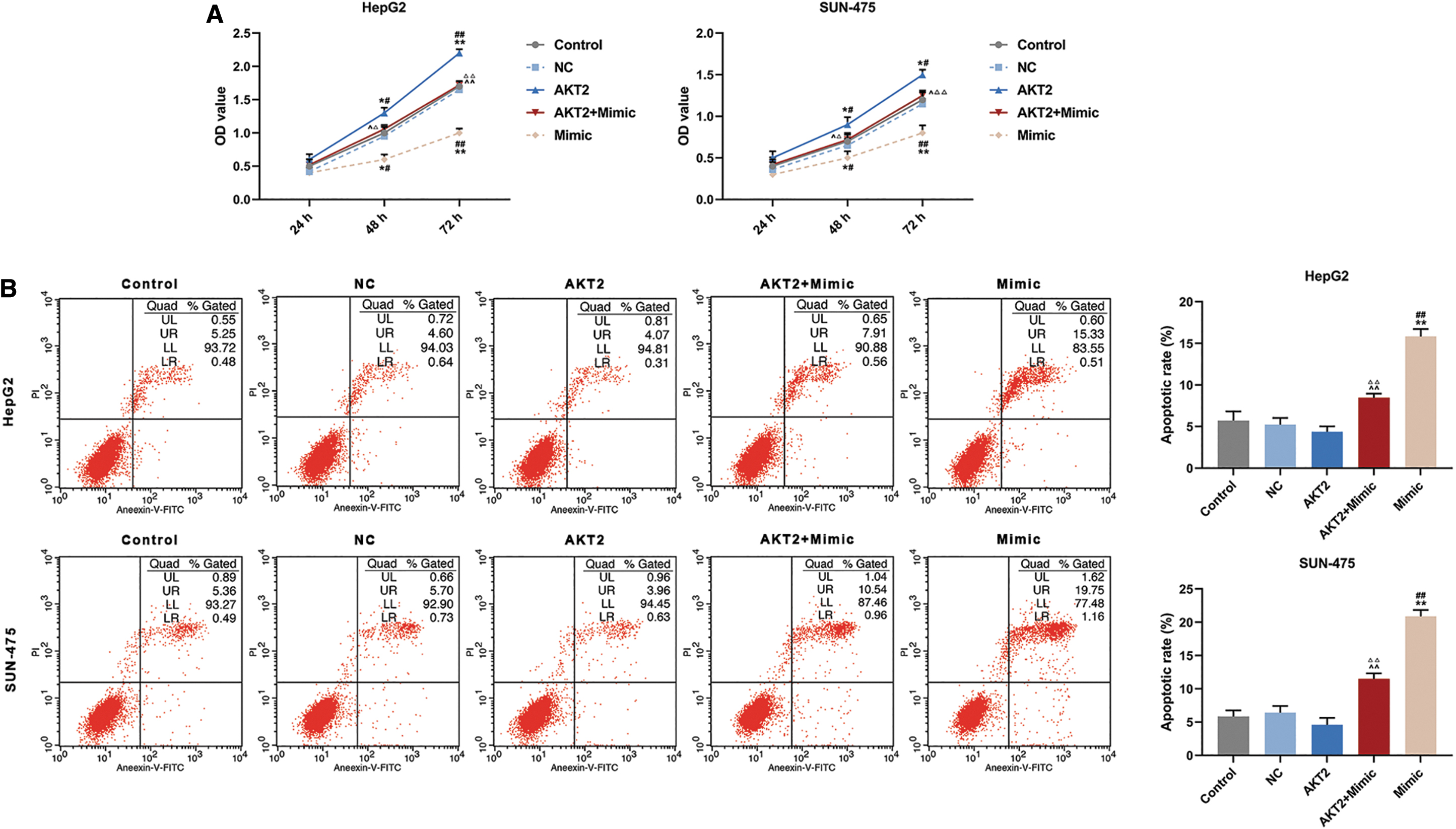
Overexpression of AKT2 partly regulated the effect of miR-664b-5p mimic on viability and apoptosis in HepG2 and SUN-475 cells.
Meanwhile, similar experimental results were found in flow cytometry analysis. The apoptotic rates of HepG2 and SUN-475 cells were determined in different groups (Fig. 4B). Compared with Control and NC groups, apoptotic rates of HepG2 and SUN-475 cells showed a decreased trend in AKT2 group. Apoptotic rates of HepG2 and SUN-475 cells in the AKT2 + Mimic group increased significantly compared with Control and NC groups (Fig. 4B, p < 0.01), but greatly decreased in AKT2 + Mimic group compared with Mimic group (Fig. 4B, p < 0.01).
Overexpression of AKT2 partly regulated the effect of miR-664b-5p mimic on migration and invasion in HepG2 and SUN-475 cells
The authors found that overexpression of AKT2 markedly increased migration and invasion in A549 and SPC-A1cells (Fig. 5A–F, p < 0.05 or p < 0.01), while overexpression of miR-664b-5p markedly decreased migration and invasion rates (Fig. 5A–F, p < 0.01). Migration and invasion rates of HepG2 and SUN-475 cells in AKT2 + Mimic group decreased greatly compared with AKT2 group (Fig. 5A–F, p < 0.01), but significantly increased in AKT2 + Mimic group compared with Mimic group (Fig. 5A–F, p < 0.05 or p < 0.01).

Overexpression of AKT2 partly regulated the effect of miR-664b-5p mimic on migration and invasion in HepG2 and SUN-475 cells.
Discussion
Vascular endothelial growth factor receptor, epidermal growth factor receptor, fibroblast growth factor receptor, and platelet-derived growth factor receptor have been identified as the targets in HCC treatment. 27 However, although the inhibition of these targets has been shown as effective, none of them demonstrated desired clinical results. 28 Therefore, it is necessary to further explore a new effective target and the effect mechanism of the target in the treatment of patients with HCC.
miRNA expression has regulatory potential, tissue, and disease specificities, and is therefore recognized as a potential target to help diagnose and predict different types and stages of cancer. 29 Studies demonstrated that miRNAs play a pivotal roles in the development of liver cancer. 30,31 Previous study supported that miRNA-519c-3p acts as a tumor oncogene, as it promotes tumor growth and metastasis of HCC. 32 miR-125b-5p inhibits cell proliferation, migration, and invasion of HCC cells. 33 Certain miRNAs have been shown to promote or inhibit HCC progression, and these results revealed that miR-664b-5p can act as a tumor suppressor in HCC progression. In this study, the authors observed that miR-664b-5p was downregulated in liver cancer tissue, suggesting that miR-664b-5p could be a potential therapeutic target or biomarker for HCC. To confirm whether miR-664b-5p was a novel effective target in HCC progression, the authors further explored its effect on cell viability, migration, invasion, and apoptosis of HepG2 and SUN-475 cells, and found that miR-664b-5p functioned as a potential tumor suppressor to inhibit the development of HCC.
Evidence indicated that AKT2 plays a pivotal in the development of cancer. 18 It has been revealed that AKT2 expression is downregulated to promote PCa cell migration. 19 AKT2 plays an important role during breast cancer progression via regulating specific downstream proteins, 20 and promotes colorectal cancer metastasis. 34 More importantly, increasingly researches reported the effect of AKT2 on HCC. 35,36 miRNAs exert their biological effects through modulating target genes. 37 Therefore, to explore the underlying effect mechanism of miR-664b-5p in HCC, the authors explored the relationship between miR-664b-5p and AKT2. miRNAs play pivotal roles in tumorigenesis and metastasis by negatively modulating gene expression through complementary binding to the 3′-untranslated region of target mRNAs. 38 In the present study, bioinformatic analysis predicted that miR-664b-5p can target the 3′-untranslated region of AKT2. Such a prediction was further verified by performing dual-luciferase reporter assay, qRT-PCR, and Western blot analysis, and the data also showed that AKT2 expression can be inhibited by the overexpression of miR-664b-5p in HepG2 and SUN-475 cells.
The present study is the first to report that miR-664b-5p suppressed the viability, migration, and invasion, but promoted apoptosis of HCC cells through negatively regulating AKT2. Accumulated researches showed that AKT2 was an miRNA target gene in HCC development. For example, Ma et al. showed that miR-296-5p downregulates AKT2 to inhibit HCC cell proliferation, migration, and invasion 22 ; research reported that miR-302b inhibits cell invasion and metastasis through directly targeting AKT2 in HCC cells 39 ; AKT2 was found to be a critical target of miR-137 in modulating cancer development and metastasis in HCC. 40
This research revealed the molecular mechanism through which miR-664b-5p suppressed the development of HepG2 and SUN-475 cells through inhibiting AKT2 expression. However, further studies in vivo are required to determine the effects of miR-664b-5p in animals with HCC and the clinical relevance of miR-664b-5p to HCC progression. Although the precise mechanisms are not fully clarified, this study suggests that miR-664b-5p may play a pivotal role in modulating the progression of HCC cells and miR-664b-5p could be explored as a potential therapeutic target for HCC.
Conclusions
In conclusion, miR-664b-5p acts as a cancer suppressor through negatively regulating AKT2 expression in HepG2 and SUN-475 cells, suggesting that miRNA-644b-5p could be used as a potential therapeutic strategy for HCC treatment.
Authors' Contributions
Substantial contributions to conception and design: H.L. Data acquisition, data analysis and interpretation: D.G. and Y.Z. Drafting the article or critically revising it for important intellectual content: S.Y. and R.Z. Final approval of the version to be published: All authors. Agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved: H.L. All coauthors have reviewed and approved of the article before submission.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.