
Abstract
Background:
Kinesin family member 22 (KIF22) is known as a regulator of cell mitosis and cellular vesicle transport. The alterations of KIF22 are associated with a series of tumors; however, its possible role in the progression of colon cancer is still unclear.
Materials and Methods:
This retrospective research collected 82 paired tissues with colon cancer. KIF22 protein and mRNA expression levels were detected by immunohistochemistry assays and Immunoblot assays, respectively. Short hairpin RNA (shRNA) plasmids were used to suppress the expression of KIF22 in HCT116 and HT29 cells, and the silencing efficiencies of shRNA plasmids targeted KIF22 were detected by quantitative PCR assays and immunoblot assays. In addition, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) assays and xenograft tumor growth assays were performed to observe cell proliferation in vitro and in vivo.
Results:
In human colon cancer tissues, the expression level of KIF22 was increased and correlated with clinical pathological features, including tumor stage and clinical stage (p = 0.034, and p = 0.015, respectively). Suppression of KIF22 inhibited cell proliferation and xenograft tumor growth.
Conclusion:
KIF22 might play an important role in the regulation of cell proliferation in colon cancer and might therefore serve as a promising therapeutic target.
Introduction
Colon cancer is a common malignant tumor in the digestive tract, which is the second most common cancer in women and the third in men. 1 In view of the lack of obvious early symptoms, early diagnosis of colon cancer is still difficult. 2,3 In addition, colon cancer has a high metastatic rate, and ∼10%–15% of patients with colon cancer has brain metastasis at the time of diagnosis. 4 In decades, colon cancer became the third leading cause of death in the world. 5 Despite advances in diagnosis and treatment, the main therapeutic targets could hardly meet the therapeutic expectations, and the 5-year survival rate for advanced colon cancer has not improved obviously over the years. 6 An increasing number of patients with colon cancer as well as the poor prognosis have been promoting the continuous improvement of early diagnosis and more findings of novel therapeutic targets.
Vale et al. first reported Kinesin in giant squid axons. 7 –12 Notably, kinesin family has been reportedly associated with transporting important molecules in cells. 11,12 For example, KIF26B was overexpressed in breast cancer and associated with poor prognosis, associating with larger tumor, lymph node metastasis. Moreover, the expression of KIF26B was positively correlated with ER. 13 Similarly, Kinesin family member 22 (KIF22), also named kinesin-like DNA binding protein (KID), could bind to both microtubules and chromosomes, transporting organelles, proteins, and mRNAs. Subsequently, previous studies indicated that KIF22 was widely expressed in different species and tissues, such as mammalian neuronal cells, sea urchin eggs, and Drosophila. 8 –10 KIF22 has been reported to be involved in the progression of several cancers. 14 In recent studies, the effects of KIF22 on transcription regulation were reported in some types of cancers. 15 –17 Yu observed that inhibition of KIF22 decreased cell proliferation in cancer by delaying mitotic exit. 15 They found that KIF22 had a high expression level in several cancers tissues and promoted cell proliferation by suppressing the expression of CDC25C to accelerate mitotic exit. 15 However, the expression levels and roles of KIF22 in colon cancer remain unclear.
In this study, we observed that the KIF22 protein was abnormally overexpressed in colon cancer tissues compared with the normal paracancer tissues, and the patients with high expression of KIF22 had higher tumor stage and clinical stage. Knockdown of KIF22 inhibited cell proliferation, xenograft tumor growth both in vitro and in vivo. Our findings showed that KIF22 might play an important role in colon cancer, and therefore KIF22 could be considered as a potential therapeutic target in colon cancer.
Materials and Methods
Tissues collection and immunohistochemical assays
Eighty-two tissues of colon cancer and the adjacent normal tissues were collected from patients who underwent surgery in both the Tangxian People's Hospital in Hebei province and Baoding second hospital. Tissues were formalin fixed and paraffin embedded, and then sections were made in 4 μm. Those sections were heated in the oven at 75°C for 30 min and deparaffinized in xylene, rehydrated in alcohol (100%, 100%, 95%, 85%, and 75%). Immunohistochemical (IHC) assays were introduced according to the specifications of the assays (Cell Signaling Technology, Danvers, MA). In brief, antigen was retrieved by citric acid buffer in a microwave for 15 min, and then cooled at room temperature. By using 3% H2O2 we blocked endogenous peroxidase for 5 min at room temperature and washed three times using phosphate-buffered saline (PBS) buffer. Then, sections were incubated with mouse anti-KIF22 antibody (Thermo Fisher Scientific, Waltham, MA; 1:200) at 4°C overnight. The second day, sections were washed three times in PBS and incubated with secondary antibody-horseradish peroxidase (Proteintech, Sanying, China; 1:1000) for 1 h at room temperature, washed, stained with diaminobenzidine (Cell Signaling Technology, Pudong, China) for 5 min. The images were captured by microscope (Nikon, Tokyo, Japan).
We scored for KIF22 nuclear expression based on staining area and staining intensity. We used semiquantitative results to judge the percentage of microscopically positive cells and staining intensity. The number of positive cells was <5% for 0, 5%–25% for 1, and 26%–50% for 2. In points, 51%–75% were 3 points, and 76%–100% were 4 points. Positive staining intensity is given as follows: 0 for no color, 1 for light yellow, 2 for brown, and 3 for brown. The two scores are multiplied by the positive rating: 0 is negative (−), 1–4 is weakly positive (+), 5–8 is positive (++), and 9–12 is strongly positive (+++). In this study, we divided the positive staining of KIF22 expression as low (− and +) and high (+ and ++) expression.
Cell culture
HCT116 (ATCC) and HT29 (ATCC) cells were maintained with high glucose DMEM (Gibco, Waltham, MA), including 10% fetal bovine serum (FBS; Gibco) or PRMI 1640 (Gibco), respectively, supplemented with 20% FBS. All cells were incubated in 5% CO2 at 37°C. The cell lines were purchased from the Beijing Xiehe cell resource center (Beijing, China).
Knockdown of KIF22 and formation of stable colony
The short hairpin RNA (shRNA) plasmids of KIF22 and negative control plasmids were provided by the hospital; shRNA sequence: 5′AAGCAAGATTGGAGCTACTCGTC3′. HCT116 and HT29 cells were transfected with shRNA and negative control plasmid using lipofectamine 3000 (Thermo, Waltham, MA). Then, after 72 h, shHCT116 and shHT29 cells were cultured in medium with Puromycin (Sigma, St. Louis, MO; 1.5 μg/mL) for 2 weeks to select stable colony.
Real-time quantitative polymerase chain reaction
Trizol (15596026; Invitrogen, CA) was used to extract total mRNA from human colon cancer cells. Subsequently the RNA was reverse transcribed by reverse transcriptase (M1701; Promega, WI). In addition, total mRNA was then reverse transcribed to produce cDNA by synthesis system. Quantitative PCR was performed using SYBR Ex Taq kit (638319; Takara, Japan), and the expression levels of KIF22 were normalized to the expression of GAPDH. KIF22 primer sequence: F-5′CGGCCTTTTACCAATGAGAGC3′, R-5′GACCAAGCAATTCTTTCTGAGACA3′. 18 GAPDH primer sequence: F-5′CGACCACTTTGTCAAGCTCA3′ and R-5′GGTTGAGCACAGGGTACTTTATT3′.
Immunoblot assays
Total protein was collected from cells using RIPA, PMSF (Solarbio, Beijing, China; 100:1), and 100 μL of it was added to six wells each, incubated for 30 min, centrifuged for 30 min at 4°C, and stored at −20°C until used. Thirty micrograms total protein was added to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and separation in 90 V. Then bands were removed to polyvinylidene fluoride membrane (Solarbio) in 250 mA for 130 min. The membrane was washed three times with TBST, and blocked in 10% nonfat milk, incubated with rabbit-anti-KIF22 antibody (Abcam, Cambridge, United Kingdom; 1:2000) overnight at 4°C. The second day, the membrane was washed with TBST and incubated with goat antirabbit antibody (Proteintech; 1:10,000) for 1 h at room temperature. Finally, protein bands were collected using ECL reagent kit (Solarbio) and collected images. β-Actin antibody (Cell Signaling Technology; 1:1000), Ki67 (Abcam; 1:2000), and proliferating cell nuclear antigen (PCNA; Cell Signaling Technology; 1:2000) protein obtained by western blot in a similar way.
Colony formation assays
Cells were suspended in the completed medium, and cell density was adjusted to 100 cells/mL with complete medium. Then, every well was added 2 mL complete medium with 200 cells in six-well plates, cultured at 37°C 5% CO2 for 2 weeks. Finally, cells were fixed with 4% paraformaldehyde (PFA) for 10 min and stained with 0.1% Crystal Violet. The number of colonies was counted by manual under the microscope.
CCK8 assay for cell proliferation
Cells were collected at log-growth phase and seeded into 96 wells, 3000 cells/well, with 100 μL PRMI 1640 containing 10% FBS, 1% penicillin-streptomycin solution in 5% CO2 at 37°C. After 48 h, 10 μL CCK8 (Sigma) was added to each well, and incubated for 4 h with cells in 5% CO2 at 37°C. Then, the result of optical density was detected by spectrophotometry at 570 nm. All experiments were repeated three times.
Xenograft tumor growth assays
Nude BalB/c mice (6–8 weeks, 18–22 g) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). shControl and shKIF22 HCT116 cells (5 × 106 cells/mice) were suspended and transplanted into subcutaneous armpit of nude mice. Tumor length and width were measured every 2 d, calculating tumor volume (V = 0.5 × L × W 2 , V: volume, L: length, W: width). 19 After 29 d, all mice were killed, and xenograft tumors were removed and photographed, fixed with 4% PFA. Protein expression level of KIF22 or Ki67 was detected by IHC staining in xenograft tumors.
Statistical analysis
Data were analyzed with SPSS 22.0 software (SPSS, Inc., IBM Corp., Armonk, NY). For the IHC experiments, associations between KIF22 expression and the clinicopathological features were evaluated using χ 2 tests. Data are shown as the mean ± standard deviation in vitro and in vivo experiments. Student's t-test was used for statistical comparisons. The statistics for the tumor growth curve have been calculated with two-way analysis of variance followed by post-test corrections for a single time point. A value of p < 0.05 was set to be statistically significant.
Result
KIF22 showed abnormal high expression in tumor tissues and associated with clinical features of colon cancer
To explore the potential link between KIF22 and colon cancer growth, 82 colon cancer tissues were collected from patients with surgical therapy. We further conducted IHC assays to detect KIF22 expression, and found that KIF22 was mainly located in the nucleus of colon cancer tissues (Fig. 1A). However, KIF22 expression was low in the normal paracancer tissues (Fig. 1B), compared with tumor tissues (Fig. 1A, B), suggesting a potential role of KIF22 in colon tumorigenesis.

KIF22 was overexpressed in colon cancer compared with the normal paracancer tissues.
We further divided these samples into two groups, including KIF22-low and high groups, according to the staining intensity of KIF22 (Fig. 1A). The clinicopathological characteristic difference between KIF22 high and low expression patients was analyzed. Data revealed that KIF22 expression in colon cancer was remarkably correlated with tumor grade and clinical stage; however, no significant link was identified between KIF22 expression and other clinical features (Table 1).
Relationships between KIF22 and Clinicopathological Characteristics in Eighty-Two Patients with Colon Cancer
KIF22, kinesin family member 22.
P < 0.05.
All data showed that KIF22 was highly expressed in human colon cancer tissues, and KIF22 was associated with the clinical characteristics of colon cancer.
Stable clone of suppression KIF22 in colon cancer cells by the transfection of shRNA plasmids
To further study the role of KIF22 in colon cancer, stable cell clone of suppression KIF22 in the cancer cells was established by shRNA. As shown in Figure 2A, according to the results of quantitative PCR assays, KIF22 mRNA levels in the cells transfected with its shRNA were lower than those in control cells (p < 0.05, respectively). Furthermore, the protein levels of KIF22 were confirmed using immunoblot in vitro. As shown in Figure 2B, KIF22 expression was decreased in protein levels when transfected with shRNA in vitro (p < 0.05, respectively).
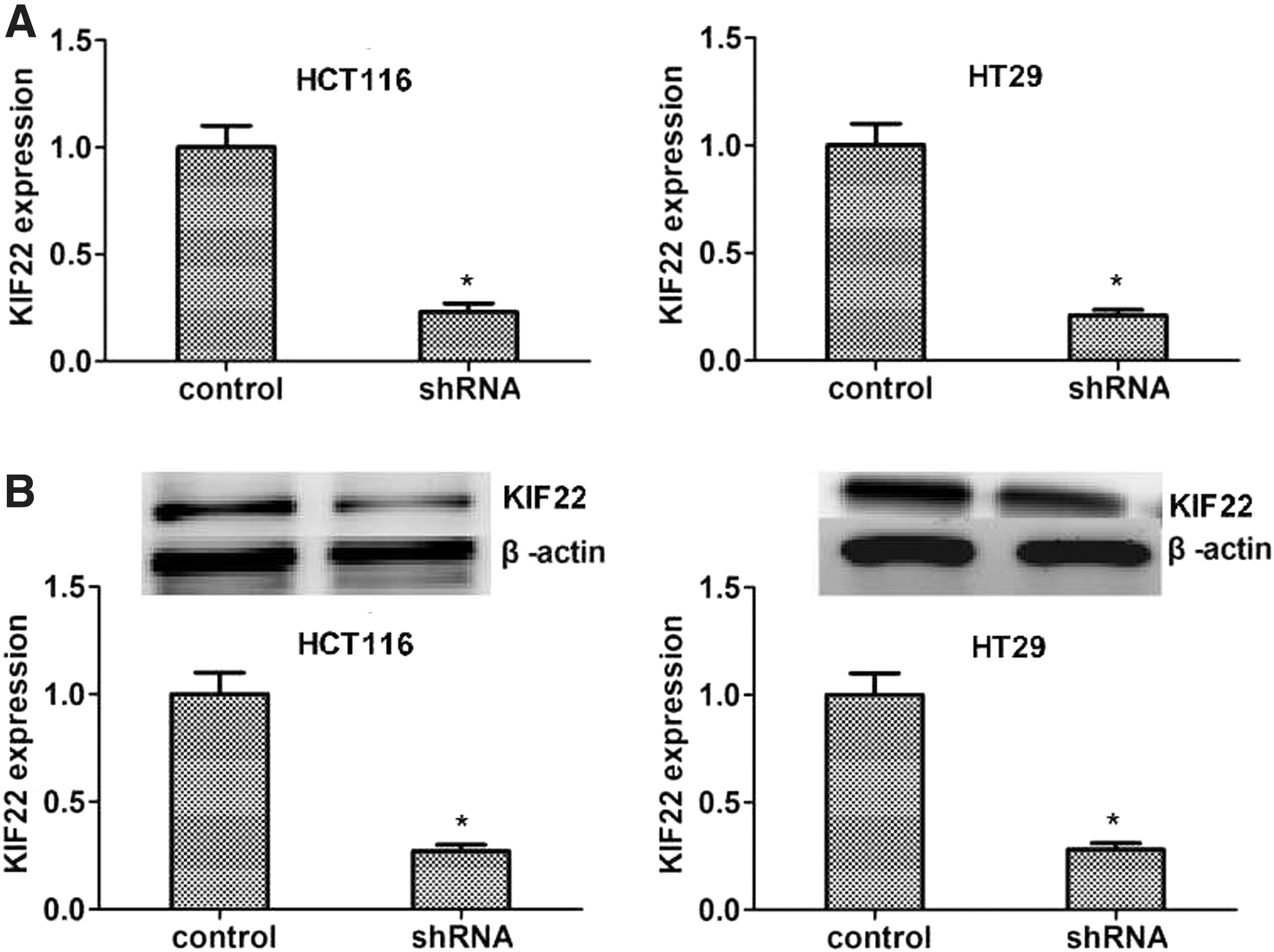
Stable clone of suppression KIF22 with shRNA in vitro.
Suppression of KIF22 inhibited proliferation of colon cancer cells in vitro
In previous reports, suppression of KIF22 inhibited cell proliferation in cancer cell. 15 However, there was no report of KIF22 in the progression of colon cancer. To explore the role of KIF22 in this cancer, colony formation assays were performed in vitro and cells were transfected with shRNA, showing that knockdown of KIF22 decreased colony formation capacity (Fig. 3A). Incubating for 2 weeks, compared with negative control cells, KIF22-depleted cells displayed fewer colonies (p < 0.05, respectively). To further assess cell proliferation triggered by KIF22, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) assays were introduced in above cells. As shown in Figure 3B, the result was similar to colony formation assays, KIF22-depleted cells had a low cell proliferation compared with negative control cells (p < 0.05, respectively). In previous studies, Ki67 18,19 and PCNA 20,21 were accepted protein markers associated with cell proliferation. Protein expression levels of Ki67 (Fig. 3C) and PCNA (Fig. 3D) were detected by immunoblot in cells transfected with shRNA and negative control cells, showing that suppression of KIF22 led to a low expression of Ki67 and PCNA (p < 0.05, respectively). These data indicated the involvement of KIF22 in cell proliferation of colon cancer in vitro.
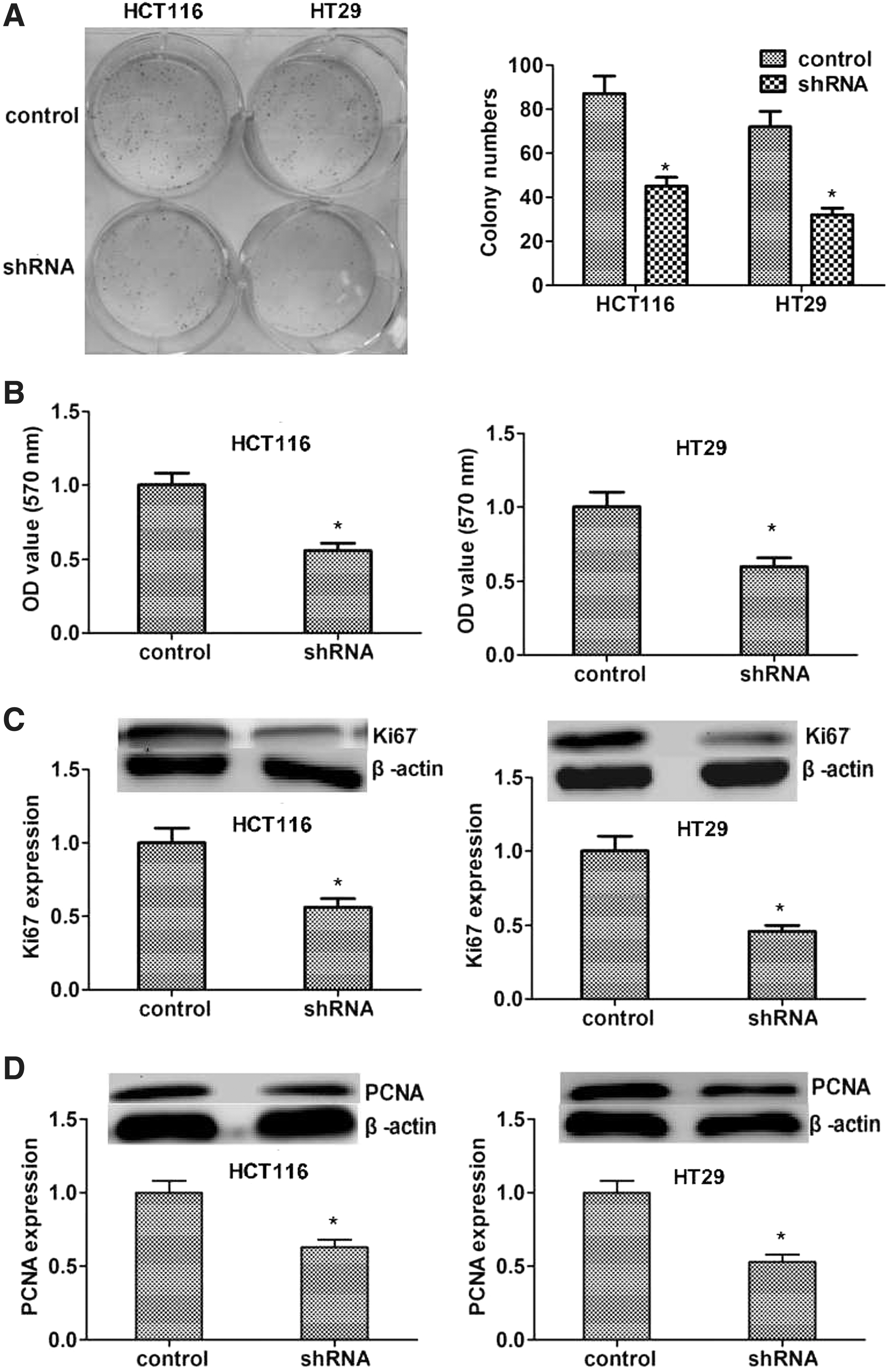
The depletion of KIF22 inhibitor proliferation in vitro.
Knockdown of KIF22 inhibited xenograft tumor growth
Above data showed that KIF22 effected cell proliferation in vitro. To further study the role of KIF22 in colon cancer, xenograft tumor growth assays were performed to observe tumor growth in mice. shControl cells and shKIF22 HCT116 (5 × 106 cells) were injected subcutaneously into armpit of mice, and tumor size was measured and calculated every 3 d. As shown in Figure 4A, xenograft tumor volume from shHCT116 cells was smaller than that from HCT116 cells at every checkpoint. After 30 d, all tumors were removed from mice, and expression level of KIF22 protein was observed by IHC in tissues of xenograft tumors, showing that tumors from control cells had higher expression than those from shKIF22 HCT116 cells (Fig. 4B, C).
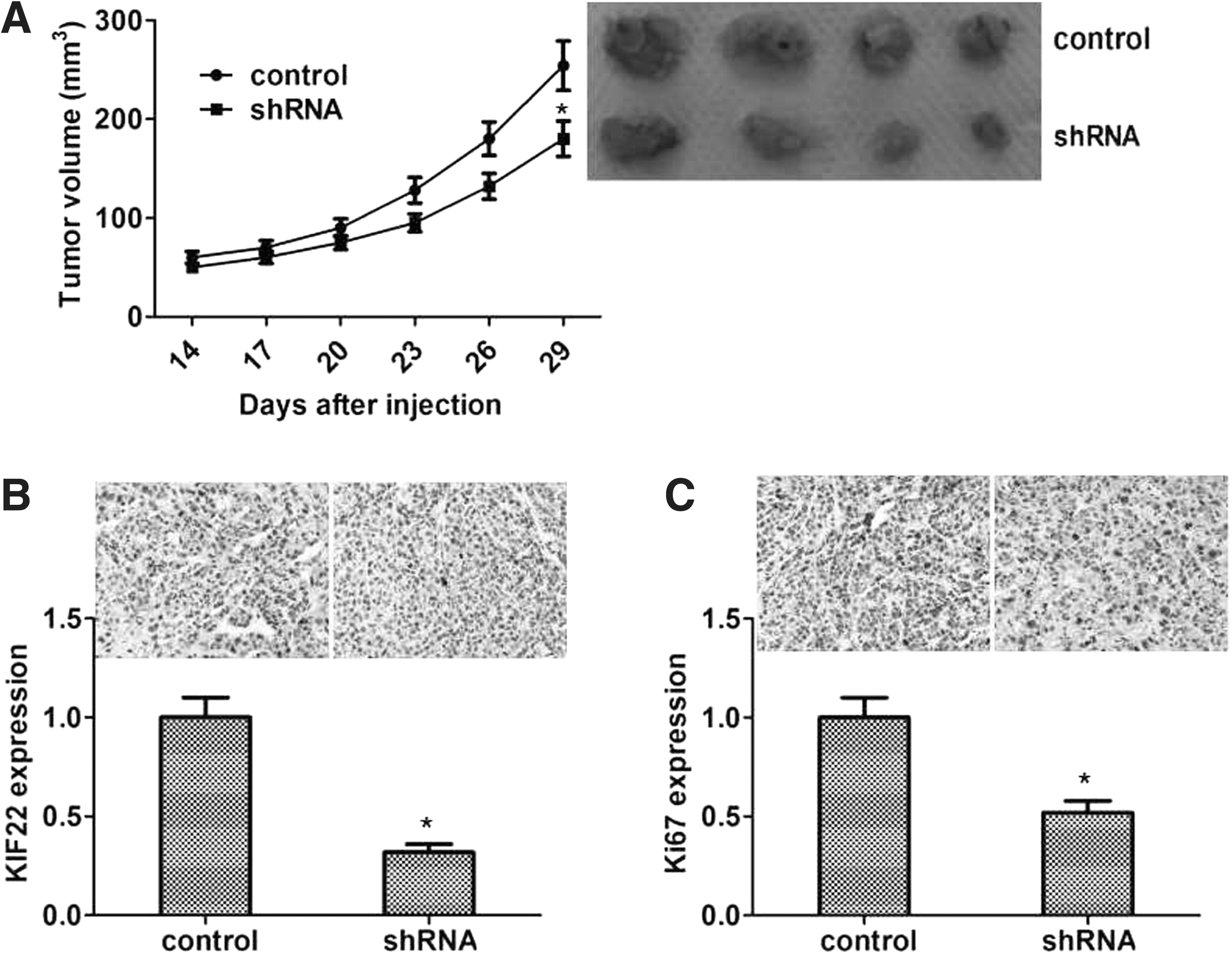
Knockdown of KIF22 inhibitor xenograft tumor growth.
Discussion
Colon cancer is a common malignancy of the digestive tract. Notably, the morbidity and mortality among patients with colon cancer have been dramatically increasing over decades. 22 Early diagnosis of colon cancer without any symptoms also emphasized the importance of early diagnosis of colon cancer. 4 Advanced colon cancer is highly metastatic, resulting in increasing mortality rates among those patients. 23 Therefore, it is urgent to find more effective biomarkers and therapeutic targets. In this study, we found that KIF22 was associated with the prognosis of patients with colon cancer, and confirmed that KIF22 can promote the development and progression of colon cancer through in vitro and in vivo experiments, suggesting that KIF22 could serve as a novel therapeutic target for colon cancer.
In previous studies, KIF22 was reported to show high expression in several cancer tissues and associated with proliferation. Yu detected mRNA and protein expression levels of KIF22 in 62 primary breast cancer tissues and paired adjacent normal tissues, showing that KIF22 was highly expressed in primary breast cancer compared with adjacent tissues. 15 Notably, to further explore the relationship between KIF22 and cancer progression, they analyzed KIF22 in several types of cancers in oncogene database, confirming that KIF22 was highly expressed in ovarian cancer, lung cancer, and cervical cancer. 15 In our study, 82 tissues of colon cancer and adjacent normal tissues were collected to detect the expression levels of KIF22. By analyzing KIF22 expression levels in cancer tissues with patients' clinical information, we noticed that expression levels of KIF22 were associated with clinical stage. Other studies also reported that kinesin family played an essential role in mitosis 14,24 –26 and the abnormal expression in cancers to contributing to proliferation, such as KIF18A, 27 KIF26B, 15 KIF14, 28 –30 and EG5. 31 –33 Specific inhibitors of EG5 are in Phase I or Phase II clinical trials. 34,35 Yu reported that KIF22-depleted cells displayed a fast S to G2/M phase but suppressed mitotic exit. 15 Similarly, our study showed that suppression of KIF22 obviously inhibited proliferation and xenograft tumors. KIF22 was known as a novel transcriptionally targeting CDC25C, and negatively regulated CDC25C in mRNA and protein level. 15 There exist several phosphorylated sites, including Ser427 and Thr463, in KIF22, 36 and Thr463 is conserved in many species. 37,38 Ohsugi et al. reported that the phosphorylation of KIF22 at Thr463 inhibited KIF22 connected to microtubules but increased the affinity to chromosomes. 36 Overexpressed KIF22 in cancers promoted cell proliferation by inhibiting CDC25C and increasing mitotic exit. 15 To further investigate the role of KIF22 in colon cancer, we established stable KIF22 suppression cells in vitro, and the cell proliferation assays showed that knockdown of KIF22 inhibited colony formation and cell growth. In previous reports, Ki67 and PCNA were protein markers to detect cell proliferation in cells, so we further evaluated the expression levels of Ki67 and PCNA in vitro, and found that Ki67 and PCNA had low expression in cells transfected with shRNA. Those data indicated that suppression of KIF22 inhibited cell proliferation by affecting the related proteins such as Ki67 and PCNA in vitro.
As was reported, human breast cancer cell line, MDA-MB-435, both control and KIF22 depletion cells were inoculated into the mammary fat pads of nude mice. Interestingly, after 7 weeks, the results showed that xenograft tumors from KIF22-depleted cells had small tumor volume. 15 Similarly, in our study, the animal experiments were designed to verify role in vivo. As shown in Figure 4A and B, suppression of KIF22 in cells blocked xenograft tumor growth in nude mice. This result was similar to cellular level. Miki reported that KIF22 was regulated by SIAH-1. 12 Pike reported a novel pathway between KIF22 and EGFR, CAR. KIF22 bound to microtubules, which were important in receptor traffic, including EGFR. 39,40 KIF22 could regulate microtubule stability and upregulated EGFR signaling, which promoted CAR phosphorylation and relocalization at plasma membrane to promote cell division. 41 In our study, suppression effects of KIF22 were observed, and then downstream pathways should be further detected. The limitation of this study is that this is a retrospective research with a small number of cases in single center. In addition, the overexpression of KIF22 should be conducted to further observe the role in vitro and in vivo, and the precise mechanism of KIF22 in the progression of colon cancer needs further study.
In summary, this is the first study of KIF22 in the progression of colon cancer, and we observed that KIF22 showed high expression of KIF22 in tissues of colon cancer and was associated with high clinical stage. Inhibition of KIF22 suppressed cell proliferation and xenograft tumors. Therefore, KIF22 might serve as a novel and promising molecular target for colon cancer therapy.
Ethics Approval and Consent to Participate
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. The animal study was carried out in accordance with the guidelines approved by the Animal Experimentation Ethics Committee of the secondary hospital of Tianjin Medical University. The protocol was approved by the Committee; all surgery was performed under sodium pentobarbital anesthesia, and all efforts were made to minimize suffering.
Consent for Publication
All of the authors have agreed to publish this article in our journal if it is accepted.
Availability of Data and Material
The dataset supporting the conclusions of this article is included within the article.
Footnotes
Author Contributions
B.L. and F.-C.Z. carried out the experiment of molecular biology and the animal experiments. S.-X.Y., S.-J.L., and B.-Y.L. participated in the design of the study and performed the statistical analysis. All of the authors conceived of the study, participated in its design and coordination, and helped to draft the article.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.