
Abstract
Background:
The effects of miR-524-5p on breast cancer (BC) have not been investigated, though studies show that miR-524-5p has an anticancer function. Thus, this study investigated the effects of miR-524-5p on BC cells and its potential molecular mechanism.
Materials and Methods:
The expression of miR-524-5p from the collected BC samples was determined. Cell counting kit-8 (CCK-8) assay was performed to examine the effect of miR-524-5p on BC cells viability. The target for miR-524-5p was predicted by bioinformatics and further verified by luciferase assay. Wound healing assay and transwell assay were performed to determine cell migration and invasion of BC cells. The expressions of Follistatin-like 1 (FSTL1) and related proteins in epithelial–mesenchymal transition (EMT) were detected by Western blotting and quantitative real-time polymerase chain reaction.
Results:
MiR-524-5p was low-expressed in BC samples, and upregulation of miR-524-5p suppressed BC cell viability, migration, and invasion. FSTL1 was predicted as a target for miR-524-5p. In addition, overexpressed FSTL1 effectively abolished the effect of miR-524-5p on inhibiting FSTL1 expression, and reversed the inhibitory effects of miR-524-5p on the migration, invasion of BC cells as well as the effect of miR-524-5p on regulating the expressions of matrix metalloproteinase 2 (MMP2), matrix metalloproteinase 9 (MMP9), E-cadherin, and N-cadherin.
Conclusions:
Our findings suggest that miR-524-5p targeting FSTL1 adversely affects the progression of migration, invasion, and EMT of BC cells, thus, miR-524-5p is possibly a target for BC treatment.
Introduction
Breast cancer (BC) is one of the major causes of cancer-related death among women worldwide. 1 Surgery and chemotherapy are therapeutic strategies, which are widely used for treating BC, and are frequently associated with adverse side-effects. 2,3 However, for patients with advanced BC, the efficacies of surgery and chemotherapy are low due to distant metastasis of cancer. 4 Distant metastasis is an important factor contributing to BC development, 5 –7 and many BC patients have metastatic BC at the time of diagnosis, and their 5-year survival rates are only ∼27%. 8 Similar to other cancers, the occurrence, development, and metastasis of BC are closely related to the abnormal expressions of multiple genes such as oncogenes, tumor-suppressor and metastasis-related genes. 9 Although early detection and treatment strategies of BC have been greatly improved, BC cells have strong invasion and metastasis abilities; as a result, the morbidity, mortality, and survival rates of BC patients are still not satisfactory. 10 –12 Therefore, a novel approach to the prevention and treatment of BC is urgently needed.
MicroRNAs (miRNAs) are noncoding RNAs with vital biological function in cells. 10 Evidence showed that abnormal expressions of miRNAs are highly related to the occurrence and development of various cancer types, and that miRNAs could function as typical oncogenes and tumor-suppressor genes. 13 In addition, recent studies have proved that miRNAs can be used as biomarkers for early screening, diagnosis, and treatment of various cancers. 14 –16 MiR-524-5p was reported to suppress melanoma cell growth through targeting BRAF and ERK2 17 and inhibiting glioma progression by directly regulating Hes1 and Jagged1. 18 Studies also showed that miR-524-5p has the ability to regulate cancer cell proliferation and epithelial–mesenchymal transition (EMT) progression. 19 However, there is a lack of study on the functions of miR-524-5p in BC.
In this study, the expression of miR-524-5p in BC tissue samples was determined, and the function of miR-524-5p in growth, migration, invasion, and EMT of BC cells was explored. Furthermore, we determined the possible target for miR-524-5p. The purpose of the research was to determine whether miR-524-5p is a target for BC treatment.
Materials and Methods
Cell culture
Human BC SK-BR-3 and MDA-MB-453 cell lines (American Type Culture Collection, Rockville, MD) 10,20,21 were cultured in Dulbecco's modified Eagle's medium (DMEM; Gbico, MA) containing 10% fetal bovine serum (FBS; Gbico) at 37°C with 5% CO2 in a humid environment.
Tissue samples
BC lesions and corresponding noncancerous samples (Table 1) were collected from 20 BC patients who underwent surgical excision at Zhuji People's Hospital between January 2017 and January 2019. BC tissues were stored in a refrigerator at −80°C for later use. All patients did not receive chemotherapy or radiotherapy before the surgery. The current research was approved by the Ethics Committee of Zhuji People's Hospital. Informed consent was signed by all participants.
Clinicopathological Variables and Expression of miR-524-5p
The median was the cutting point, higher than the median was the high expression, lower than the median was the low expression.
Transfection
MiR-524-5p mimics (mimic), Follistatin-like 1 (FSTL1) overexpression vector (FSTL1), mimic control and negative control (NC) were obtained from GenePharma (Shanghai, China). The mimic and FSTL1 were diluted to 20 μm by RNase-free H2O (Beyotime, Shanghai, China) and stored at −20°C in a refrigerator for later use. 1.0 × 106 BC cells were seeded into each well of six-well plates in 2 mL complete medium and grown until they were 60%–70% confluent. Two micrometers mimic and FSTL1, which were diluted by 100 μL DMEM, were added to 3 μL Lipofectamine 2000 (Invitrogen, MA). The above two mixtures were mixed and incubated for 15 min at room temperature. Finally, the mixed liquid was added to the cells of each well, and 1.8 mL DMEM was then added to the cells and held for an additional 48 h. Mimic control and NC were conducted in parallel.
Quantitative real-time polymerase chain reaction
miRNAs were extracted from clinical BC samples and BC cells (SK-BR-3 and MDA-MB-453 cells) using a miRcute miRNA Isolation Kit (TianGEN, Beijing, China). In brief, for BC samples, a grinding rod was used to grind the samples in liquid nitrogen in a 1.5 mL centrifugal tube containing lysis buffer, while for BC cells the cells were collected into 1.5 mL centrifugal tube and added to lysis buffer. Two hundred microliters chloroform was then added to the cells and shaken for 1 min. After resting the cells for 5 min at room temperature, the cells were centrifuged for 20 min (13,400 g), the miRNA solution was transferred into a new 1.5 mL tube, and added to ethanol and centrifuged for another 15 min (13,400 g). The sediments were miRNAs, and RNase-free H2O was used to dilute the miRNA sediments. Total RNAs were extracted from BC cells using TRIzol reagent (Invitrogen). In brief, the cells were lysed by TRIzol and collected into a new 1.5 mL centrifugal tube; chloroform was added to the tube and centrifuged for 20 min (1400 g). The supernatant was collected, and added to an equal volume of isopropanol and centrifuged for 5 min (1400 g), and the miRNA sediments were diluted using RNase-free H2O. Then, PrimeScript RT kit (Takara, Dalian, China) was used to reverse transcribe miRNAs and mRNAs into cDNAs according to the instructions. Finally, gene expression was determined by quantitative real-time polymerase chain reaction (q-PCR) assays using Verso 1-step RT-qPCR Kit (Thermo Scientific, MA) in ABI 7500 Fast Real-Time PCR System (Applied Biosystems, CA), and the condition of q-PCR was set at 95°C for 30 s, at 60°C for 30 s, 45 cycles at 60°C for 30 s. 17 U6 and β-actin served as internal references. All primer sequences are shown in Table 2. mRNA and miRNA were quantified by 2−▵▵CT method. 18
Quantitative Real-Time Polymerase Chain Reaction Primers
Cell counting kit-8 assays
Cell counting kit-8 (CCK-8; Bioengineering Institute, Nanjing, China) was used to determine cell viability. CCK-8 contains WST-8 [2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4disulfophenyl)-2H-tetrazolium, monosodium salt]. In CCK-8 measurement, the dye of WST-8 was reduced by dehydrogenase in cells to form a water-soluble orange-colored product (formazan). The amount of the produced formazan dye by cellular dehydrogenases is correlated with the number of living cells. Therefore, the cell viability can be simply estimated by recording the optical density of formazan at 450 nm using a microplate reader. 22 In brief, after the transfection, the cells were seeded into 96-well plates at 1.0 × 104 cells in 100 μL complete medium. After growing for 24, 48, and 72 h, the cells were incubated by CCK-8 reagent (0.5 mg/mL) for 15 min. Finally, the absorbance of each well was detected at 450 nm using a microplate reader (Infinite M200 PRO; Tecan Austria GmbH, Austria).
Wound healing assay
After the transfection was completed, the cells were placed into six-well plates at 3.5 × 105 cells in complete medium and continued to culture until the cell confluence reached 95%. Then, a vertical wound in each well was created by a 20 μL pipette tip, and DMEM without FBS was added into each well. Images ( × 100) in each well were collected at 0 and 24 h under a phase-contrast microscope (Axio Lab.A1 pol; Leica, Solms, Germany). ImageJ software (Version 1.8.0) was used to analyze the images in this assay.
Transwell assays
Transwell cell culture chambers were precoated by Matrigel (Corning Life Sciences, NY), and the transwell inserts were placed into a 24-well plate. The transfected cells were diluted into 2 × 105 and pipetted into the upper chambers containing suspension solution with 0.2 mL EMEM without FBS, and the corresponding complete medium was added into the lower chamber. The cells were incubated for 24 h, the upper-side of the polycarbonate membrane was wiped, leaving the underside of the membrane containing invaded cells. Finally, the cells were stained by crystal violet for 15 min at room temperature. The cell numbers from three random areas on each membrane were counted under a phase-contrast microscope ( × 200) (Axio Lab.A1 pol; Leica). ImageJ software (Version 1.8.0) was used to analyze the images.
Luciferase reporter assays
The fragments of FSTL1-3′-UTR containing wild-type and mutant binding sites for miR-524-5p were inserted into pmirGLO luciferase vectors (GenePharama). After the FSTL1-3′-UTR WT (or MUT) were cotransfected with mimic by Lipofectamine 2000 for 48 h, the cells were gathered to perform Dual-luciferase Reporter Assay (Promega, CA). The luciferase activity of the cells was determined by GloMax fluorescence reader (Promega). PsiCHECK-2 vector transfected into cells served as internal control.
Western blot assays
Total proteins from the cells were isolated by RIPA lysis buffer (Beyotime), and the total protein concentrations were determined by a BCA assay kit (Pierce, MA). Finally, total protein (30 μg) was separated on 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels, electroblotted and transferred to NC membranes (Millipore, MA). Then, the membranes were blocked by 5% skimmed milk for 1 h at room temperature and incubated with antibodies [FSTL1 (1:1000, ab11805, 50 kD; Abcam, CA), matrix metalloproteinase 9 (MMP9) (1:1000, ab73734, 78 kD; Abcam), matrix metalloproteinase 2 (MMP2) (1:1000, ab37150, 72 kD; Abcam), E-cadherin (1:1000, ab40772, 97 kD; Abcam), N-cadherin (1:1000, ab18203, 130 kD; Abcam), and β-actin (1:1000, 42 kD, ab8226; Abcam)] at 4°C overnight. Next day, HRP-conjugated secondary antibodies [goat antirabbit IgG secondary antibody (1:5000, L3012, Signalway Antibody, PA), goat antimouse IgG secondary antibody (1:5000, ab205719; Abcam), and donkey antigoat IgG secondary antibody (1:5000, ab6885; Abcam)] were incubated with the membranes for 1 h at room temperature. Finally, SuperSignal West Pico Chemiluminescent Substrate (Thermo Scientific) was used to incubate the membranes for signal detection. Image Lab™ Software (version 3.0) was used for densitometric analysis and quantification of the Western blotting data (Bio-Rad Laboratories, Inc., Hercules, CA).
Statistical analysis
Student's t test and one-way analysis of variance (ANOVA) were used to analyze the data by SPSS software (version 18.0). Post hoc tests were performed by LSD and Dunnet's. The data were shown as means ± standard deviation. All experiments were conducted in triplicate. Statistically significant result was shown as p < 0.05.
Results
MiR-524-5p was low-expressed in BC tissues
A comparative analysis of miR-524-5p expression level in clinic primary BC and adjacent samples was conducted, and the result revealed that miR-524-5p expression was obviously lower in BC tissue compared with that in adjacent tissues (p < 0.01) (Fig. 1A). Moreover, for clinical samples, we took the median as the cutoff point, higher than the median was regarded as high expression and lower than the median was regarded as low expression. As shown in Table 1, there was a significant negative correlation between TNM stage and the miR-524-5p expression in these patients.
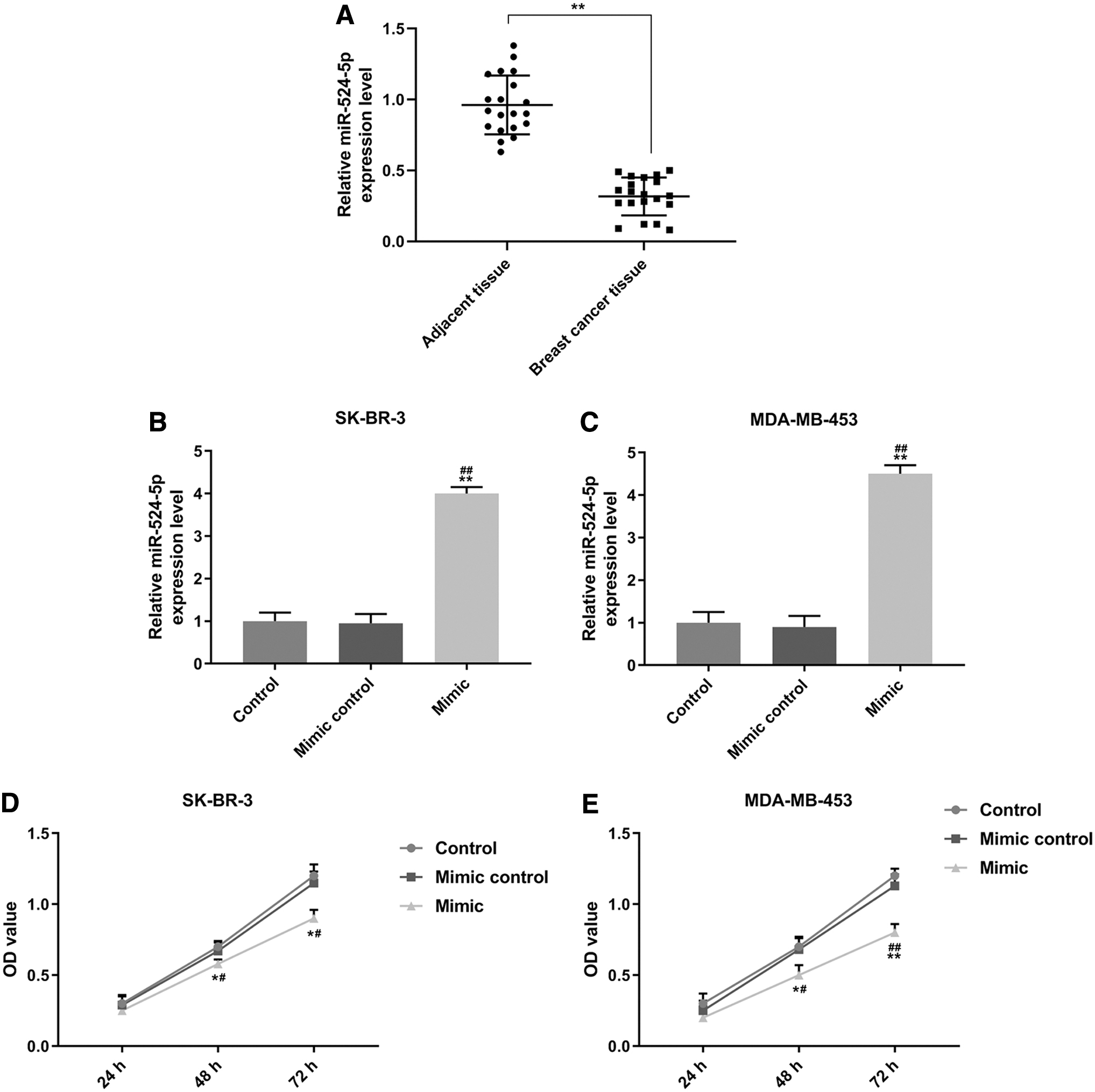
MiR-524-5p was low-expressed in BC tissues and inhibited BC cell viabilities.
Overexpressed miR-524-5p inhibited viabilities of SK-BR-3 and MDA-MB-453 cells
To determine the functions of miR-524-5p in BC cells, miR-524-5p mimic was transfected into BC cells, and subsequently, the changes in viabilities of SK-BR-3 and MDA-MB-453 cells were detected after 48 and 72 h. As shown in Figure 1B and C, the expression of miR-524-5p had no difference in control and mimic control groups, while it was upregulated in the two cells in mimic group (p < 0.01), indicating that miR-524-5p mimic was successfully transfected into the two cells. We also found that cell viability in mimic control and control groups had no statistical difference, while in mimic group the viability of the two cells was significantly reduced, compared with control and mimic control groups (48 h: p < 0.05; 72 h: SK-BR-3, p < 0.05; MDA-MB-453, p < 0.01, respectively) (Fig. 1D, E).
Overexpressed miR-524-5p inhibited migration and invasion of SK-BR-3 and MDA-MB-453 cells
Whether miR-524-5p regulated SK-BR-3 and MDA-MB-453 cell migration and invasion was determined, as shown in Figure 2A and B, 24 h after scratches, no significant difference was observed in the cell migration into the wounded (clear) area of the cell monolayers in control and mimic control groups. However, in the mimic group, relative migration rate was significantly reduced compared with control and mimic control groups (SK-BR-3, p < 0.05; MDA-MB-453, p < 0.01, respectively). As for invasion ability, as shown in Figure 2C and D, similar to wound healing assays, in the mimic group, the relative invasion rate was significantly reduced compared with control and mimic control groups (SK-BR-3, p < 0.01; MDA-MB-453, p < 0.01, respectively). Thus, the results showed that miR-524-5p could inhibit the migration and invasion of SK-BR-3 and MDA-MB-453 cells.
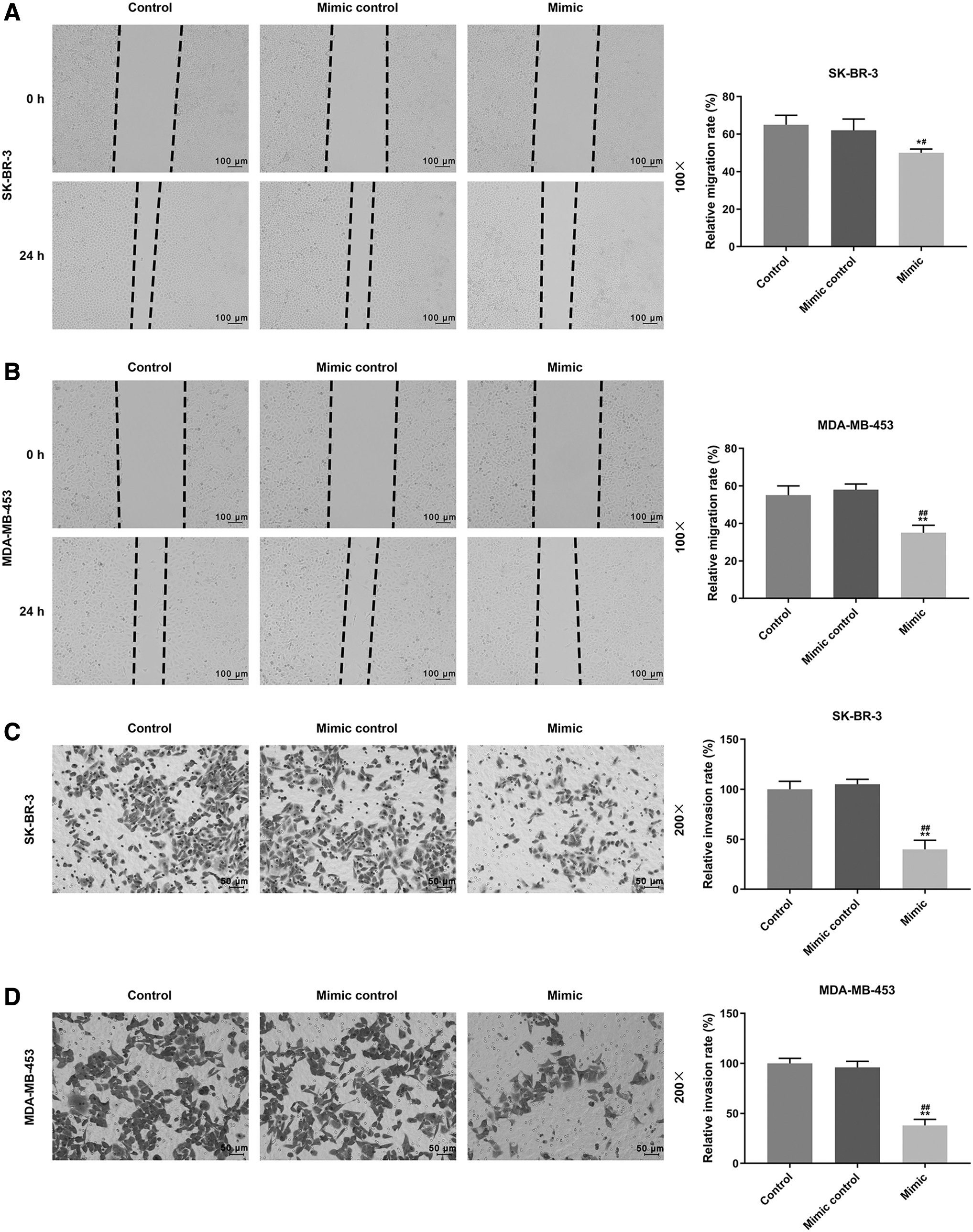
Overexpressed miR-524-5p inhibited migration and invasion of SK-BR-3 and MDA-MB-453 cells.
MiR-524-5p specifically targets FSTL1
TargetsScan predicted that the possible target for miR-524-5p was FSTL1, as the target sequence at 1945–1951 base was in pairs with FSTL1-3′-UTR in the study (Fig. 3A). To further confirm the prediction, luciferase reporter assays were conducted. As shown in Figure 3B and C, luciferase activities decreased in the two cells cotransfected with wild-type FSTL1 and miR-524-5p mimic compared with NC group (SK-BR-3, p < 0.01; MDA-MB-453, p < 0.01, respectively). However, after cotransfection with FSTL1 mutation (mut) and miR-524-5p mimic, no differences in the luciferase activities in the Control+FSTL1 and miR-524-5p+FSTL1-mut groups were identified. Therefore, FSTL1 is targeted by miR-524-5p.
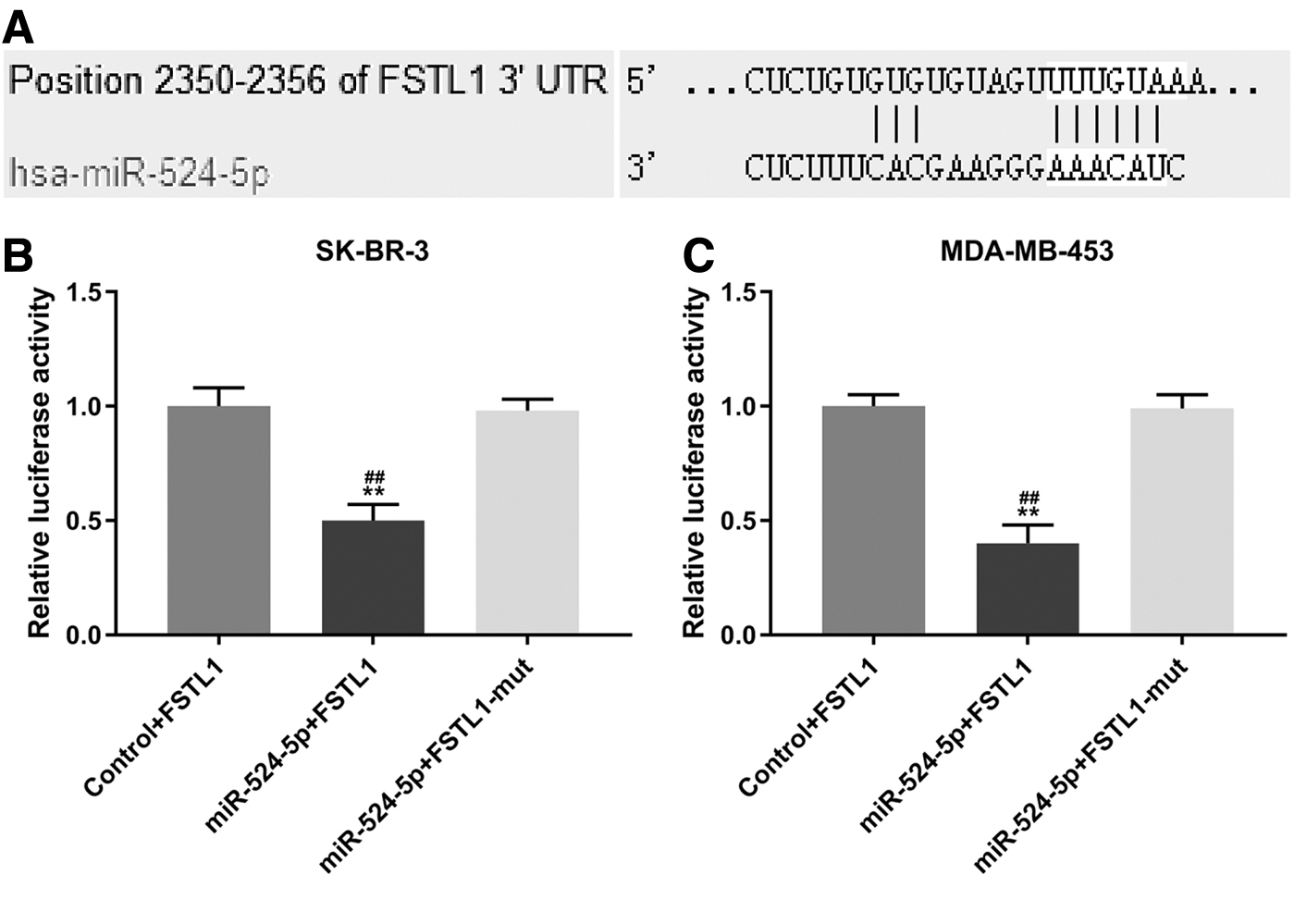
MR-524-5p specifically targets FSTL1.
Overexpressed FSTL1 partly reversed the inhibitory effects of miR-524-5p on viabilities of SK-BR-3 and MDA-MB-453 cells
We determined whether the inhibitory effects of miR-524-5p on SK-BR-3 and MDA-MB-453 cell viabilities were mediated by downregulating FSTL1, as shown in Figure 4, overexpression of FSTL1 significantly increased the viabilities of the two cells, compared with control and NC groups (p < 0.05), while miR-524-5p mimic greatly reduced the cell viability as compared with control and NC groups (48 h, p < 0.05; 72 h, p < 0.01, respectively). However, in FSTL1+mimic group, the inhibitory effect of miR-524-5p mimic was partly reversed by overexpressed FSTL1 compared with mimic group (48 h, p < 0.05; 72 h, p < 0.05 and p < 0.01, respectively).

Overexpressed FSTL1 partly reversed the inhibitory effects of miR-524-5p on viabilities of SK-BR-3 and MDA-MB-453 cells.
Overexpressed FSTL1 partly reversed the inhibitory effect of miR-524-5p on migration and invasion of SK-BR-3 and MDA-MB-453 cells
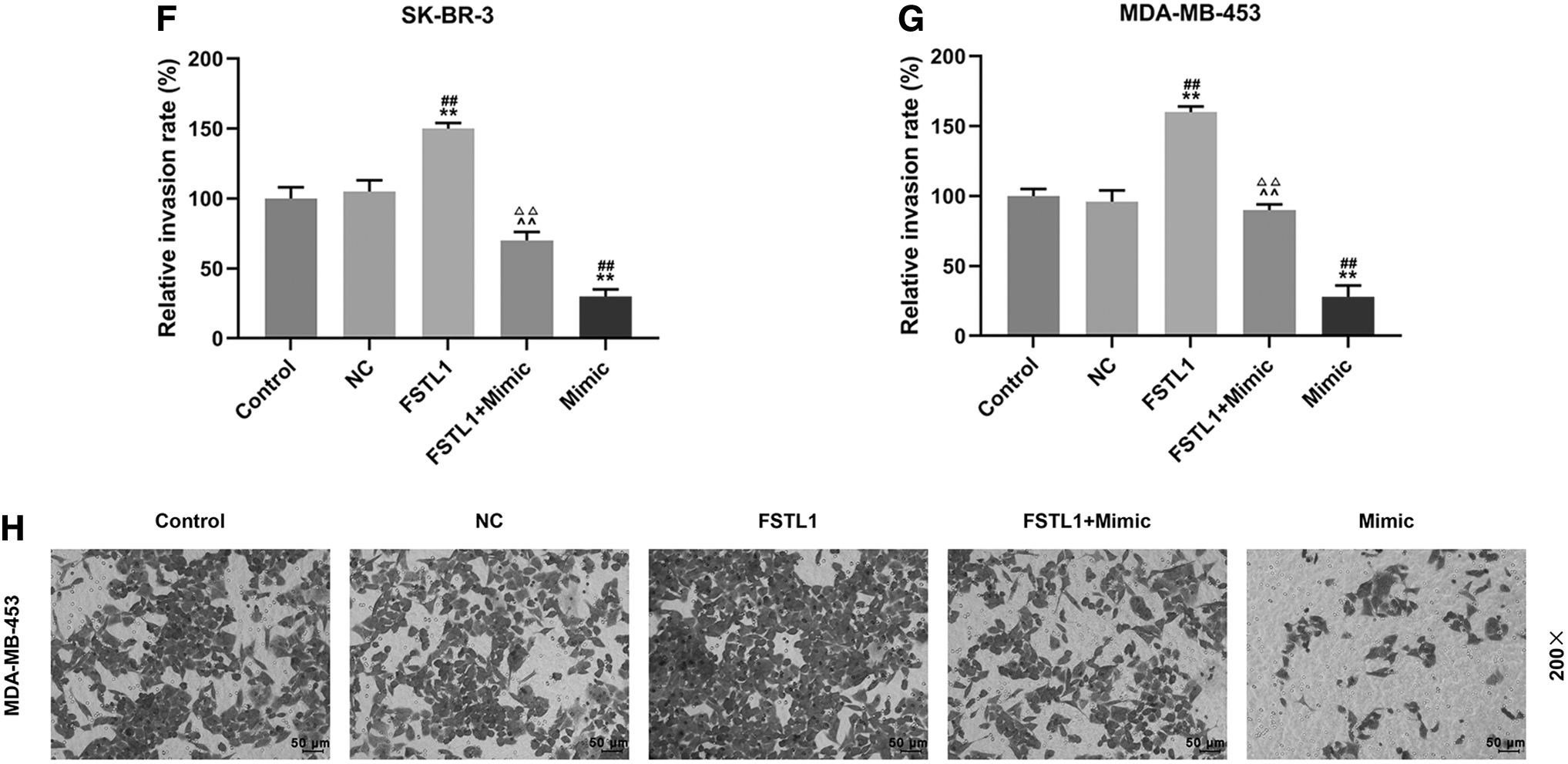
Whether the inhibitory effect of miR-524-5p on migration and invasion of SK-BR-3 and MDA-MB-453 cells was mediated through downregulating FSTL1 was determined by conducting wound healing and transwell assays. As shown in Figure 5A–D, in FSTL1 group, 24 h after the wounding experiment, the relative migration rates were increased compared with control and NC groups (p < 0.01) both in SK-BR-3 and MDA-MB-453 cells. In mimic group, the relative migration rates were reduced in the two cells as compared with control and NC groups (p < 0.01), while in FSTL1+mimic group the inhibitory effect of miR-524-5p mimic on cell migration in the two cells was partly reversed by overexpression of FSTL1 compared with FSTL1 and mimic groups (SK-BR-3, p < 0.01; MDA-MB-453, p < 0.01 and p < 0.05, respectively). As shown in Figure 5E–H, the changes in cell invasion were similar to the results in wound healing assays. In the FSTL1 group, the relative invasion rates in the two cells were significantly increased as compared with control and NC groups (p < 0.01), while in mimic group the relative invasion rates of the two cells were reduced compared with control and NC groups (p < 0.01). However, in FSTL1+mimic group, the inhibitory effect of miR-524-5p mimic on cell invasion was partly reversed by overexpressed FSTL1 in the two cells as compared with FSTL1 and mimic groups (p < 0.01). Thus, the results showed that overexpression of FSTL1 partly reversed the inhibitory effect of miR-524-5p on migration and invasion of SK-BR-3 and MDA-MB-453.
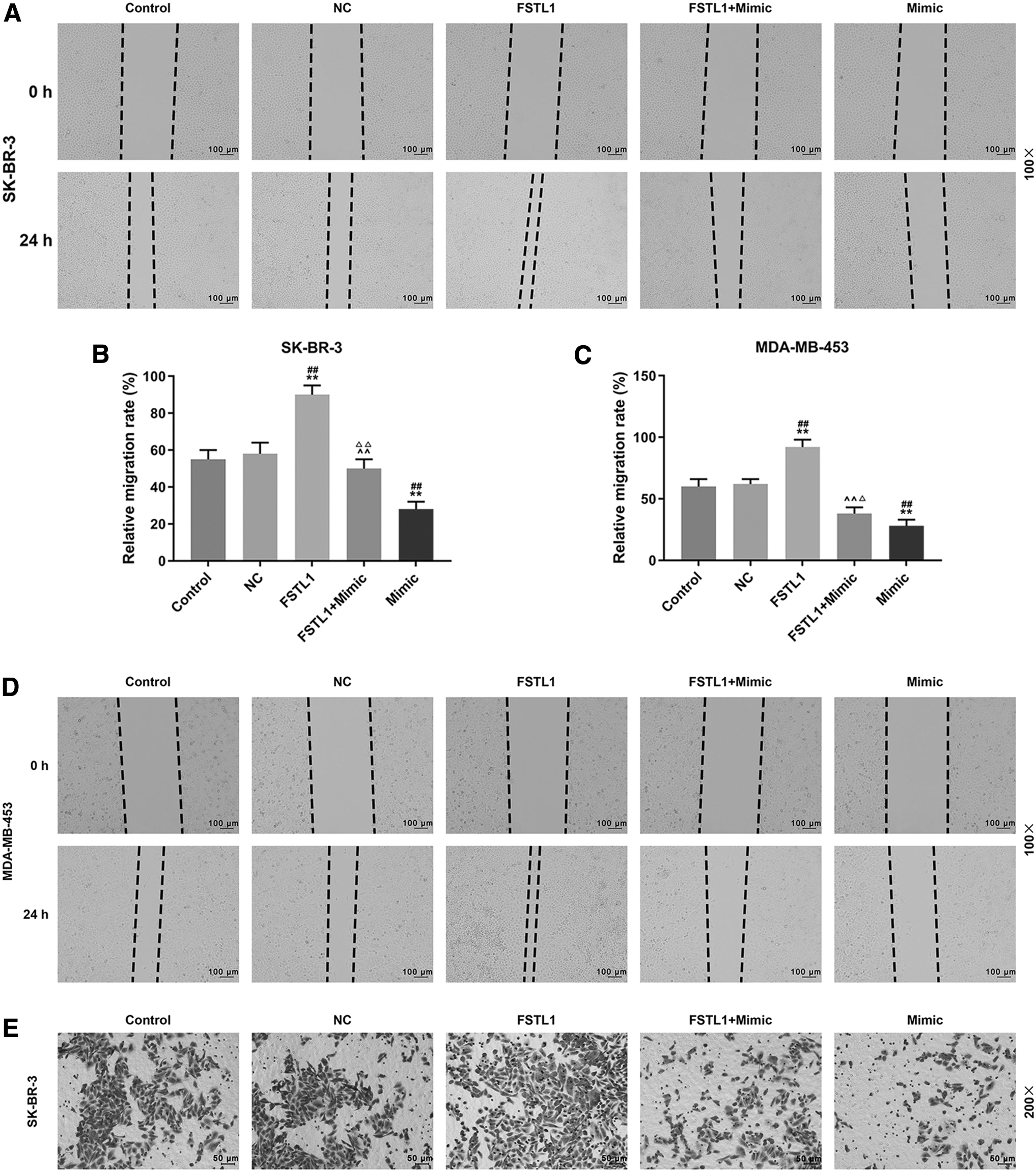
Overexpressed FSTL1 partly reversed the inhibitory effect of miR-524-5p on migration and invasion of SK-BR-3 and MDA-MB-453 cells.
Overexpressed FSTL1 partly reversed the inhibitory effect of miR-524-5p on the expression of FSTL1
The relation between miR-524-5p and FSTL1 was determined by Western blot and q-PCR assays, as shown in Figure 6A, B, and D, E, FSTL1 protein expression in SK-BR-3 and MDA-MB-453 cells was upregulated in FSTL1 group compared with control and NC groups (p < 0.01); however, FSTL1 protein was downregulated in mimic group compared with control and NC groups (p < 0.05 and p < 0.01, respectively). In FSTL1+mimic group, the expression of FSTL1 protein was lower than that in the FSTL1 group (p < 0.05) but higher than that in the mimic group (p < 0.01). The mRNA expression level of FSTL1 in the two cells after transfection with FSTL1 and/or mimic was consistent with its protein expression (Fig. 6C, F). Thus, the results showed that miR-524-5p specifically targets FSTL1.
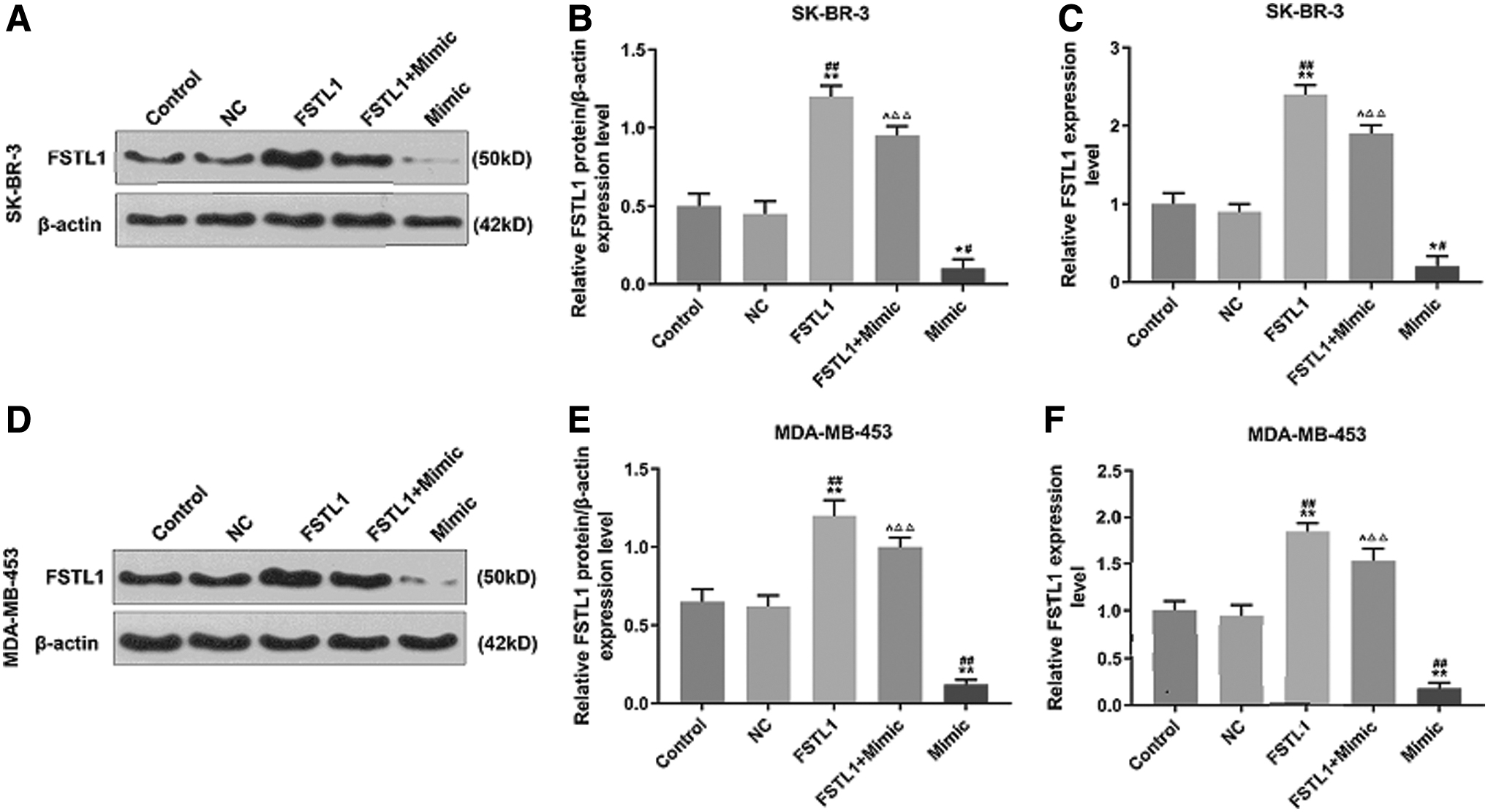
Overexpressed FSTL1 partly reversed the inhibitory effect of miR-524-5p on the expression of FSTL1.
Overexpressed FSTL1 partly reversed the inhibitory effect of miR-524-5p on EMT progression
We further detected the level of transcription and translation of some key factors involved in EMT. As shown in Figure 7A–C and E–G, in FSTL1 group, the protein expressions of MMP2, MMP9, and N-cadherin in SK-BR-3 and MDA-MB-453 cells were upregulated (p < 0.05), while the protein expression of E-cadherin was downregulated compared with control and NC groups (p < 0.01). In mimic group, the protein expressions of MMP2, MMP9, and N-cadherin in the two cells were significantly downregulated (p < 0.01), whereas the protein expression of E-cadherin was upregulated compared with control and NC groups (p < 0.01). In FSTL1+mimic group, the effects of miR-524-5p on these proteins were reversed by FSTL1 in the two cells compared with FSTL1 and mimic groups (p < 0.05), and the mRNA expression levels of these factors in the two cells were consistent with the protein expressions (Fig. 7D, E and I, J). Thus, the results proved that FSTL1 partly reversed the inhibitory effect of miR-524-5p on EMT progression, further confirming that the regulatory effects of miR-524-5p on the cell viability, migration, and invasion of SK-BR-3 and MDA-MB-453 cells were associated with the inhibition of FSTL1.

Overexpressed FSTL1 partly reversed the inhibitory effect of miR-524-5p on EMT progression.

Discussion
Cancer metastasis is a major cause of death for cancer patients. 23 Abnormal changes in cancer-related genes contribute greatly to the occurrence and development of cancer and cancer metastasis. 9,24 Researchers proved the potential values of miRNAs in cancer screening, diagnosis, and treatment. 14 –16 miRNAs are a type of small noncoding RNAs with 18–25 nucleotides in length, and they are vital regulators in regulating 30%–60% human gene expressions in the biological processes such as cell proliferation, angiogenesis, EMT, and apoptosis. 25 MiR-524-5p was recently reported to have the ability of inhibiting cancer cell growth such as melanoma and glioma. 17 –19 However, there is a lack of report on the role of miR-524-5p in BC. Thus, this study determined the role of miR-524-5p in BC, and we found that miR-524-5p was low-expressed in BC tissues compared with adjacent tissues. Such a phenomenon in this study is consistent with previous studies, 17 –19 indicating that miR-524-5p plays an important role in the progression of BC; however, the underlying mechanisms still require further investigation. Subsequently, the effects of overexpressed miR-524-5p on biological abilities of BC progression were detected, and we found that miR-524-5p remarkably inhibited the viability, migration, and invasion of BC cells, suggesting that miR-524-5p has a functional role in BC.
miRNAs can affect the pathogenesis of various diseases through specifically targeting certain mRNA. 14 In this study, to further detect the antitumor effect mechanism of miR-524-5p, the result from bioinformatics prediction showed that FSTL1 was a potential effector of miR-524-5p, which was validated by luciferase assay results. FSTL1 is involved in a series of physiological and pathological processes such as osteoarthritis, reactive arthritis, and ulcerative, 26 and it regulates biological processes, including cell proliferation, immune response, cell apoptosis, and metabolism. 26 –29 This study showed that miR-524-5p targeted FSTL1 and had an inhibitory effect on BC cells. In addition, it is reported that FSTL1 is directly regulated by miR-198 to inhibit cell migration. 30 MiR-32-5p also regulates FSTL1 to inhibit cell survival in Mycobacterium. 31 To further confirm the specific mechanism of the effect of miR-524-5p/FSTL1 signal on BC, CCK-8, wound healing, transwell, Western blot, and q-PCR assays were performed. The results showed that FSTL1 partially reversed the inhibitory effect of miR-524-5p on the viability, migration, and invasion ability of BC cells. Moreover, miR-524-5p could downregulate the expression of FSTL1 at protein and mRNA levels, suggesting that miR-524-5p could reduce the transcription and translation of FSTL1 and exert the inhibitory effect on BC cell viability, migration, and invasion.
EMT plays a vital role in the metastasis of cancer cells.
32
–36
During EMT, epithelial cells gradually transform into interstitial cells, causing cell migration and invasion.
37,38
EMT process can be characterized by decreased expressions of adhesion molecules (e.g., E-cadherin) and increased expression of N-cadherin.
39,40
MMP2 and MMP9 are EMT markers that could be increased significantly during EMT process.
41,42
Therefore, changes in EMT-related gene expressions serve as important markers in indicating the degree of metastasis in tumor cells. In the present research, our results showed that miR-524-5p/FSTL1 could significantly regulate migration and invasion of BC cells. Furthermore, we detected expressions of relative key factors at protein and mRNA levels, and found that miR-524-5p downregulated MMP2, MMP9, and N-cadherin expressions, upregulated the expression of E-cadherin at protein and mRNA levels. However, FSTL1 produced the opposite effect to miR-524-5p. The data also revealed that FSTL1 could partly reverse the function of miR-524-5p in the expressions of above factors, which further proved that FSTL1 was targeted and inhibited by
This study found that miR-524-5p attenuated the progression of migration, invasion, and EMT in BC cells by downregulating FSTL1 expression. However, the mechanisms through which the inhibitory effect of miR-524-5p on viability and EMT of BC cells was realized should be further determined, such as a previous research figured that miR-524-5p had a relationship with Wnt signaling pathway. 43 Therefore, it might be developed as a new research for investigating the further mechanisms of miR-524-5p on cancer cells.
Conclusions
In conclusion, the research results indicate that miR-524-5p attenuates the progression of migration, invasion, and EMT in BC cells by downregulating FSTL1 expression; therefore, miR-524-5p could be served as a therapeutic target for BC treatment.
Footnotes
Authors' Contributions
T.J. and Y.Z. made substantial contributions to conception and design; TJ, YZ and TZ performed data acquisition, data analysis, and interpretation; TJ and YZ drafted the article or critically revising it for important intellectual content; all authors gave their final approval of the version to be published; T.Z prepared the agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.