
Abstract
Background:
Bioinformatics showed that long non-coding RNA (lncRNA) pgm5-as1 was regulated in patients with colorectal cancer (CRC), and miR-484 was also regulated in CRC. We aimed at determining the modulatory pathway of lncRNA pgm5-as1 in CRC cells, and whether miR-484 was involved in the pathway.
Materials and Methods:
The target gene of pgm5-as1 was predicted by bioinformatics and verified by dual luciferase assay. Transcription levels of pgm5-as1 and miR-484 were determined by quantitative real-time polymerase chain reaction. Viability, migration rate, invasion, and growth of SW480 and HCT116 cells were determined by Cell Counting Kit-8 (CCK-8), wound healing assay, transwell, and colony formation assay, respectively.
Results:
pgm5-as1 was upregulated in CRC tissues and cell lines; however, its downregulation contributed to the decreasing of cell viability, growth, migration, and invasion of SW480 and HCT116 cells. Moreover, miR-484 was predicted as the target of pgm5-as1, and the downregulation of pgm5-as1 partially restored the elevated cell viability, growth, migration, and invasion that were induced by the inhibition of miR-484 expression in SW480 and HCT116 cells.
Conclusions:
The loss of miR-484 expression in CRC might be involved in the promotion and metastasis of CRC, which may be caused by the overexpression of pgm5-as1. Hence, the downregulation of pgm5-as1 could be a therapeutic target in the prevention or intervention of CRC.
Introduction
Colorectal cancer (CRC) is one of the most common malignant tumors of the digestive system. 1,2 Recently, the morbidity and mortality of CRC are increasing rapidly in many less developed regions. 1 According to the epidemiological report in China, CRC incidence is the third highest among all malignant tumors and the mortality rate is the fourth highest. 1 One of the most important causes of poor prognosis in CRC is metastasis, 3 and about 25% of CRC patients have metastasis at the time of their diagnosis; moreover, ∼50% of CRC patients suffer from metastasis over time. 3 At present, the treatment of CRC is largely based on surgical resection. 4 However, due to the complexity of CRC etiology, and its high malignancy, rapid growth, and strong invasion ability, 5-year overall survival rate of the disease is still low. 5 Therefore, further studies should be conducted to find new therapeutic targets and treatment pathways that can further prolong the survival time for CRC patients.
Long non-coding RNAs (lncRNAs) are a type of RNA molecules located in the nucleus or cytoplasm, with transcript length longer than 200 nt. 6,7 lncRNAs do not encode proteins, and most of them are produced through transcribing RNA polymerase II and thereafter the alternatively splicing. 6,8 RNAs regulate expression levels of target genes at epigenetic, transcriptional, and post-transcriptional levels. 6 Studies have found that lncRNAs are involved in many regulatory progresses, such as chromatin modification, DNA methylation, histone modification, genomic imprinting, transcriptional activation, transcriptional interference, and nuclear transport. 9 –12 lncRNAs are considered to be closely linked to the mechanism of diseases, including cancer, 13 as abnormally expressed lncRNA can be found in the lesions, blood, and other specimens of CRC patients. 14,15
New types of lncRNAs related to CRC have been discovered and their regulatory roles in the occurrence, development, and invasion of CRC have been further elucidated as well. 16,17 The abnormality of lncRNA expression now served as a new biological indicator for CRC prevention, diagnosis, prognosis, and treatment. 18 –20 Recently, Zhu et al. revealed that p53-induced lncRNA (pgm5-as1) was upregulated between the stage II and III of CRC. 21 However, a few researches demonstrated the potential of pgm5-as1 as a target for CRC treatment and the mechanism of the lncRNA in CRC progression. Thus, this study aimed at exploring the underlying modulatory pathway of lncRNA pgm5-as1 in CRC cells, expecting to find diagnostic or therapeutic biomarkers for CRC treatment.
Materials and Methods
The ethics
The experiments and sample collection in this study were conducted and approved by the Ethic Board of Hubei Provinical Hospital of Integrated Chinese and Western Medicine. Twenty patients in stage I and II and 20 patients in stage III and IV whose tissues were extracted all signed the informed consent.
Cell culture
HIEC, SW480, HT29, HCT116, Caco-2, and LoVo cell lines were purchased from Shanghai Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences (CAS). HIEC, SW480, HT29, HCT116, and Caco-2 cells were cultured in DMEM (D0819; Sigma-Aldrich, St. Louis, MO), whereas LoVo cell line was cultured in Ham's F-12K (21127030; Thermo Fisher, Waltham, MA). Ten percent fetal bovine serum (FBS, F8192; Sigma-Aldrich) was added into all the cell lines. The cells were cultured with 5% CO2 at 37°C.
Grouping
SW480 and HCT116 cells were divided into Blank, siNegative Control (siNC), and sipgm5-as1 groups. The cells in the Blank group were treated by medium only, those in the siNC group were transfected with siNC (GenePharma, Shanghai, China), and those in the sipgm5-as1 group were transfected with sipgm5-as1 (GenePharma). To determine the roles of miR-484 and pgm5-as1 in CRC, SW480 and HCT116 cells were grouped into Mimic control, Mimic, Inhibitor control and Inhibitor, and sipgm5-as1+inhibitor groups, which were, respectively, transfected with miR-484 mimic control (scrambled sequence for the mimic of miR-484), miR-484 mimic (sequence to increase function of miR-484), miR-484 inhibitor control (scrambled sequence for the inhibitor of miR-484), miR-484 inhibitor (sequence to inhibit function of miR-484), and miR-484 inhibitor and sipgm5-as1 (GenePharma). The transfection was conducted following the instruction of Lipofectamine 3000 (L3000015; Thermo Fisher).
Quantitative real-time polymerase chain reaction
Total RNAs were extracted from the tissues collected from the CRC patients by using TRIzol reagent (15596018; Thermo Fisher). The total RNAs in SW480 and HCT116 cells (1 × 106 cells) were extracted as aforementioned. cDNAs complemented to the RNAs were reverse-transcribed by PrimeScript RT reagent kit (Takara Biotechnology Co., Ltd., Dalian, China). Twelve microliters 2 × SYBER Green master mix (4913850001; Roche, Shanghai, China), 1 μL forward primer (10 μM), 1 μL reverse primer, 5 μL cDNA template, and 7 μL ddH2O were thoroughly mixed together. Polymerase chain reaction was conducted at 95°C for 90 s, at 95°C for 30 s, at 68°C for 30 s, and at 72°C for 35 s, for 40 cycles in Bio-Rad IQ5 thermocycler (Bio-Rad, CA). MRNA levels were calculated by 2-ΔΔCt method. All primers used are listed in Table 1.
Primers Used in the Study
Cell Counting Kit-8
The cell viability was determined after sipgm5-as1 had been transfected into SW480 and HCT116 cells for 24, 48, and 72 h. Viabilities of the cells were measured after 48 h when miR-484 mimic, miR-484 inhibitor, or sipgm5-as1 was transfected as described earlier. Briefly, after 5 × 105 cells had been prepared, Cell Counting Kit-8 (CCK-8) (96992-100TESTS-F; Sigma-Aldrich) was performed. Optical density of each sample was determined on a microplate reader (Multiskan; Thermo, Waltham, MA) at 450 nm.
Colony formation assay
Briefly, 1 × 106/mL SW480 and HCT116 cells were collected and seeded at about 500 cells/well in 2 mL 1.2% cooled agarose in a mixture with culture medium at 1:1. Thereafter, the cells were incubated for another 14 d and then stained by 0.5% crystal violet. After washing, colony numbers of cells were counted under a microscope. The experiment was conducted in triplicate.
Transwell
SW480 and HCT116 cells (2 × 106) were collected. Matrigel-coated membrane (BD Biosciences, San Jose, CA) was coated in the interlayer of a Transwell (8-μm; BD Biosciences). The cells were pipetted into the upper chamber of Transwell, and 400 μL 10% FBS was added into the bottom chamber. The cells were incubated with 5% CO2 at 37°C for 48 h and stained by 0.2% crystal violet to count invaded cells from the upper chamber.
Wound healing assay
SW480 and HCT116 cells (2 × 106) were collected and seeded into a plate to produce a monolayer of cells. Then, a scratch was created in the middle of the monolayer by using a 200-μL pipette. The cells were then incubated with culture medium in 5% CO2 at 37°C for 48 h. The relative distance was calculated according to the mean value of the distance on the top, middle, and bottom.
Dual luciferase assay
LncBase Predicted v.2 predicted that pgm5-as1 and miR-484 share a possible complementary sequence. To further confirm the prediction, a mutant sequence of pgm5-as1 was created by using Quick-Change Site-Directed Mutagenesis kit (Stratagene, CA). All transfection procedures were performed following the instruction of Lipofectamine 3000 (L3000015; Thermo Fisher). The cells were co-transfected with miR-484 inhibitor and pRL-TK with mutated pgm5-as1 or not (Promega, WI), and they were incubated for 48 h. Luciferase activity was detected by using Dual-Glo luciferase assay kit (Promega).
Statistics
The data collected were statistically analyzed in OriginPro 2017, with n = 5 for each cell experiment. One-way analysis of variance was performed in the comparison of multiple groups. Turkey test was used for the comparison between two groups. A p-value of less than 0.05 was considered to be statistically significant.
Results
pgm5-as1 was upregulated in CRC tissues and cell lines, but its downregulation led to the decrease in the proliferation of cancer cells
The expression of pgm5-as1 was detected in CRC tissues at different stages and in CRC cell lines, including HIEC, SW480, HT29, HCT116, Caco-2, and LoVo cells. To determine the role of pgm5-as1 in CRC, the expression of pgm5-as1 was inhibited in SW480 and HCT116 cells, and proliferation of the cells was measured by CCK-8 and colony formation assay. We found that the relative pgm5-as1 expression in CRC tissues derived from patients in stage I and II was higher than that in normal tissue; meanwhile, the relative pgm5-as1 expression in stage III and IV was significantly increased compared with stage I and II (Fig. 1A, ^^ p, && p < 0.01). Moreover, the relative expression of pgm5-as1 was increased in SW480, HT29, HCT116, Caco-2, and lovo cells compared with that in HIEC cells, and the highest expression was observed in SW480 and HCT116 cells (Fig. 1B, Δ p < 0.05, ΔΔ p < 0.01); thus, SW480 and HCT116 cells were selected to be used in further studies. After the successful transfection of sipgm5-as1 in SW480 and HCT116 cells in the sipgm5-as1 group, the expression of pgm5-as1 was significantly reduced (Fig. 1C, **p, ## p < 0.01). Moreover, the cell viabilities of SW480 or HCT116 cells at 48 or 72 h in the sipgm5-as1 group were lower than those in the siNC or Blank group (Fig. 1D, E, **p, ## p < 0.01). Consistently, the relative colony numbers of SW480 and HCT116 cells were decreased in the sipgm5-as1 group compared with those in the siNC or Blank group (Fig. 1F–I, **p, ## p < 0.01). Taken together, pgm5-as1 expression was increased in tumor and cell lines, and the downregulation of lncRNA pgm5-as1 caused inhibition of CRC cells proliferation.
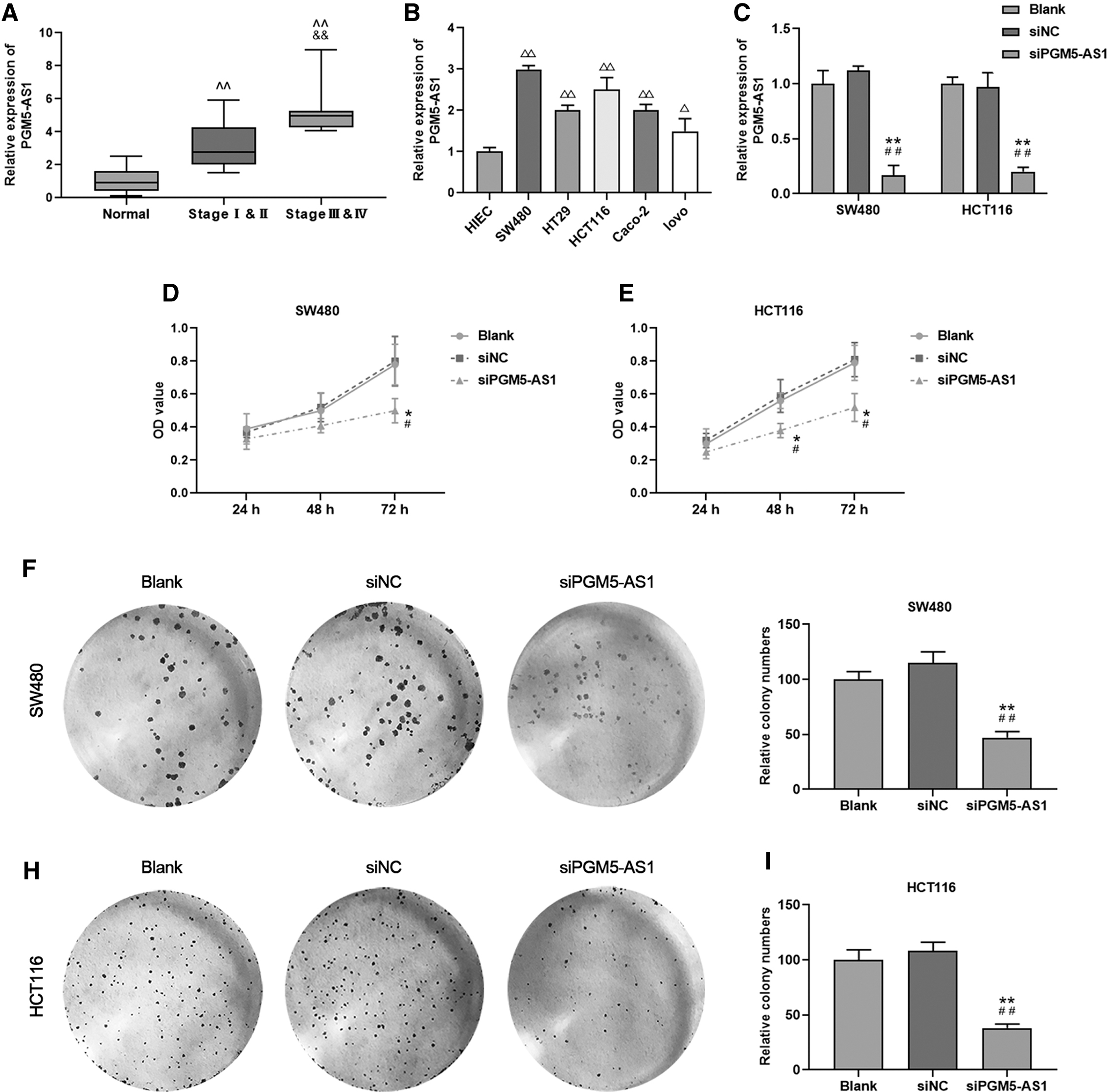
lncRNA pgm5-as1 expression in CRC tissue and cell lines whereas its downregulation contributed to the decreasing of proliferation.
The downregulation of pgm5-as1 reduced the migration and invasion abilities of CRC cell lines
To further understand the effect of the downregulation of pgm5-as1 on the potential of metastasis of CRC cells, cell migration and invasion abilities were determined under the condition of downregulated pgm5-as1 in SW480 and HCT116 cells in the sipgm5-as1 group. We found that the relative migration and invasion rates in SW480 and HCT116 cell lines in the sipgm5-as1 group were significantly lower than those in the insiNC or Blank group (Fig. 2A–H, *p, # p < 0.05, **p, ## p < 0.01). Thus, the downregulation of pgm5-as1 could inhibit the metastasis potential of CRC cells.

The downregulation of pgm5-as1 led to the decrease of migration and invasion rates.
miR-484 was the target of pgm5-as1
To explore the downstream miRNA that pgm5-as1 worked with, we predicted the target of pgm5-as1 as well as investigated the correlation between the expressions of pgm5-as1 and its target miRNA. Interestingly, there were possible complementary sequences between pgm5-as1 and miR-484 expression (Fig. 3A), and the relative luciferase activity in the Inhibitor group was significantly higher than that in the Blank group when pgm5-as1 sequence was not mutant in both SW480 and HCT116 cell lines (Fig. 3B, C, **p < 0.01), indicating the combination of pgm5-as1 and miR-484. In addition, the relative expression of miR-484 in SW480 and HCT116 cells was elevated when pgm5-as1 was silenced in the sipgm5-as1 group, compared with that in the siNC or Blank group (Fig. 3D, E, **p < 0.01). The results suggested that pgm5-as1 could interact with miR-484, and the expression of the two non-coding RNAs was negatively correlated in CRC cells.

miR-484 was the target of pgm5-as1.
The downregulation of pgm5-as1 partially restored the elevated proliferation capacity by the inhibition of miR-484 expression
To explore whether the interaction between pgm5-as1 and miR-484 was involved in CRC progression, the changes in the proliferation of SW480 and HCT116 cells with or without regulation of pgm5-as1 or miR-484 were determined. Here, we revealed that the relative expression of miR-484 in SW480 and HCT116 cells in the Mimic group was significantly higher than that in the Mimic control group (Fig. 4A, B, **p < 0.01). However, there was no statistical difference in miR-484 expression between Inhibitor control and Inhibitor groups (Fig. 4A, B), indicating that miR-484 could not be greatly inhibited in CRC cells. Surprisingly, in SW480 and HCT116 cells, the cell viability and relative colony numbers were reduced greatly in the Mimic group compared with those in the Mimic control group, but they were elevated in the Inhibitor group than in the Inhibitor control group (Fig. 4C–H, *p, # p, ^p < 0.05, **p, ## p, ^^ p < 0.01). Moreover, cell viability and relative colony numbers of SW480 and HCT116 cells were reduced in the sipgm5-as1+Inhibitor group compared with the Inhibitor group (Fig. 4C–H, *p, # p, ^p < 0.05, **p, ## p, ^^ p < 0.01). Thus, the findings suggested that miR-484 could act as a suppressor to CRC cells, and such an effect could be counteracted by pgm5-as1.

The downregulation of pgm5-as1 partially restored the elevated proliferation capacity after the inhibition of miR-484 expression.
The downregulation of pgm5-as1 partially offset the increased metastasis of CRC cells induced by the inhibition of miR-484 expression
We investigated the effect of the interaction between pgm5-as1 and miR-484 on the migration and invasion of CRC cells, and the data demonstrated that the relative migration and invasion rates were significantly reduced in the Mimic group compared with the Mimic control group, but they were increased in SW480 and HCT116 cells in the Inhibitor group compared with the Inhibitor control group (Fig. 5A–H, *p < 0.05, **p, ## p < 0.01). More importantly, relative migration and invasion rates of SW480 and HCT116 cells were significantly reduced in the sipgm5-as1+Inhibitor group compared with the Inhibitor group (Fig. 5A–H, ^^ p < 0.01). Thus, these results suggested that the metastasis potential of CRC cells could be promoted by the downregulation of miR-484; however, the downregulation of pgm5-as1 could offset such an effect partially.
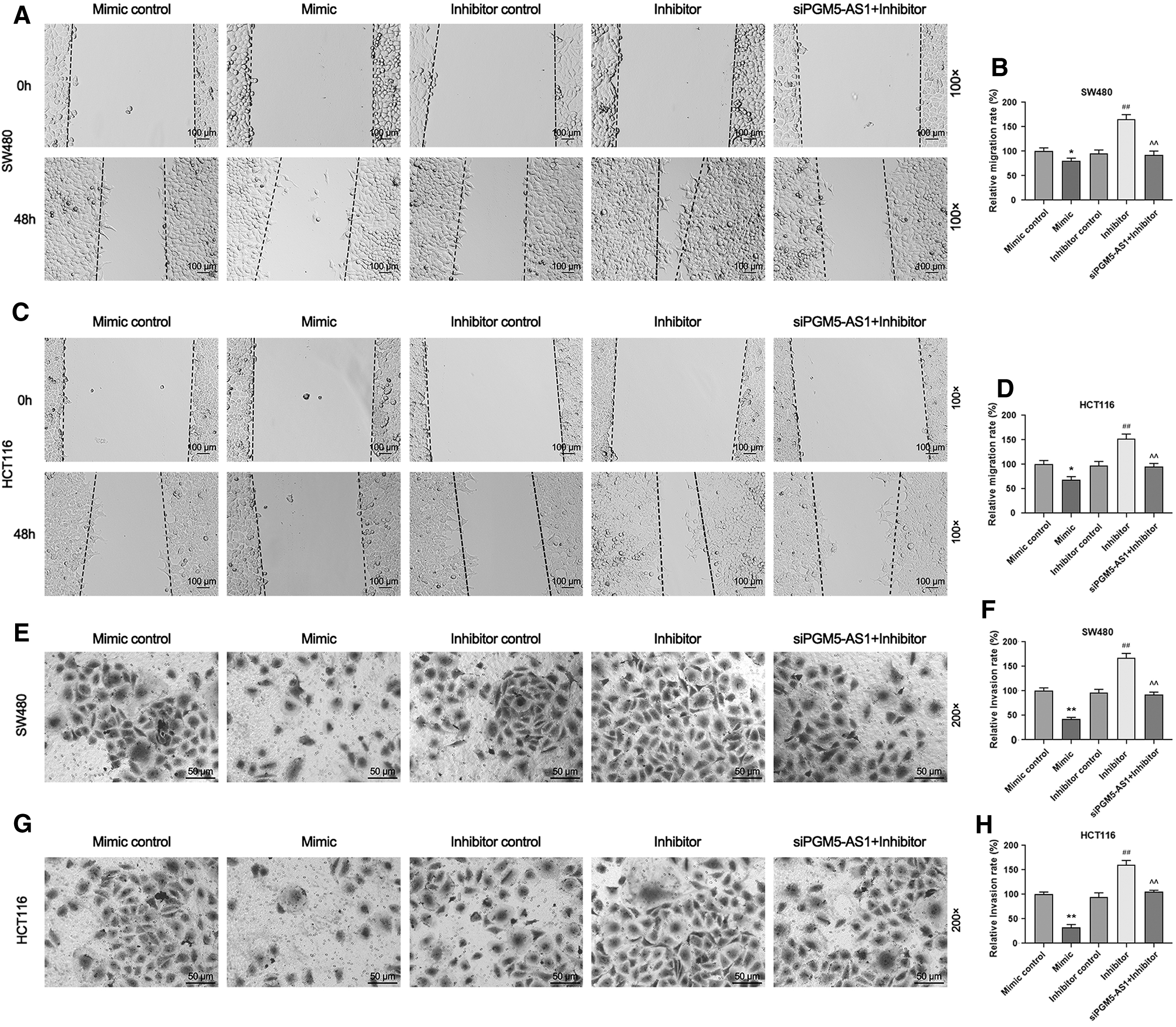
The downregulation of pgm5-as1 partially offset the elevated capability of metastasis of CRC cells after the inhibition of miR-484 expression.
Discussion
This study revealed that pgm5-as1 was upregulated during CRC progression, and the downregulation of pgm5-as1 led to the decreasing of CRC proliferation, migration, and invasion abilities. Further, miR-484 was predicted as the target of pgm5-as1, and the expression of miR-484 was negatively related with the proliferation, migration, and invasion of CRC cells. However, the downregulation of pgm5-as1 could counteract with the inhibition of miR-484, and the regulatory pathway discovered in this study contributes to the intervention of CRC cell progression.
pgm5-as1, which could promote CRC procession, was found to be increased in CRC tissue and cell lines. Specifically, downregulated pgm5-as1 inhibited the proliferation of SW480 and HCT116 cells compared with HIEC cells. Moreover, migration and invasion rates of the cells demonstrated consistent results to those of the proliferation. Zhu et al. noted that pgm5-as1 in the samples from the CRC patients in stage III and IV were upregulated and positively correlated with the unfavorable overall survival rate. 21 pgm5-as1 overexpression significantly suppressed the proliferation, migration, and invasion of esophageal squamous cell carcinoma cells in vitro and tumor growth in vivo. Thus, the expression of pgm5-as1 should be further determined. From the findings in the current and previous studies, we speculated that pgm5-as1 might be a cancer-promoting lncRNA during CRC progression.
Further, miR-484 was found to be negatively related to the proliferation, migration, and invasion of CRC in SW480 and HCT116 cells. Xie et al. showed that miR-484, which could be sponged by lncRNA ZFAS1, played a negative role in the CRC process.
22
lncRNA H19 promotes epithelial
This study found complementary sequences between pgm5-as1 and miR-484 through bioinformatics; moreover, dual luciferase assay data revealed that pgm5-as1 could interact with miR-484. 25,26 Further, the expression of pgm5-as1 and miR-484 was found to be negatively correlated, and the downregulation of pgm5-as1 counteracted the effect of inhibiting miR-484 on the proliferation, migration, and invasion of SW480 and HCT116 cells to some extent. Under normal circumstances, lncRNAs exert regulatory effects through interacting with miRNA. 10,27,28 Thus, it could be hypothesized that miR-484 could be regulated by pgm5-as1 and, therefore, plays a role in the development of CRC.
Conclusions
To conclude, the loss of miR-484 expression in CRC might be involved in the promotion and metastasis of CRC, which may be the result of overexpressed pgm5-as1. Hence, the downregulation of pgm5-as1 could serve as a therapeutic target in the prevention or intervention of CRC. However, the role of pgm5-as1 still needs to be determined.
Footnotes
Authors' Contributions
Substantial contributions to conception and design: Y.S., L.Q.; Data acquisition, data analysis, and interpretation: Y.L., Y.Z.; Drafting the article or critically revising it for important intellectual content: X.G., Y.Z.; Final approval of the version to be published: All authors; Agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved: K.W.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.