
Abstract
Background:
Prostate-specific membrane antigen (PSMA) has emerged as a promising target for developing radionuclide therapy (RNT) in prostate cancer; however, accumulation of PSMA-RNT in salivary glands can result in irreversible xerostomia. Methods to prevent PSMA-RNT-related xerostomia could be clinically useful; however, little is known about PSMA expression in salivary glands of preclinical animal models. Using [18F]DCFPyL autoradiography/biodistribution, PSMA expression levels were determined in salivary glands of various preclinical monkey and rodent species and compared with humans.
Methods:
Binding affinities (Kd) and PSMA levels (Bmax) were determined by in vitro [18F]DCFPyL autoradiography studies. In vivo rodent tissue uptakes (%ID/g) were determined from [18F]DCFPyL biodistributions.
Results:
[18F]DCFPyL exhibited low nanomolar Kd for submandibular gland (SMG) PSMA across all the species. PSMA levels in human SMG (Bmax = 60.91 nM) were approximately two-fold lower compared with baboon SMG but were two- to three-fold higher than SMG PSMA levels of cynomolgus and rhesus. Rodents had the lowest SMG PSMA levels, with the mouse being 10-fold higher than the rat. In vivo rodent biodistribution studies confirmed these results.
Conclusions:
SMG of monkeys exhibited comparable PSMA expression to human SMG whereas rodents were lower. However, the results suggest that mice are relatively a better small animal preclinical model than rats for PSMA salivary gland studies.
Introduction
Prostate-specific membrane antigen (PSMA), also known as glutamate carboxypeptidase II or folate hydrolase 1, is encoded by the FOLH1 gene. 1 PSMA is overexpressed on the cell membrane of most primary and metastatic prostate cancers (PCa) and in the neovasculature of solid tumors. 2,3 PSMA has remarkably high expression levels in PCa cells and low expression in healthy tissues except for kidneys, salivary glands, and lacrimal glands. Hence, PSMA has emerged as a promising target for radionuclide therapy (RNT) for metastatic PCa. 4 There is increasing evidence of clinically meaningful responses to PSMA-targeted RNT labeled by using both β′ (177Lutetium; 177Lu) and α (225Actinium; 225Ac) emitting radionuclides. 5,6 Several PCa patients, who failed earlier therapies, showed positive responses to 225Ac-labeled PSMA, including radiographic and complete biochemical responses. 7,8 Despite this significant progress, PSMA-targeted RNT can cause damage to salivary glands and lacrimal glands, resulting in dry mouth (xerostomia) and dry eyes (xerophthalmia), respectively. 9 –11 To design effective strategies to protect these tissues from PSMA-targeted RNT, a better understanding of PSMA expression and its regulation in salivary and lacrimal glands is needed.
The salivary complex includes the parotid, submandibular, and sublingual glands, as well as many minor (accessory) salivary glands located throughout the oral cavity and upper airway. The salivary glands are complex organs composed chiefly of two epithelial cell types: acinar and ductal epithelium. 12 The PSMA is exclusively expressed on the apical lumen of the acinar epithelium in all types of salivary glands in humans. Although PSMA is well known for its carboxypeptidase and folate hydrolase activities and likely plays a role in secretion, its exact role in salivary gland function remains unknown. 13 –15 In general, the acinar epithelium is responsible for the production of a plasma-like primary saliva product composed of salivary proteins. 16 In humans, the acinar epithelium is particularly sensitive to damage from ionizing radiation. Moreover, acinar cells have poor regenerative capacity and a stem cell niche has not been clearly identified in humans. 9,17 –19 In patients, PSMA-mediated salivary gland accumulation of RNT agents and the associated ionizing radiation can result in irreversible damage to the acini, leading to reduced secretory function and atrophy of the tissue. The resultant salivary hypofunction from RNT has a profound negative impact on quality of life, including oral function (e.g., difficulty masticating, swallowing, and speaking), impaired sense of taste, tooth decay, and oral infections (e.g., oral candidiasis, periodontal disease). 10,11,20 –22 Likewise, the permanence of xerostomia caused by PSMA-targeted α-emitting (225Actinium), and β′-emitting radionuclides (177Lutetium) has been shown to be dose dependent. 5,10,23,24 This unwanted and often irreversible side-effect has hindered the rapid development of PSMA-targeted alpha RNT. Because of the high clinical importance of PSMA-targeted RNT in treating PCa, there is an urgent need to develop suitable preclinical models to reliably investigate strategies to protect salivary glands from radiation-induced damage.
Through proof-of-concept preclinical studies, efficacious clinical studies aimed at preventing uptake of radionuclides may be achievable. 25,26 Proper animal models will play a vital role in establishing methods to protect the salivary glands; however, little is known about PSMA expression levels in salivary glands of commonly used laboratory animals. Identification of preclinical animal models with appropriate levels of PSMA will expedite testing of various strategies to protect the salivary glands from PSMA-targeted RNT. Herein, we determined and compared the concentration of PSMA in salivary glands across various species by using 2-(3-{1-carboxy-5-[(6-[ 18 F]fluoro-pyridine-3-carbonyl)-amino]-pentyl}-ureido)-pentanedioic acid, [ 18 F]DCFPyL 27 (Fig. 1A), a synthetic radioligand exhibiting high affinity and specificity for PSMA via autoradiography and rodent biodistribution studies, and correlated these findings with immunofluorescence (IF) confocal microscopy.

Materials and Methods
PSMA conservation across common animal models
Protein sequence conservation was compared across human, mouse, rat, cynomolgus, and rhesus species by using a multiple sequence alignment program (Clustal Omega;
Tissue samples
Fresh healthy salivary glands were collected from two cynomolgus monkeys, two rhesus monkeys, three mice (NCR-nu/nu athymic), and three rats (F344/SAS Fischer); fast frozen; and stored (−80°C) until use. Fresh frozen salivary glands from two humans (Human Cooperative Tissue Network) and two baboons (Bioreclamation IVT) were obtained and stored (−80°C) until use. All human tissues had been examined grossly by a board-certified pathologist (B.M.W.), portioned, either formalin-fixed or embedded in OCT, snap frozen in an isopentane/liquid nitrogen slurry, and stored at −80°C until use. PC3-PSMA tumors (prostate cancer cell line transfected with human PSMA obtained from a mouse xenograft) were fast frozen to be used as a positive control.
Synthesis and radiolabeling of [ 18 F]DCFPyL
[ 18 F]DCFPyL was prepared according to our recently reported literature method with a radiochemical yield of 32%–43% (decay corrected, n > 50) and purity >98% (Fig. 1B) in a 45 min synthesis time. 27 The molar activity was 1200–2600 Ci/mmol (44,000–96,200 GBq; end of synthesis).
In vitro autoradiography
Frozen submandibular salivary glands (SMG) of mice, rats, baboons, cynomolgus monkeys, rhesus monkeys, and humans were cryosectioned at 20 μm thickness (Leica CM3050S) and mounted on glass slides followed by air drying. Saturation assays were performed with mounted slides that were pre-incubated in the assay buffer [0.1% BSA, 0.1% aprotinin (100 U/mL), 50 mM Tris (pH 7.5), 1.5 mM EDTA, 150 mM saline (0.9%), 5 mM MgCl2] for 15 min at room temperature, which were incubated (2 h) with increasing concentrations of [ 18 F]DCFPyL (0.3 nM–36 nM) representing total bound activity (Bt). Non-specific binding (Bnsp) was determined by incubating the tissue sections with [ 18 F]DCFPyL (0.3 nM–36 nM) in the presence of the non-radioactive PSMA ligand DCFPyL (1 μM) at the same concentrations. Afterward, incubation slides were washed twice with Tris 50 mM at 4°C for 4 min, dipped once in cold (4°C) distilled water, and allowed to dry. After drying, the slides were exposed to phosphor imaging plates (Fuji BAS-SR2025) for 24 h. After exposure, the plates were scanned by using the Fuji FLA-5100 scanner to produce digital images from which regions of interest [ROIs; phosphostimulated luminescence units per mm2 (PSL/mm2); Image Gauge 4.0)] were drawn to determine specific bound (Bsp; Bt-Bnsp = Bsp). ROIs (PSL/mm2) were converted to molar concentrations by using specific activity of [ 18 F]DCFPyL, as previously described. 32 Briefly, selected slides were counted on a gamma counter (PerkinElmer 2480 Wizard3) to determine the radioactive content (counts per minute, CPM) and then exposed to the phosphor imaging plate from which PSL/mm2 units were determined by drawing ROIs around the entire slide. Using the linear relationship of PSL/mm2 to the CPM determined from the entire slide (CPM = slope × PSL/mm2), the ROIs from each tissue section (20-μm thickness) could be converted from PSL/mm2 to CPM/mm2 and used to calculate relative PSMA molar concentrations in the salivary gland sections. Using the molar concentrations (Bsp), the Kd and Bmax were derived for each species. Data were analyzed by using a non-linear regression curve fit, including one-site hyperbola (GraphPad Prism 7).
In vivo biodistribution
Biodistribution studies were performed in rats [F344/SAS Fischer (Strain code: 403, Charles River), female; n = 5] and mice [NCR-nu/nu athymic (Strain code: 553, Charles River), female; n = 5] at 1 h post-injection (intravenous) of [
18
F]DCFPyL [Rat:3.7 MBq (100 μCi); Mouse:1.85 MBq (50 μCi)]. After 1 h, the animals were euthanized (CO2 asphyxiation); blood/tissues from each animal were excised; the animals were weighed; and radioactive content was determined by using a gamma counter (Counting efficiency 52.4%, 400–1200 Kev). The radioactive content (CPM) of the tissue samples were corrected for background (by counting an empty vial) and all samples were decay corrected to the start time of the sample counting, including the radioactive dose. For each tissue, the percent injected dose per gram (%ID/g) was determined by using the following equations: %ID/g = {[CPM tissue/CPM total injected dose]/[tissue weight (g)]} × 100 %ID/g (normalized to a 20g mouse) = (%ID/g) × [(body weight of mice)/(20 g)] %ID/g (normalized to a 125g rat) = (%ID/g) × [(body weight of rat)/(125 g)]
Tissue to blood and tissue to muscle ratios were calculated as Tissue (%ID/g)/Blood (%ID/g) and Tissue (%ID/g)/Muscle (%ID/g), respectively. All animal studies were performed in accordance with IACUC-approved protocols.
Immunofluorescent staining
Sections (5 μm) were cut from PC3-PSMA xenograft, human, baboon, and mouse SMG. Sections were stained with anti-PSMA antibodies (human monoclonal, Novus Biologics) for PC3-PSMA, human, and baboon, and with mouse monoclonal antibody-FITC (LifeSpan Biologicals) for mouse and counterstained with DAPI. Anti-human PSMA monoclonal antibody was detected by using an anti-mouse secondary (Alexa488) at 1:200 concentration. Sections were imaged by using an Olympus Fluoview confocal microscope (15 z-stack, 40 × objective).
Results
PSMA sequence conservation across species
PSMA orthologs from all species demonstrated high sequence identity (81.1%) (Supplementary Fig. S1A and B) compared with human PSMA. Amino acid residues predicted to represent the binding pocket and catalytic domain of the PSMA enzyme exhibited 100% identity, as previously reported for some of the orthologs tested. 1 Therefore, a PSMA substrate inhibitor such as DCFPyL would be expected to bind with comparable affinities within the PSMA binding pocket across these species. 33
In vitro autoradiography
In vitro autoradiography saturation studies (Figs. 2 and 3A) with the SMG gland of mice, rats, baboons, cynomolgus monkeys, rhesus monkeys, and humans were performed to determine: (i) PSMA concentration levels (Bmax) and (ii) binding affinity (Kd) of [ 18 F]DCFPyL to PSMA. SMG sections for all species exhibited high PSMA specific binding (Bsp) ranging from 90% to 95% with high [ 18 F]DCFPyL binding affinities (Kd, 0.42–1.39 nM; Fig. 3B). The highest binding affinity (0.42 nM) was observed for human SMG, with slightly lower affinities observed for cynomolgus monkeys (0.55 nM) and rats [(0.76 nM); Fig. 3B]. Lower binding affinities of two to three-fold compared with human SMG were observed in rhesus monkey (1.08 nM), mouse (1.24 nM), and baboon (1.39 nM). The binding affinity of [ 18 F]DCFPyL for human SMG glands (Kd = 0.42 nM) was comparable to published values for human prostate cancer cells (Kd = 0.49 nM). 34 In summary, our studies demonstrated that [ 18 F]DCFPyL exhibits low nanomolar binding affinities for SMG PSMA for all tested species (Fig. 3B).
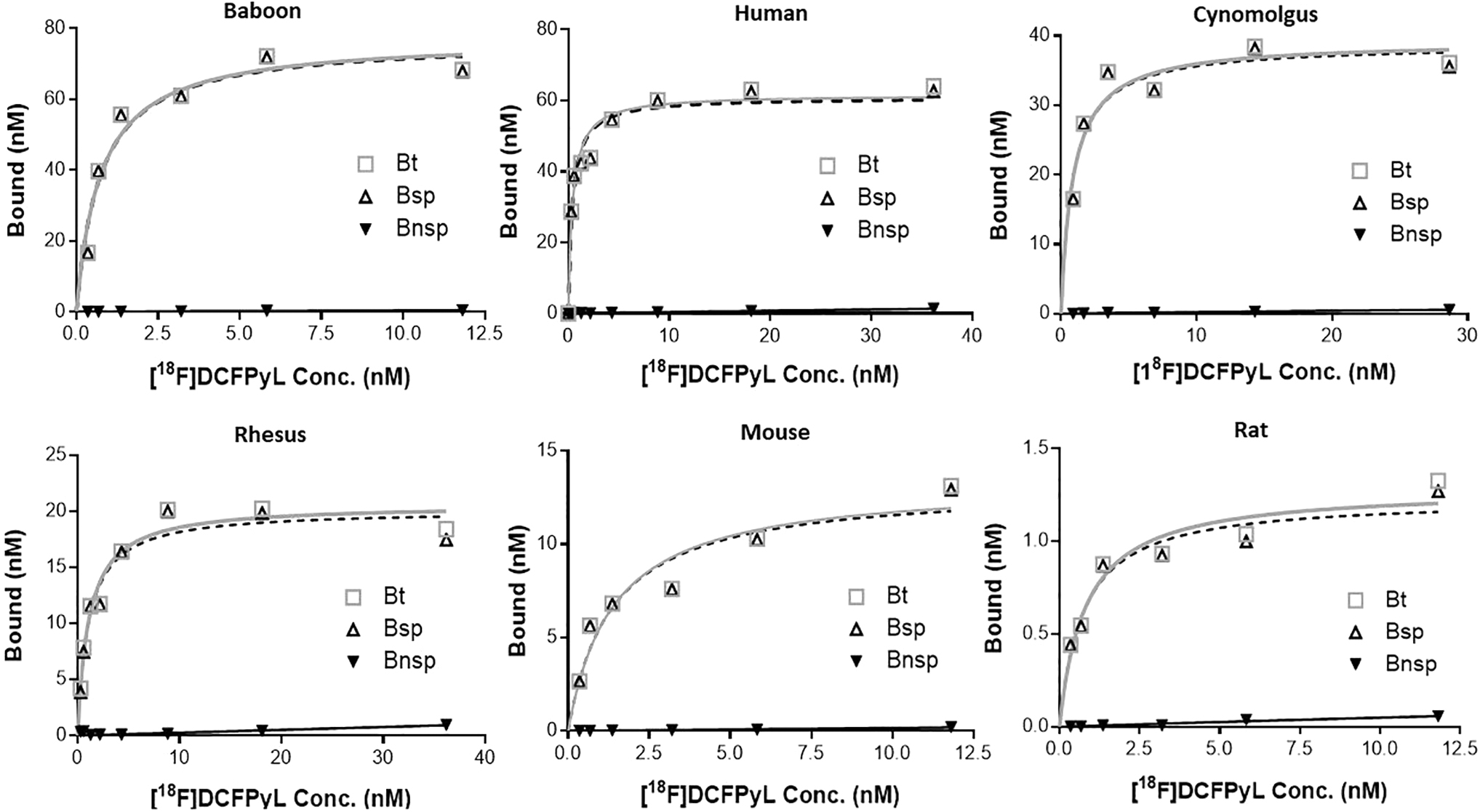
Representative plots of in vitro autoradiography saturation binding studies of human, non-human primates (baboon, cynomolgus, rhesus) and rodent (mouse, rat) submandibular glands; each point (n = 6 points) is derived from ROI analysis. For each plot: Bt = Bound total; Bnsp = Bound non-specific; Bsp = Bound specific (Bt-Bnsp = Bsp). ROI, regions of interest.
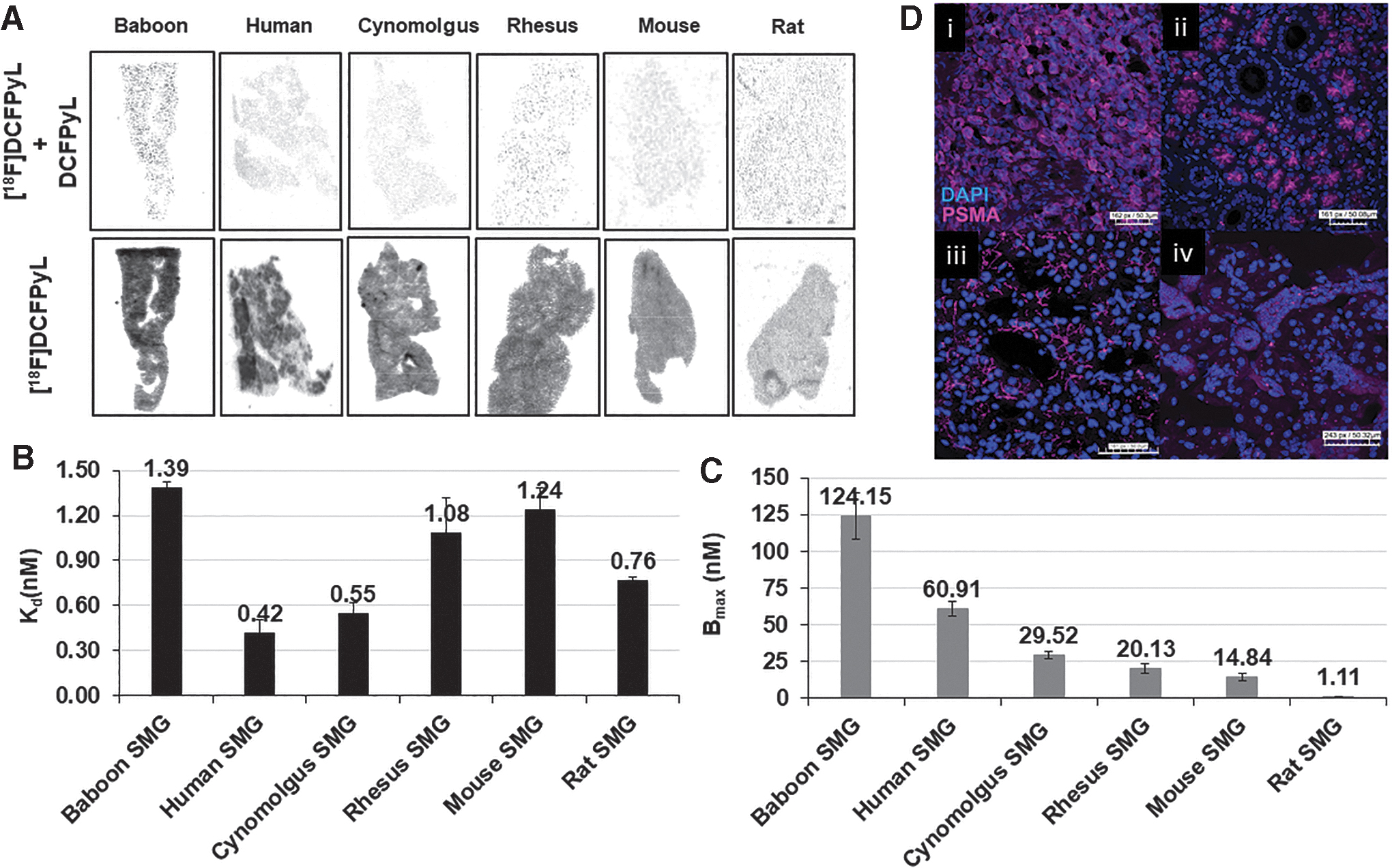
The PSMA concentration (Bmax) in SMG across the species was determined from the saturation assays (Fig. 3C) and ranged from 124.15 nM to 1.11 nM. Baboons expressed the highest level of PSMA concentration with a Bmax value of 124.15 nM and this was ∼2-fold higher than humans (Bmax = 60.91 nM). Cynomolgus and rhesus monkey SMG displayed a Bmax of 29.52 nM and 20.13 nM, ∼2-fold less than that of human SMG. The PSMA level (Bmax) in mice and rat SMG was 14.84 nM and 1.11 nM, respectively. These results indicate that [ 18 F]DCFPyL would be able to detect PSMA in SMG in vivo for all the species tested except for the rat due to low PSMA expression.
Submandibular PSMA expression determined by IF
The protein expression and localization of PSMA in submandibular glands (SMGs) of human, baboon, mouse, and PC3-PSMA tumors were determined by IF (Fig. 3D). PSMA IF demonstrated strong, apical membranous staining exclusively in the acini of SMG in humans and baboons whereas PC3-PSMA tumor cells demonstrated a strong, diffuse cytomembranous pattern of expression. Mouse SMG demonstrated a weak cytomembranous pattern of staining with moderate focal puncta arranged in a perinuclear pattern. These results support the in vitro binding study data demonstrating that humans and baboons exhibit similar protein expression whereas mice have noticeably lower expression. To confirm these results, publicly available RNA sequencing data were queried to determine the human and mouse SMG FOLH1 expression (Supplementary Fig. S2). FOLH1 mRNA is expressed at low levels in the mouse salivary gland, compared with moderate expression of FOLH1 in the human salivary glands. These data agree with in vitro binding data.
In vivo biodistribution in mice and rats
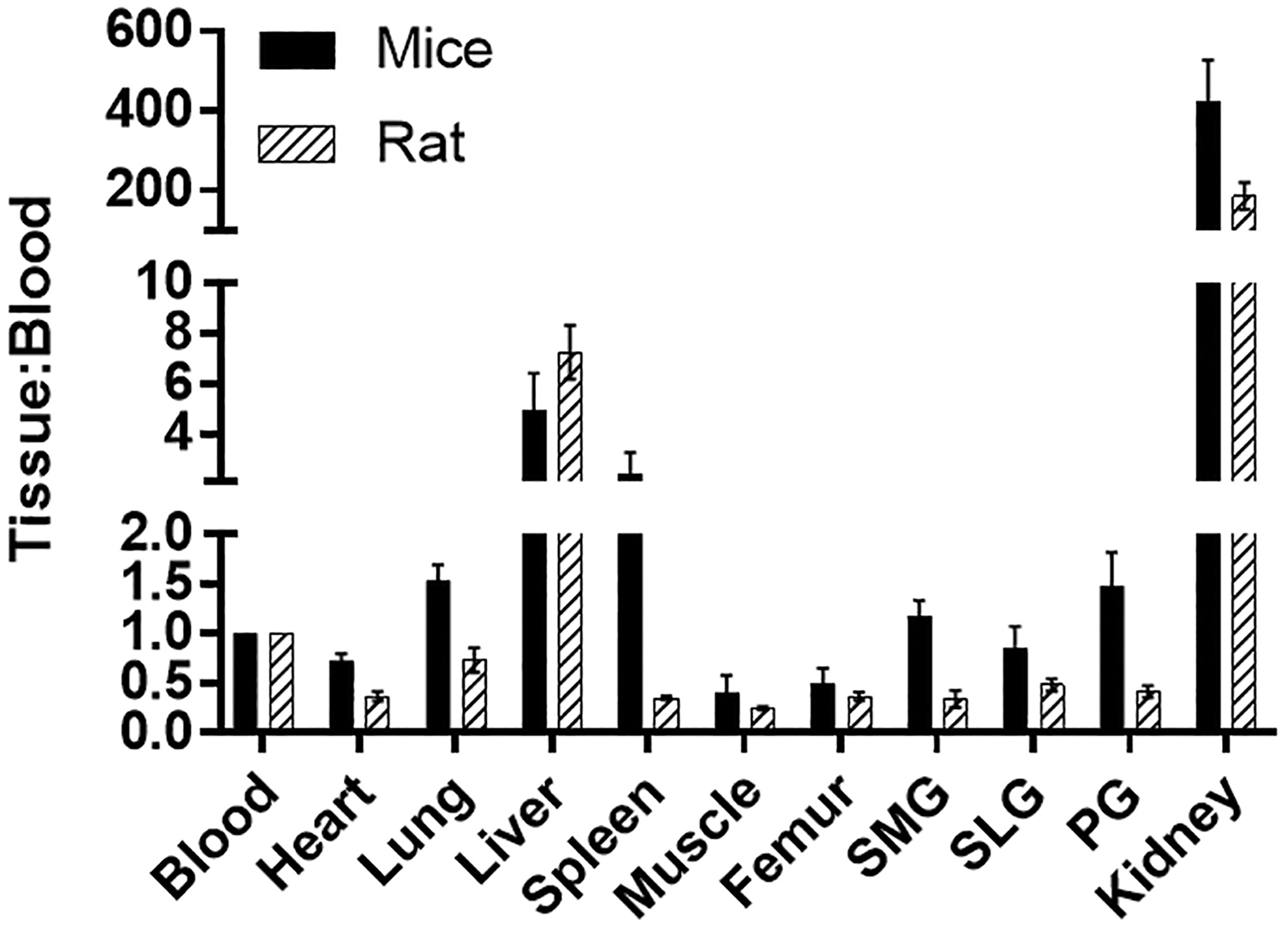
The biodistribution of [ 18 F]DCFPyL was determined in mice and rats at 1 h post-injection (Fig. 4), with the highest uptakes (%ID/g) in the kidney for both mice (132%ID/g) and rats (33%ID/g). The [ 18 F]DCFPyL uptake in mouse submandibular (SMG, 0.37%ID/g), sublingual (SLG, 0.26%ID/g), and parotid (PG, 0.46%ID/g) glands was higher than in rat SMG (0.065%ID/g), SLG(0.054%ID/g), and PG(0.085%ID/g). Further, the uptake in rat salivary glands is similar to the uptake in the muscle (0.04%ID/g), which represents non-target tissue binding. The tissue to blood ratios for all the organs was calculated from the %ID/g of tissue (Fig. 5). The tissue to blood ratio (T:B) of the SMG, SLG, and PG for mice was found to be 1.17, 0.85, and 1.48 whereas the T:B in rat SMG, SLG, and PG was 0.34, 0.48, and 0.41, respectively. All major salivary glands of the mice showed higher T:B than the rats (SMG, 3.4-fold; SLG, 1.7-fold; and PG, 3.6-fold). The rat salivary gland T:Bs were comparable to muscle (0.25), indicating that the uptake observed in salivary glands of rats may not represent PSMA binding but the radioactivity associated with the tissue blood volume. In both mice and rats, the highest uptakes were observed in kidneys with a T:B of 424.5 and 186.4, respectively. Similar to the T:B, the tissue to muscle ratio (T:M) of mouse SMG (3.2), SLG (2.2), and PG (3.9) glands was higher than that of rats (SMG:1.5, SLG:1.3, and PG:1.9; Fig. 6). These higher levels of PSMA observed in mice compared with rats in vivo agree with the in vitro autoradiography results.
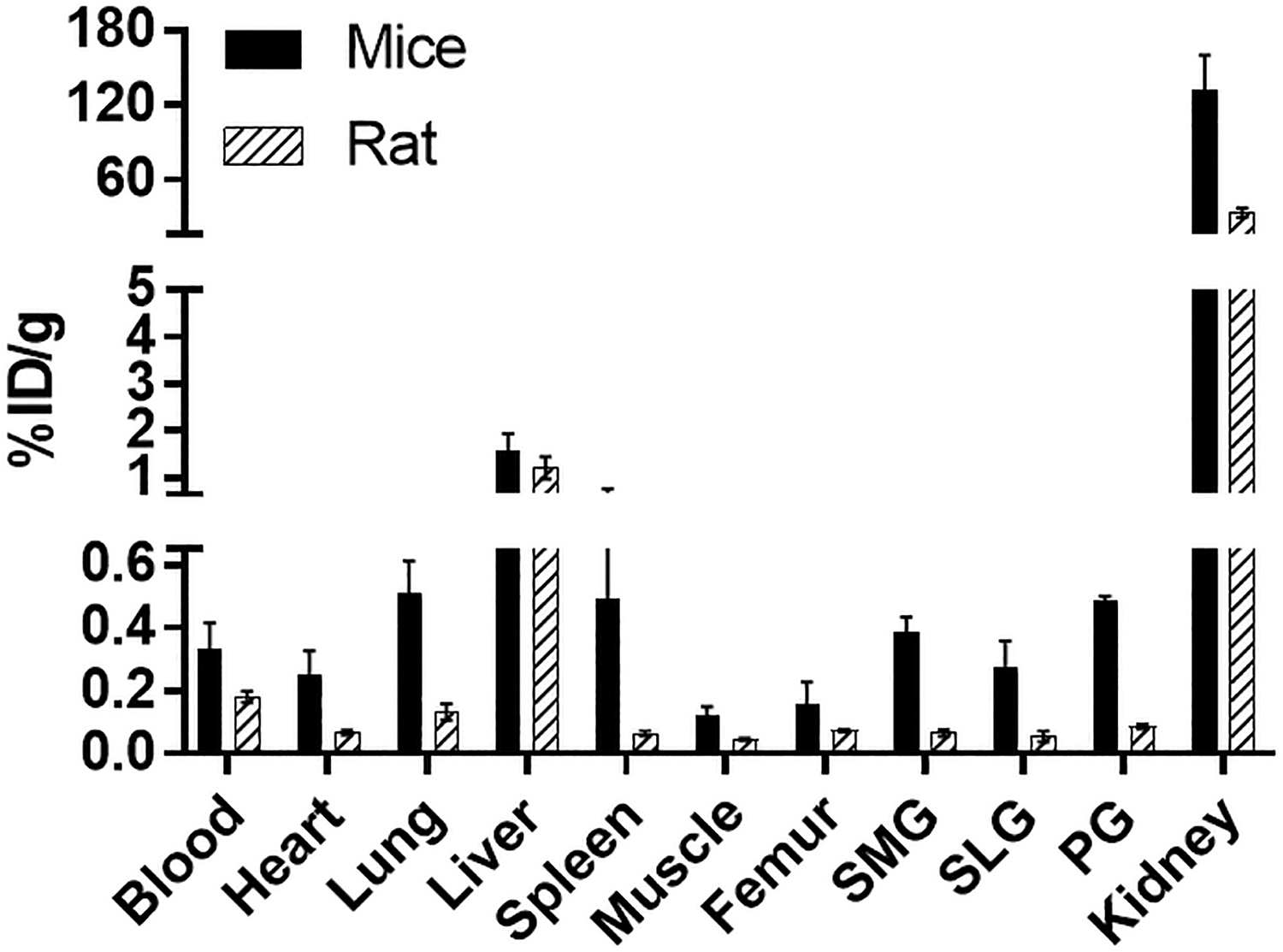
Comparison of [ 18 F]DCFPyL biodistribution in rats and mice at 1 h after injection of [ 18 F]DCFPyL. Each bar represents %ID/g ± SD (for mice: %ID/g normalized to 20 g mice; for rats: %ID/g normalized to 125 g rat; n = 5 for each group). SMG, SLG, and PG represent submandibular, sublingual, and parotid glands, respectively.

Discussion
These studies indicate that the binding affinity of [ 18 F]DCFPyL for PSMA was similar across the animal species compared with humans, although the PSMA expression levels varied. Because DCFPyL and other synthetic PSMA ligands have been designed specifically for human PSMA, it is important to first understand the protein similarities across these animal models. Overall, there is high amino acid conservation for PSMA across the tested species, with high similarity between humans and baboons, less so for cynomolgus and rhesus monkeys, and lowest for mice and rats. However, in the predicted binding domain of PSMA, there is 100% identity across the non-human primate and rodent species compared with humans.
Further, not only should the ideal animal salivary gland model have PSMA binding domains that are similar, but also PSMA expression levels in the salivary glands should approximate those found in humans as well. PSMA expression in the salivary glands is highly specific to the apical membrane of acini but overall, protein expression is low when compared with other acini-specific salivary proteins (e.g., AQP5 or NKCC1,
Despite the potential of internalization to affect PSMA densities, the in vitro autoradiography results compared favorably with the in vivo biodistribution results in which PSMA was found to be greater in mouse salivary glands compared with rats. To determine whether these PSMA concentrations are enough for visualization of the salivary glands in PET imaging studies, it would be important to determine the target to non-target ratios, which can be predicted from the mathematical equation:
where B = Bound concentration, F = Free concentration, Bmax = Maximum receptor concentration, and Kd = Dissociation constant. Using the equation just cited, the target to non-target ratio of 2 to 1 was predicted for mice SMG, indicating that [
18
F]DCFPyL may be useful to image these glands in vivo. On the other hand, the target to non-target ratio for rat SMG was found to be 0.126, which indicates that in vivo PSMA-specific binding would be indistinguishable from the background. Thus, [
18
F]DCFPyL may be useful to monitor PSMA-mediated changes in salivary glands in mice while investigating various methods to preserve salivary gland function post
Conclusion
Our data compare the expression of PSMA in salivary glands of commonly used preclinical animal models. These data should allow researchers to choose the best preclinical animal model for performing salivary gland studies associated with PSMA. Hopefully, this will aid and encourage efforts to develop methods to spare the salivary glands during PSMA-targeted RNT.
Authorship Confirmation Statement
The contributions of each author toward this article are given next.
Jyoti Roy: Study conception and design, acquisition of data, analyzing and interpreting data, drafting of article, and critical revision; Blake M. Warner: Acquisition of data, analyzing and interpreting data, drafting of article, and critical revision; Falguni Basuli: Synthesis of [ 18 F]DCFPyL, critical revision; Xiang Zhang: Synthesis of [ 18 F]DCFPyL, critical revision; Karen Wong: Acquisition of data, critical revision; Thomas Pranzatelli: Acquisition of data, critical revision; Anita T. Ton: Acquisition of data, critical revision; John A. Chiorini: Study conception, critical revision; Peter L. Choyke: Study conception and design, critical revision; Frank I. Lin: Study conception and design, critical revision; Elaine M. Jagoda: Study conception and design, acquisition of data, interpretation of data, and critical revision.
Footnotes
Disclosure Statement
There are no potential conflicts of interest to disclose.
Funding Information
This project has been funded in whole or in part with federal funds from the National Cancer Institute, National Institutes of Health, under Contract No. HHSN261200800001E. The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government. This research was supported [in part] by the Intramural Research Program of the NIH, National Cancer Institute, Center for Cancer Research.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.