
Abstract
Background:
The melanoma differentiation-associated gene-7 (Mda-7)/interleukin-24 (IL-24) is a tumor killing cytokine, the bystander effect of which can be enhanced through tethering to tumor homing peptides (THPs).
Materials and Methods:
After fusing tLyP-1, RGR, and buforin as THPs to Mda-7/IL-24, enzyme-linked immunosorbent assay (ELISA) was used to determine the secretion potency of the recombinant proteins. The killing potency of plasmids expressing IL-24, IL-24.tLyP1, IL-24.RGR, and
Results:
The ELISA result showed that contrary to IL-24.RGR and buf.IL-24,
Conclusion:
IL-24 can tolerate tLyP-1 peptide modification by retaining its secretion potency. Tethering tLyP-1 to IL-24 can induce more apoptosis than its modified versions by RGR or buforin.
Introduction
The melanoma differentiation-associated gene-7 (Mda-7), which was renamed interleukin-24 (IL-24), belongs to the IL-10 cytokine family and encodes a protein with 206 amino acids length.
1
In terms of activity, Mda-7/IL-24 has an immune inflammatory role in physiological conditions.
2
However, when it is overexpressed by a plasmid or a recombinant adenovirus (
A dozen THPs were evaluated to improve the delivery of cytokines and to increase their therapeutic effects in the tumor microenvironment. CRGRRST (RGR) as a pancreatic THP is transported to the target cell nucleus after internalization. A putative receptor for RGR peptide is the platelet-derived growth factor receptor-β, which is highly expressed on the tumor cells surface. 12,13 Johansson et al. showed that combining modified interferon-γ and tumor necrosis factor-α with C-terminal RGR can enhance immunotherapy by both specific entry and through retaining the tumor microenvironment. 14 The tLyP-1 peptide (CGNKRTR) is new among the THPs, which in addition to its internalization by the lymphatic endothelial and tumor cells, can also induce apoptosis. 15,16 tLyP-1 binds to neuropilin-1 (NRP1) and/or neuropilin-2 (NRP2) receptors, which are over-expressed in a wide variety of human tumor cells and as a result could penetrate into them, effectively. 17 Buforin, another anticancer peptide, can selectively target cancerous cells through interaction with ganglioside. 18,19 It crosses over the bilayer lipid, combines with the nucleic acid of the tumor cells, and triggers mitochondrial-dependent apoptosis. 20 Many studies were conducted on the potential antitumor activities of the aforementioned peptides, but none considered the impact in fusion with a cytokine structure such as Mda-7/IL-24.
Therefore, it was hypothesized that tethering RGR, tLyP-1, and buforin to the Mda-7/IL-24 sequence can enhance its antitumor activity and specificity through precise attachment to neighboring malignant cell receptors. In this study, fusions of 3 different peptides, including RGR, tLyP-1, and buforin with Mda-7/IL-24 were evaluated in terms of death induction. The cell killing activity of the modified IL-24 with 3 different peptides was compared in vitro on Huh-7 hepatoma cell line and LX-2 hepatic stellate cells as the normal cell line.
Materials and Methods
Cell lines and culture
The human liver cancer cell (Huh-7), and Ad-293 cell lines were purchased from the national cell Bank, Pastor Institute of Tehran. A Human Hepatic stellate cell (LX-2) was generously provided by Prof. Scott L. Friedman (Mount Sinai School of Medicine, New York, NY). The LX-2 cells are defined as an immortal cell that can preserve the main characteristics of the normal line; hence, it was included as the control cell. All cells were cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% or 5% (for LX-2) fetal bovine serum plus 100 IU/mL penicillin and 100 mg/mL streptomycin (Bioidea Inco., Iran) under 37°C and 5% CO2 atmosphere condition.
Preparation of plasmids expressing IL-24, IL-24.RGR, IL-24-buforin and IL-24.tLyP-1
The plasmids expressing IL-24(pc/IL-24) and
Primer Pairs Used for Amplification of New Constructs
Underlined sequences represent a KpnI and b XhoI recognition sites.
IL, interleukin.
DNA transfection
One day before the transfection, 2 × 105 cells were seeded in a 12-well plate. Transfection was performed with 1 μg plasmid vectors (empty pcDNA3.1 (pc), pc/IL-24, pc/IL-24.RGR, pc/IL-24.tLyP-1, and
Enzyme-linked immunosorbent assay
To assess the presence of new constructs in the supernatants of the transfected cells, the plasmids were transfected into Ad-293 cells and incubated for 72 h. Next, the concentrations of IL-24, IL-24.RGR, IL-24.tLyP-1, and
Cell viability
To evaluate the cell viability, MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide) assay was performed in quadruplicate (Sigma, Germany). A total of 5 × 103 cells were seeded in a 96-well plate and incubated at 37°C in a 5% CO2 for 24 h. After transfection, the cells were incubated for another 36 h. Then, the MTT reagent was added, and the incubation continued for a further 4 h until purple precipitates were visible. Afterward, cells were treated with absolute isopropanol and incubated for 4–5 min at room temperature while gently shaking. Finally, the absorbance of the formazan solution was read spectrophotometrically at 570 nm by a microplate reader (FLUOstar Omega; BMG LABTECH, Germany).
Apoptosis assay
As the most reliable method, the apoptosis assay was carried out by Annexin V staining method (PE-Annexin V Apoptosis Detection kit; BD Biosciences, Inc.). According to manufacturer's instruction, the transfected cells were collected after 36 h of incubation, then washed twice by cold phosphate-buffered saline, and resuspended in binding buffer 1X at a concentration of 106 cells/mL. Next, the cells were stained with phycoerythrin-conjugated Annexin-V and 7-amino-actinomycin (7-AAD) and incubated for 15 min in a dark room. Finally, the binding buffer 1X was added to each tube and analyzed by flow cytometry (BD Biosciences) within 1 h. Data analysis was completed using the FlowJo software. The sum of early and late apoptosis percentages was considered as the total percentage of apoptosis.
cDNA synthesis and quantitative real-time PCR
Isolation of total cellular RNA from cells was performed 24 h after transfection, using Total RNA Isolation System kit (DENAzist Asia, Inc., Iran). The concentration, purity, and integrity of RNA were checked, using gel electrophoresis and spectrophotometry by measuring absorbance at 260 and 280 nm. Almost 1 μg total RNAs was applied to synthesize the cDNA by the Synthesis Premix (Takara Bio, Inc., Japan), according to the manufacturer's instruction.
Quantitative real-time PCR (qPCR) was performed on the QuantStudio Real-Time PCR System, using specific primers for GADD153 and BAX genes (Table 2). Amplification was performed using the temperature cycles: 95°C for 15 min as the first denaturation step, followed by 40 cycles at 95°C, 60°C, and 72°C for 30, 25, and 30 s, respectively. For the control of the amplification specificity, melting curve analysis was performed at 65°C–95°C to separate the specific product from the nonspecific products. The β-actin was selected as a housekeeping gene to normalize the gene expression data. 2−ΔΔCt method was applied to analyze the relative change in gene expression based on the similar efficiency of the qPCRs.
Primer Sequences of GADD-153, BAX, and β-Actin (Reference Gene)
BAX, BCL2-associated X.
The modeling of new modified IL-24-receptor interaction
To evaluate the impact of peptide modifications on the structure of native IL-24 and their interaction with cognate receptors, three-dimensional (3D) models of these constructs were provided. For this purpose, the I-TASSER server was used and the 3D models of these proteins were obtained. 21 IL-20 subfamily used a complex of heterodimer receptor IL-20R1, IL-20R2 (type I), and somehow IL-22R1. The crystal structure of IL-20R1/IL-20R2 complex (4DOH) served as template for docking with IL-24. 22 So, the structures of IL-24 and IL-20R1/IL-20R2 complex (4DOH) were aligned by PyMOL program. 23 Moreover, hydrogen bonds of all modifications were also calculated by SWISS-PDB Viewer program. 24
Statistical analysis
All data were obtained after three independent experiment and the results were analyzed by one-way analysis of variance followed by Tukey post-test evaluation, using Prism version 7 software (GraphPad Software, Inc., San Diego, CA). Values of p < 0.05 were considered statistically significant.
Results
The expression potency of pc/IL-24, pc/IL-24.RGR, pc/IL-24.tLyP-1, and pc/buf.IL-24
The cloning results of two different sequence insertion into the pcDNA3.1 plasmid showed the correct process after evaluating by PCR screening, restriction pattern, and sequencing analysis. The sequencing result demonstrated the correct array of cytokine, linker, and peptide in plasmids, as expected. The efficiency of cell transfection by a known GFP expression control plasmid (pEGFP-C1 vector; Pasteur Institute of Iran) showed a satisfactory transfection, when evaluated in Ad-293, Huh-7, and LX-2 cells. The GFP signal counting under a fluorescent microscope revealed the transfection rates for Ad-293, LX-2, and Huh-7 cells ∼80%, 70%, and 50%, respectively.
To examine the gene expression potency of the new constructs, the concentration of Mda-7/IL-24 in the supernatants of the transfected 293 cells were quantified by enzyme-linked immunosorbent assay (ELISA) (Fig. 1). The results showed that the concentration of Mda-7/IL-24 in the supernatant of the Ad-293 cells transfected by pc/IL-24.tLyP-1, was fairly similar to pc/IL-24 receiving cells (p > 0.05). Despite this, the expression of RGR and buforin-modified proteins showed a significant decrease in comparison with the native form (p = 0.0478 and p = 0.0106, respectively). The concentration of Mda-7/IL-24 in the supernatant of the transfected cells by an empty pcDNA3.1(+) was negligible. The concentration of IL-24 and other recombinant proteins were also measured in parallel in supernatant of LX-2 and Huh-7 cells and the results showed a similar pattern.
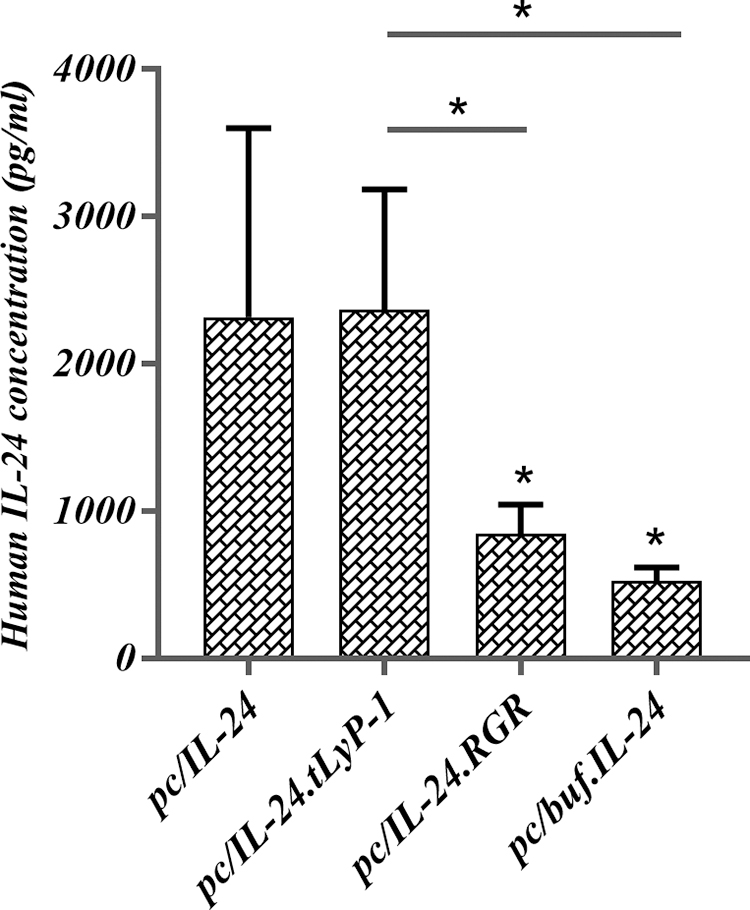
The Mda-7/IL-24 expression level analysis of constructed plasmids by ELISA. The constructs were transfected by Lipofectamine 2000 into Ad-293 cells then supernatant was evaluated after 24 h. The results indicated that the expression level of pc/IL-24.tLyP-1, was fairly similar to pc/IL-24, although the other two constructs exhibited less production (p > 0.05). The expression of pc/IL-24.RGR and
Cell viability analysis
Once the Huh-7 and LX-2 cells (normal cell) were transfected by one of the test plasmids (pc/IL-24, pc/IL-24.tLyP-1, pc/IL-24.RGR, pc/buf.IL-24, and pcDNA3.1), after 36 h after transfection, their viability was investigated, as given in Figure 2. Remarkably, no significant evidence of death-related effects was detected among different plasmid, when evaluated in LX-2 cells. A similar pattern was detected among groups in comparison with the control cells. On the contrary, the result showed a significant viability difference among plasmids, when tested on Huh-7. The MTT assay showed that all Mda-7/IL-24 expressing plasmids exhibited a growth suppressive effect (Fig. 2). However, in Huh-7 cell line, the results showed that among different forms, the antiproliferative activity of pc/Mda-7/IL-24(45.96%) and

The cell viability results of plasmids on Huh-7
Apoptosis induction was significant for tLyP-1-modified protein
The Annexin V/PE staining was used to measure the IL-24-mediated cell death effects by different constructs on Huh-7 and noncancerous LX-2 cell lines. As usual, the Q2 and Q3 quadrants were determined as the early and late stages of apoptotic cells. The flow cytometry results showed that following Huh-7 transfection, the percentage of total apoptosis (Q2+Q3) for the gated population was as follows: 27.63% for the native IL-24, 26.39% for the tLyP-1-modified form, 17.9% for pc/IL-24.RGR, and 16.49% for
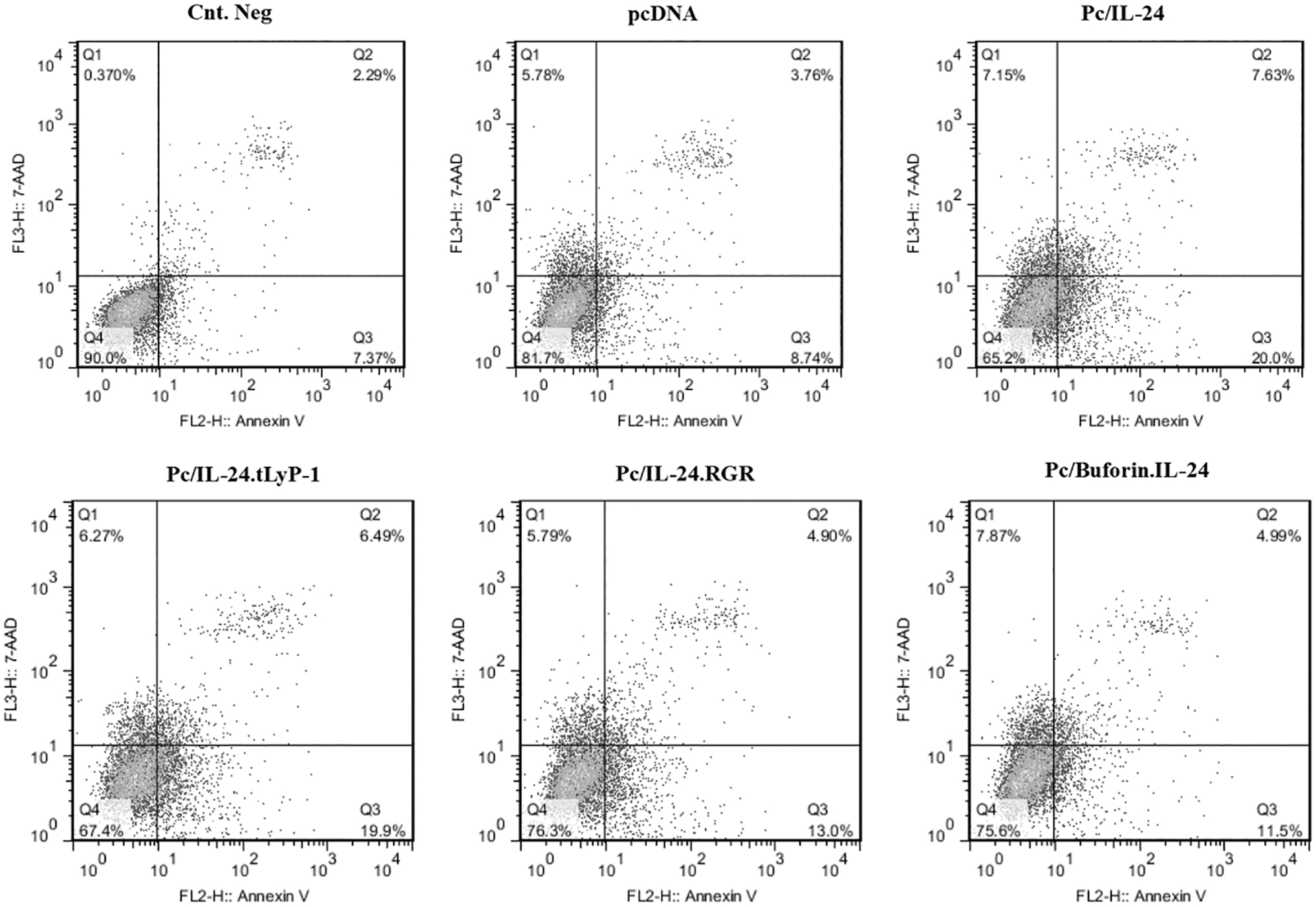
The flow cytometry results of apoptosis induction by recombinant plasmids. The dot plot graphs analysis of apoptosis by Annexin V+ staining in Huh-7 and LX-2 after transfection with pcDNA3.1, pc/IL-24, pc/IL-24, pc/IL-24.tLyP-1, pc/IL-24.RGR, and

The column chart of apoptosis percentage by recombinant plasmids.
Effect of the peptide modified-Mda-7/IL-24 on the expression level of GADD153 and BCL2-associated X genes
To find the killing effect of different recombinant plasmids on Huh-7 cells during the second step, the expression level of two pro-apoptotic genes (GADD-153 and BCL2-associated X [BAX]) were analyzed by qPCR (Fig. 5). The qPCR data analysis showed that the pc/IL-24 and pc/IL-24.tLyP-1, pc/IL-24.RGR, and

The impact of different plasmids on the expression level of GADD-153
Modeling of wild-type and recombinant IL-24 proteins
The 3D molecular models of the wild-type and peptide-modified IL-24 were constructed in I-TASSER web server (Fig. 6). These models showed that these modifications have no special effect on the final structure of IL-24/Mda-7. The 3D structures of IL-20R1 and IL-20R2 subunits and two important sites that involved in binding to the receptors were also revealed as site 1 and site 2. This modeling showed that different modified IL-24 could occupy the similar sites on the receptor without any differences (Fig. 7).

Representation of Mda-7 proteins structure and its modifications.

The superimposition of different modified IL24 proteins on cognate receptor.
Discussion
The ectopic and endogenous expression of Mda-7/IL-24 can induce apoptosis in almost every cancerous context by promoting ER stress and modulating various signal transduction pathways. 4,25,26 Moreover, the secreted form of protein is shown to harbor a potent antitumor bystander effect when docked on its cognate receptors. 27,28
Therefore, a group of modifications, such as tethering to THPs, were developed to concentrate these soluble forms into tumor tissues by enhancing the Mda-7/IL-24 bystander effect. 2 The elegant coupling of soluble form to THPs can lead to specific binding of Mda-7/IL-24 to the relevant peptide receptors, which are highly expressed on the surface of distant untransduced tumor cells. 29,30
In this study, we tethered three different THPs, including RGR, tLyP-1, and buforin to N- and C-terminal of Mda-7 and then evaluated their growth suppressive effects on the hepatocellular carcinoma cell line. The ELISA result showed that secretion of
The killing potency of the newly modified Mda-7/IL-24 produced encouraging result after tethering of tLyP-1, but was compromising for RGR and buforin. The MTT and flow cytometry results revealed that
Considering these findings, it appears that tethering tLyP-1 to the carboxyl end of Mda-7 cannot disrupt the attachment to the receptors, but this is not the case for RGR and buforin peptides.
IL-24.RGR and
Tethering THPs to Mda-7/IL-24 has led to different results, even negative impacts on its natural killing activities. The coupling of RGD-4C sequence (a cyclic form of RGD) to the N-terminal of IL-24 exhibited an enhanced apoptosis induction on MCF-7 and HepG2 cells in comparison with natural Mda-7/IL-24. 31,32 Furthermore, in a recent study, the apoptosis induction potency of Mda-7/IL-24 in the presence of RGD peptide was assessed on activated LX-2 cells. Their results showed that apoptosis was significantly enhanced in comparison with the control group. 33 On the contrary, various studies have shown that attaching some THP to Mda-7/IL-24 can decrease apoptosis induction potency. It was observed that fusing RGD4C sequence to the C-terminal of Mda-7/IL-24 could inhibit natural attachment to the cognate receptor, and adversely reduce killing activity. 34 In agreement with previously published data, the computational analysis revealed that carboxyl modification of Mda-7/IL-24 by THP sequences might disrupt the Mda-7/IL-24 attachment to cognate receptors. 35 In addition, tethering of tRGD and RGD4C peptides to the carboxyl-termini of MDA-7/IL-24 could not improve natural apoptosis property of Mda-7 in vitro. 36
To date, tLyP-1 peptide has been successfully used in conjugation with some biomaterials, 37 but there are limited publications that have described the advantage of tLyP-1 over other peptides in fusion with cytokines for cancer gene therapies. It was anticipated that tLyP-1 could improve the efficacy of Mda-7, as it increases attachment to neighboring cells cognate receptor and also enhances apoptosis of the cells to which it binds. 29 The known tLyP-1 receptors, NRP1 and NRP2 is highly expressed on the majority of tumor cell as well as surrounded vessels. 38 Therefore, a part of cell death that was induced by this cytokine was owing to attachment of peptide-modified IL-24 to tLyP-1 receptor.
Contrary to previous study, the 3D modeling of modified IL-24 and protein–receptor binding showed that these modifications have no significant effects on the final structure of proteins nor on the way they attach to the receptors. 35
Apart from the strength in this study, there were few limitations. The first is the lack of more relevant cancerous and normal cell lines. Another limitation was the absence of protein analysis, especially in signaling pathways and also the lack of protein expression analysis of GADD153/BAX, using Western blotting.
In conclusion, the tethering of peptide, such as tLyP-1 to Mda-7/Il-24 can improve its apoptosis effect on hepatocellular carcinoma cell line as much as the native IL-24 while sparing the normal stellate cell line.
Footnotes
Acknowledgments
The authors thank all staff of Bacteriology and Virology Department for their assistance. The authors also thank H. Argasi at the Research Consultation Center (RCC) of Shiraz University of Medical Sciences for his invaluable assistance in editing this article.
Authors' Contributions
M.R., A.S. M.R., and F.Z. participated in bench works. M.R., S.Y.H., A.B.B., G.R., and S.S. have developed tests at start time. M.R., S.Y.H., and S.S. designed the study. The idea came from a communication by all authors. Data processing and analysis was performed by M.R., A.S., S.S., and G.R. Article writing and processing was fulfilled by M.R., S.Y.H. and A.B.B. and then edited by all the authors.
Disclosure Statement
No competing financial interests exist.
Funding Information
The study was supported by Shiraz University of Medical Sciences, as an MSc thesis in the field of Biotechnology (Grant Number: 1396-01-10-14774).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.