
Abstract
Background:
Melanoma, belonging to a kind of skin cancer, takes a big part in cancer-associated deaths globally. Abundant documents have recorded the crucial roles of long noncoding RNA (lncRNA) in the initiation and development of tumors. lncRNA forkhead box D3 antisense RNA 1 (FOXD3-AS1) has been commonly identified as a key regulator in the progression of multiple cancers; however, the way it exerts function remains obscure in melanoma.
Materials and Methods:
FOXD3-AS1 expression was examined by RT-qPCR. The role of FOXD3-AS1 in melanoma was determined by 5-ethynyl-2′-deoxyuridine (EdU), transwell, and Western blot assays. The combination between microRNA-127-3p and FOXD3-AS1 (or four jointed box 1 [FJX1]) was confirmed by luciferase reporter and RNA immunoprecipitation assays.
Results:
FOXD3-AS1 was markedly upregulated in melanoma cells. It was validated by loss-of-function assays that cell proliferation and migration were inhibited by FOXD3-AS1 deficiency, while cell apoptosis was facilitated by FOXD3-AS1 knockdown in melanoma. Mechanistic exploration testified that miR-127-3p could bind to FOXD3-AS1 and its expression was negatively modulated by FOXD3-AS1 in melanoma. Besides, overexpression of miR-127-3p repressed melanoma progression. Moreover, miR-127-3p was certified to negatively regulate the expression of the FJX1, and miR-127-3p could combine with FJX1 in melanoma cells. Rescue assays depicted that FJX1 overexpression countervailed FOXD3-AS1 silencing-mediated inhibition on melanoma progression.
Conclusions:
Overall, FOXD3-AS1 contributes to the progression of melanoma via miR-127-3p/FJX1 axis.
Introduction
Described as a severe malignancy derived from transformation of melanocytes, melanoma has taken a major part in cancer-associated deaths in the past few years. 1 –3 In recent years, the incidence of melanoma is continuously rising, and the mortality remains at a high level because of the reoccurrence and metastasis. 4 –6 Even though traditional therapies, such as surgery and chemotherapy, could cure early stage melanoma patients, improving the therapy and prognosis of patients at later stage is still an extremely difficult problem that has not been overcome. 7 In recent years, molecular targeted therapy has been applied to cancer therapy, which is a new therapeutic notion making a specific death of tumor cells by designing the corresponding medicines to combine specific cancer sites.
Although some therapeutic targets have been reported in cancers, 8 –10 more unrevealed targets need to be explored. Therefore, to achieve the goal of developing more efficient treatments for melanoma patients, it is urgently required to find proper molecular therapeutic target.
Long noncoding RNAs (lncRNAs) have been classified as a cluster of RNAs with over 200 nucleotides in length, which cannot encode proteins. 11 –13 The critical function of lncRNAs in tumor progression has been observed by abundant studies in recent years. For example, Androgen-induced lncRNA SOCS2-AS1 accelerates tumorigenesis and development in prostate cancer. 14 lncRNA-LOWEG is associated with gastric cancer cell invasion. 15 lncRNA NKILA promotes the anticancer effect of baicalein by regulating nuclear factor-kappaB signaling in hepatocellular carcinoma. 16
In addition, accumulating evidence has also revealed the crucial function that lncRNAs exerted on the tumorigenesis of melanoma. For instance, lncRNA FALEC facilitates melanoma progression by epigenetically silencing p21. 17 lncRNA MHENCR facilitates melanoma development through regulation of miR-425/489-mediated PI3K-Akt pathway. 18 lncRNA-ATB regulates melanoma progression via miR-590-5p/YAP1 axis. 19
Forkhead box D3 antisense RNA 1 (FOXD3-AS1), a kind of lncRNA, has been mentioned to be involved in the course of cancer development according to existing studies. For illustration, lncRNA FOXD3-AS1 deficiency inhibits malignant glioma cell growth and metastasis. 20 LnRNA FOXD3-AS1 modulates the progression of neuroblastoma via regulation of PARP1-mediated activation of CTCF. 21 Despite that FOXD3-AS1 has been confirmed to act as a key regulator in glioma and neuroblastoma, 20,21 its specific role in melanoma still needs to be clarified.
In this study, FOXD3-AS1 was selected to be the object of the authors' research so as to make a thorough understanding of its regulatory role in melanoma progression. All the findings from this study suggest that FOXD3-AS1 contributes to the progression of melanoma via miR-127-3p/four-jointed box 1 (FJX1) axis, offering a novel direction for researches on melanoma treatment.
Materials and Methods
Cell culture
Four human melanoma cell lines (A2058, SK-MEL-28, SK-MEL-2, and A375) and normal human epidermal melanocytes (HEMa-LP) were all obtained from the Institutes for Biological Sciences at the Chinese Academy of Sciences (Shanghai, China), and then maintained continuously in DMEM (Thermo Fisher Scientific, Waltham, MA) with a supplement of 10% fetal bovine serum and 1% antibiotics, both from Thermo Fisher Scientific. All cells were cultured at 37°C with 5% CO2.
Cell transfection
SK-MEL-2 or A375 cells were incubated under the standard conditions and seeded into six-well plates. The short hairpin RNAs (shRNAs) sequences targeting human FOXD3-AS1 (sh-FOXD3-AS1#1/2), miR-127-3p mimics, their corresponding negative control (sh-NC), NC mimics, the FJX1 overexpressing plasmids, and empty pcDNA3.1 vector were constructed by ZonHon Biopharma Institute (Changzhou, Jiangsu, China). The transfection of above plasmids was carried out in SK-MEL-2 or A375 cells by Lipofectamine 2000 (Invitrogen, Carlsbad, CA). After 48 h, the transfected cells were collected for subsequent experiments.
Transwell migration assay
SK-MEL-2 or A375 cells after transfection were resuspended in serum-free medium and then planted into the upper chamber. After being incubated for 1 d, the membrane was removed with a cotton swab on the upper surface. The migrated cells attaching on the lower chamber were immobilized and stained by separately using methanol and crystal violet, both from Beyotime (Shanghai, China). Images were captured by use of an inverted microscope (Olympus, Tokyo, Japan).
RT-qPCR analysis
Total RNAs, which were isolated from SK-MEL-2 or A375 cells via the TRIzol Reagent (Invitrogen), were reverse transcribed into cDNA using the PrimeScript RT reagent Kit (Takara, Osaka, Japan). The RT-qPCR was performed using the TaqMan miRNA Reverse Transcription kit (Thermo Fisher Scientific). The results were normalized to GAPDH or U6 and computed by the 2−ΔΔCt method.
Subcellular fractionation
Extraction of the cytoplasm and nucleus from SK-MEL-2 or A375 cells were conducted using NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific). The RT-qPCR was used to assess FOXD3-AS1 expression. The nucleus control (U6) or cytoplasm control (GAPDH) was separately determined.
Western blotting
Transfected SK-MEL-2 or A375 cells were first collected, and then lysed through RNA immunoprecipitation assay (RIPA) buffer (Pierce, Rockford, IL). Proteins, which were collected after centrifugalizing, were separated by SDS-PAGE (10% polyacrylamide gels) and then transferred to polyvinylidene fluoride membranes. After the above treatment, the membranes were probed with primary antibodies, including anti-Bcl-2 antibody (1/2000, ab182858; Abcam, Cambridge), anti-Bax antibody (1/1000, ab32503; Abcam), anti-FJX1 antibody (1/1000, ab80264; Abcam), and anti-GAPDH antibody (1/10,000, ab245356; Abcam). Then, phosphate-buffered saline (Thermo Fisher Scientific) was applied to elute the membranes for three times, and treated with an appropriate secondary antibody. Relative expression levels of protein were assessed by ImageJ software (NIH Image, Bethesda, MD).
Luciferase reporter analysis
The sequences of FOXD3-AS1-WT, FOXD3-AS1-Mut, FJX1-WT, and FJX1-Mut covering putative miR-127-3p binding sites were obtained and then cloned into the pmirGLO luciferase reporter vectors (Promega, Madison, WI). Later, formed reporter vectors with miR-127-3p mimics or NC mimics were, respectively, cotransfected into SK-MEL-2 or A375 cells using Lipofectamine 2000 (Invitrogen), followed by incubation for 24 h. Luciferase activities were measured via a Dual-Glo Luciferase Reporter Assay Kit (Promega).
5-ethynyl-2′-deoxyuridine
5-ethynyl-2′-deoxyuridine (EdU) proliferation assay was utilized for studying cell proliferation capacity as per the protocol. After transfection, A375 or SK-MEL-2 cells were treated for 2 h by EdU (RiboBio, Nanjing, China). Following Apollo staining and DAPI staining (Thermo Fisher Scientific), EdU-positive cells were visualized via a fluorescence microscope (Leica, Mannheim, Germany).
RNA immunoprecipitation assay
A Thermo Fisher RIP kit (Thermo Fisher Scientific) was adopted for RIPA as the supplier requested. In brief, A375 or SK-MEL-2 cells were lysed in RIP lysis buffer (Solarbio, Beijing, China), and then RNAs magnetic beads (Invitrogen) were conjugated to anti-Ago2 antibody (Millipore, Bedford, MA) or anti-IgG antibody (Millipore) and then assayed by qRT-PCR analysis.
Statistical analyses
Each experiment was repeated thrice in the study. Prism 6.0 (GraphPad Software, La Jolla, CA) was used to conduct statistical analyses and data are presented as mean ± standard deviation. Significance of differences was verified by one-way analysis of variance or Student's t-test. p-value below 0.05 was considered as a significant level.
Results
FOXD3-AS1 is overexpressed and its deficiency restrains melanoma progression
According to GEPIA database, an obviously increased expression of FOXD3-AS1 in skin cutaneous melanoma tissues was noticed (Fig. 1A). As the authors know, skin cutaneous melanoma is a subtype of melanoma; therefore, they speculated that FOXD3-AS1 might involve in the development of melanoma. Then they detected FOXD3-AS1 expression in melanoma cell lines (A2058, SK-MEL-28, SK-MEL-2, and A375) and human epidermal melanocytes (HEMa-LP) was seen as a control. The results of RT-qPCR analysis depicted that FOXD3-AS1 expression was markedly higher in melanoma cell lines, especially in SK-MEL-2 and A375 cell lines, compared with HEMa-LP cell line (Fig. 1B). According to this result, the authors explored the biological effect of FOXD3-AS1 on melanoma cell growth.
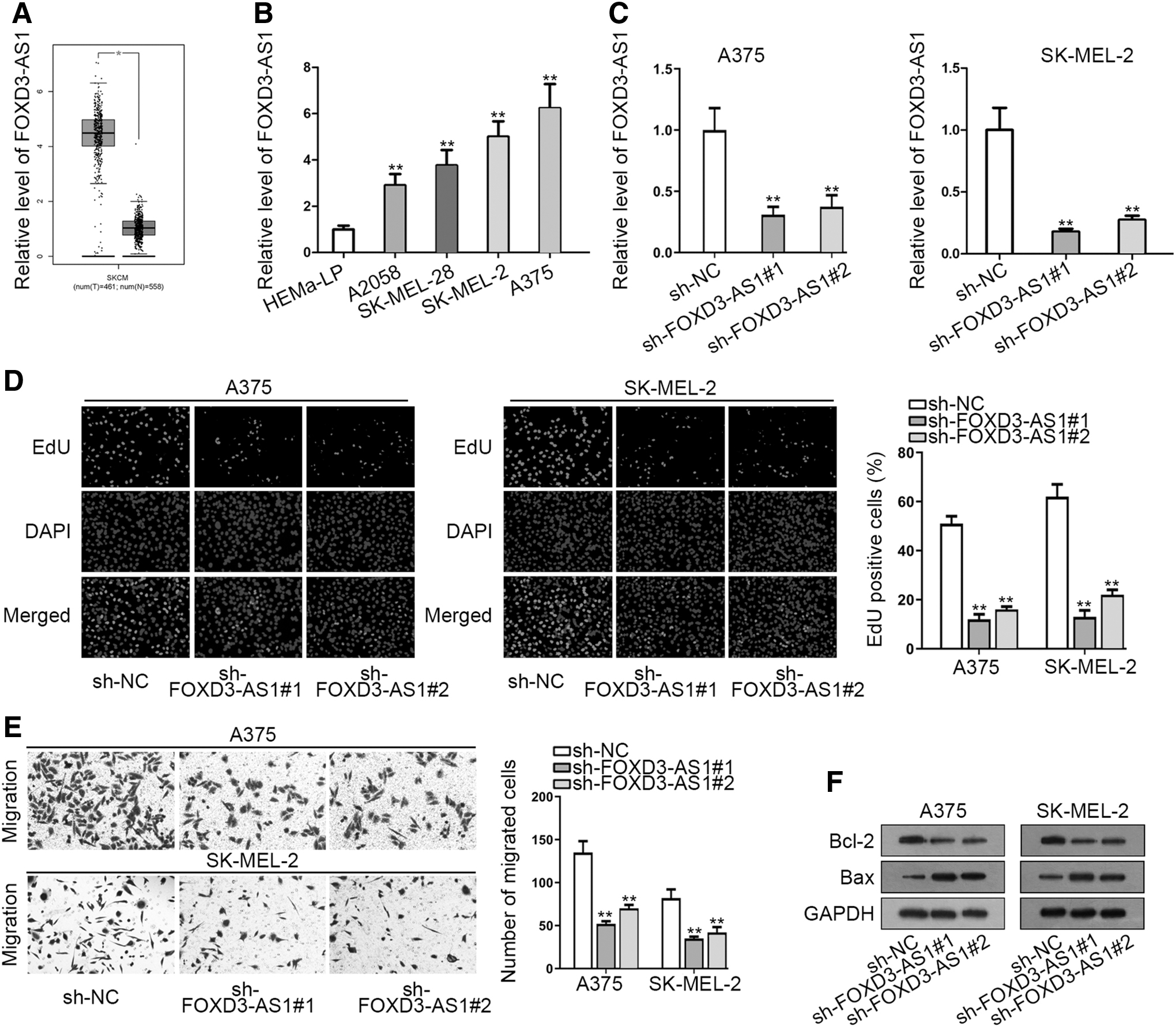
FOXD3-AS1 is overexpressed and its deficiency restrains melanoma progression.
Specific FOXD3-AS1 shRNAs (sh-FOXD3-AS1#1/2) were utilized to stably silence FOXD3-AS1 expression in SK-MEL-2 and A375 cells. RT-qPCR demonstrated that FOXD3-AS1 expression evidently declined with the transfection of sh-FOXD3-AS1 (Fig. 1C). Later, EdU assay uncovered that the proliferative ability was markedly weakened by FOXD3-AS1 deficiency in A375 and SK-MEL-2 cells (Fig. 1D). Besides, silenced FOXD3-AS1 significantly inhibited the migration of A375 and SK-MEL-2 cells (Fig. 1E). Through Western blot assay, a notably low Bcl-2 expression and a conspicuously high Bax expression were observed in sh-FOXD3-AS1-transfected cells, revealing that knockdown of FOXD3-AS1 facilitated melanoma cell apoptosis (Fig. 1F).
In brief, FOXD3-AS1 is highly expressed and its depletion impairs cell proliferation and migration whereas accelerates cell apoptosis in melanoma.
FOXD3-AS1 sponges miR-127-3p and overexpression of miR-127-3p suppresses the progression of melanoma
To make further exploration of FOXD3-AS1 relevant mechanisms in melanoma, subcellular fractionation assay was carried out. The authors first detected the distribution of FOXD3-AS1 in melanoma cells. As a result, FOXD3-AS1 was discovered to take a larger proportion in the cytoplasm of A375 and SK-MEL-2 cells (Fig. 2A), indicating the posttranscriptional regulation of FOXD3-AS1 in melanoma.
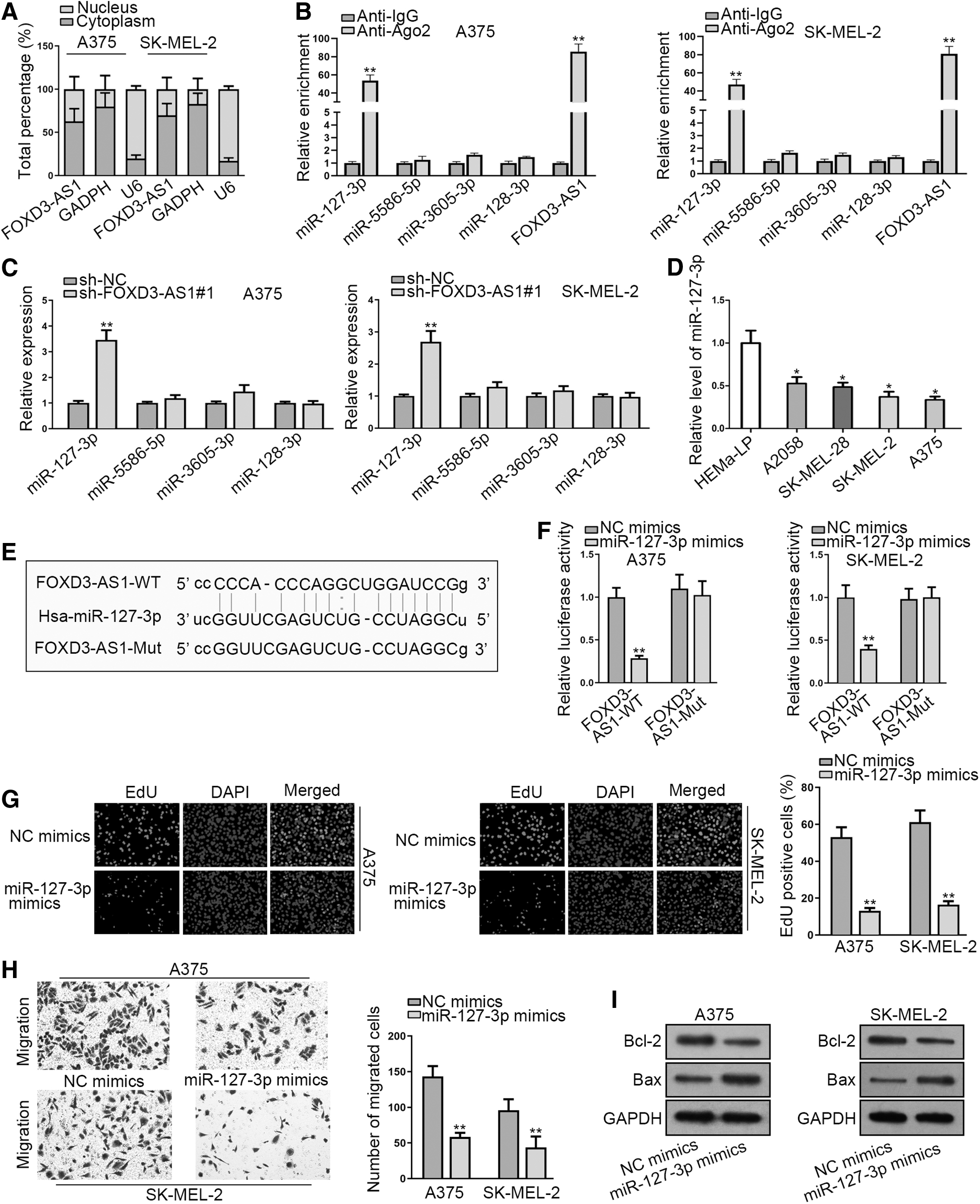
FOXD3-AS1 sponges miR-127-3p and overexpression of miR-127-3p suppresses the progression of melanoma.
Increasing evidence suggested that lncRNA could act as a competing endogenous RNA (ceRNA) to affect tumor progression and regulate downstream mRNAs by sponging microRNAs (miRNAs) at posttranscriptional level. 22,23 Based on this hypothesis, they searched starBase, an online tool to predict miRNA-lncRNA interaction, and four potential miRNAs (miR-127-3p, miR-5586-5p, miR-3605-3p, and miR-128-3p) were found to have the binding capacity with FOXD3-AS1 (Table 1).
The Potential miRNAS Bound to FOXD3-AS1 were Predicted by Starbase
FOXD3-AS1, forkhead box D3 antisense RNA 1; miRNA, microRNA.
To further narrow their selection, RIPA was performed and the results demonstrated that miR-127-3p and FOXD3-AS1 were markedly enriched in anti-Ago2 group in comparison with NC (Fig. 2B). RT-qPCR analysis showed that miR-127-3p expression was observably upregulated by FOXD3-AS1 knockdown (Fig. 2C). Furthermore, an obvious downregulation of miR-127-3p was found in melanoma cell lines by RT-qPCR (Fig. 2D). In addition, the putative binding site between FOXD3-AS1 and miR-127-3p was predicted (Fig. 2E). In luciferase reporter assay, miR-127-3p overexpression led to a remarkable reduction on the luciferase activity of pmirGLO-FOXD3-AS1-WT, while the luciferase activity of pmirGLO-FOXD3-AS1-Mut exhibited no perceivable change (Fig. 2F).
Subsequently, the biological function of miR-127-3p in melanoma was investigated via EdU, transwell, and Western blot assays. First, EdU assay displayed that upregulation of miR-127-3p suppressed the proliferation of A375 and SK-MEL-2 cells (Fig. 2G). Moreover, cell migration capability was decreased by miR-127-3p mimics in A375 and SK-MEL-2 cells (Fig. 2H). As illustrated in Figure 2I, miR-127-3p upregulation significantly reduced Bcl-2 expression, but dramatically enhanced Bax expression, indicating that upregulation of miR-127-3p facilitated the apoptosis of A375 and SK-MEL-2 cells. Taken together, FOXD3-AS1 sponges miR-127-3p in melanoma, and overexpression of miR-127-3p suppresses the progression of melanoma.
FJX1 is a target gene of miR-127-3p in melanoma
To further support ceRNA hypothesis, miR-127-3p targets were probed. Combining the searching results from miRmap, microT and PicTar databases, three candidate target genes (FJX1, KMT5A, and KIF3B) were obtained (Fig. 3A).
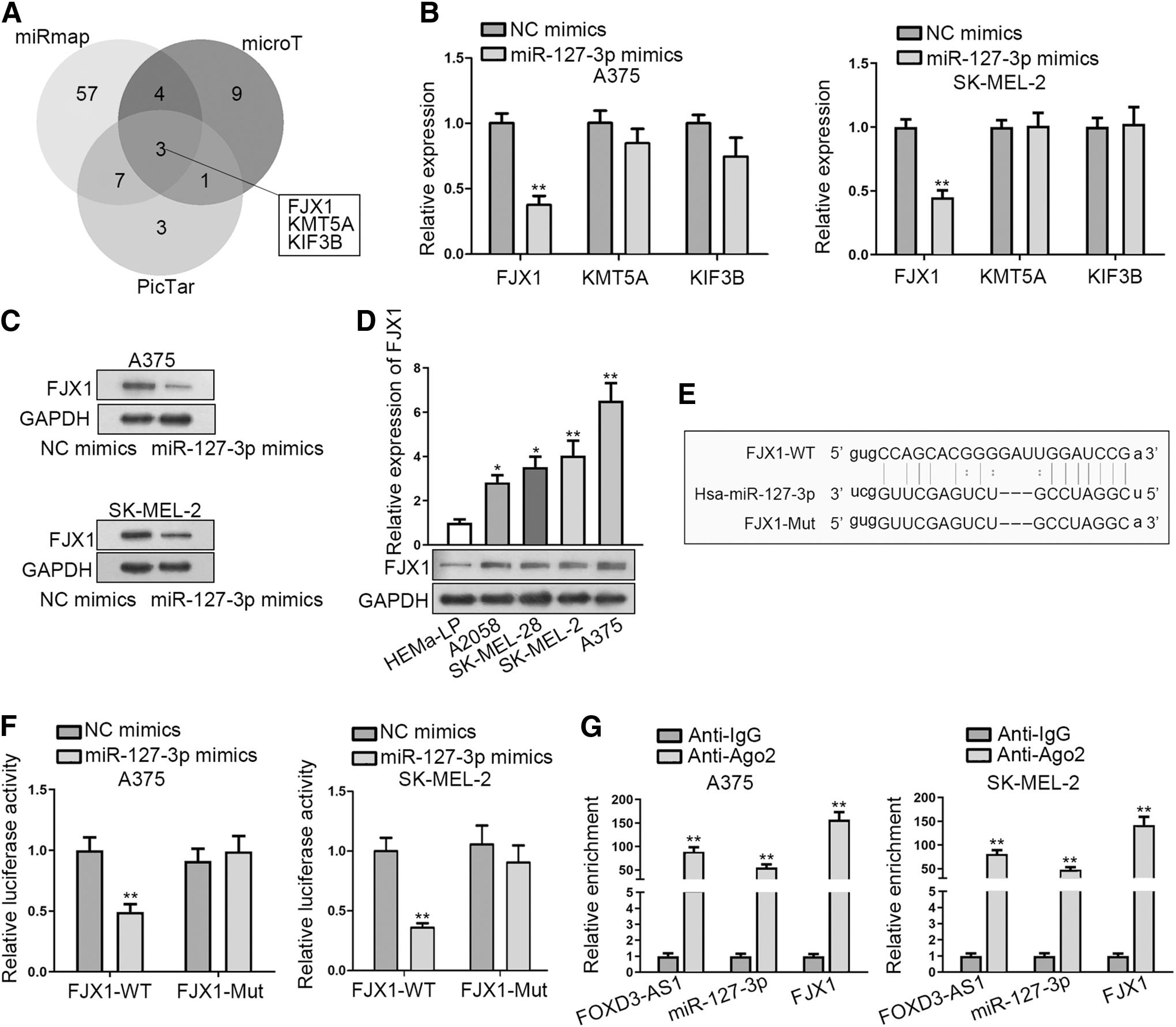
FJX1 is a target gene of miR-127-3p in melanoma.
Then, RT-qPCR assay was applied to evaluate the impact of miR-127-3p mimics on the expressions of these genes. The result demonstrated an evident downregulation of FJX1 in miR-127-3p upregulated A375 and SK-MEL-2 cells, while KMT5A and KIF3B expression presented no differences (Fig. 3B). Thus, FJX1 was selected for the follow-up investigations.
Furthermore, they observed by Western blot assay that miR-127-3p overexpression also observably alleviated FJX1 protein expression in A375 and SK-MEL-2 cells (Fig. 3C). Importantly, a high expression of FJX1 was found in melanoma cells (Fig. 3D).
Through starBase, miR-127-3p was discovered to have a binding site on FJX1 (Fig. 3E). The result of luciferase reporter assay verified that miR-127-3p mimics could weaken the luciferase activity of pmirGLO-FJX1-WT, while it did not impact that of pmirGLO-FJX1-Mut (Fig. 3F). In RIPA, FOXD3-AS1, miR-127-3p, and FJX1 were enriched in the compound precipitated by Ago2 antibody, which validated that miR-127-3p and FJX1 were involved in FOXD3-AS1-mediated ceRNA network (Fig. 3G). Collectively, FJX1 is a target gene of miR-127-3p in melanoma.
FOXD3-AS1 promotes melanoma progression via miR-127-3p/FJX1 axis
To prove whether FOXD3-AS1 promotes melanoma progression via miR-127-3p/FJX1 axis, restoration assays were conducted. Ahead of rescue assays, RT-qPCR was used to examine the transfection efficiency of pcDNA3.1/FJX1 in A375 and SK-MEL-2 cells. The result demonstrated that the expression of FJX1 was significantly increased by transfecting with pcDNA3.1/FJX1 (Fig. 4A).

FOXD3-AS1 promotes melanoma progression via miR-127-3p/FJX1 axis.
According to EdU assay, cell proliferation capability weakened by FOXD3-AS1 knockdown was restored by FJX1 overexpression (Fig. 4B). Moreover, FJX1 overexpression recovered the inhibitive effect of FOXD3-AS1 deficiency on cell migration (Fig. 4C). Western blot assay delineated that upregulation of FJX1 countervailed the FOXD3-AS1 depletion-mediated function on the expressions of apoptosis-associated proteins (Bcl-2 and Bax), which confirmed that FJX1 upregulation could reverse the promoting effect of FOXD3-AS1 silence on cell apoptosis (Fig. 4D). All these data unveiled that FOXD3-AS1 promotes melanoma progression via miR-127-3p/FJX1 axis.
Discussion
Melanoma, a malignant tumor originating from transformation of melanocytes, has induced many cancer-associated deaths in recent years. 1 –3 Abundant investigations have confirmed the regulatory function of lncRNAs in the development of diverse cancer, including prostate cancer, 14 gastric cancer, 15 hepatocellular carcinoma, 16 and melanoma. 17 Although the critical role of FOXD3-AS1 in the complicated progression of glioma 20 and neuroblastoma 21 has been uncovered, the underlying role of FOXD3-AS1 in melanoma remains to be unveiled. In this study, FOXD3-AS1 is highly expressed in melanoma. Besides, FOXD3-AS1 depletion impairs cell proliferation and migration, whereas accelerates cell apoptosis in melanoma. In sum, FOXD3-AS1 is identified as an oncogene in melanoma.
miRNAs are small RNAs with 22–24 nucleotides in length and without protein-coding ability. 24 Accumulating data from previous studies has suggested that lncRNAs play a big part in tumor progression by sponging specific miRNA. For instance, lncRNA UCA1 promotes glioma cell growth and metastasis by targeting miR-122. 25 lncRNA DGCR5 suppresses papillary thyroid carcinoma progression by regulating miR-2861 expression. 26 lncRNA PVT1 accelerates the proliferation and invasion of glioma cells through regulation of miR-200a expression. 27
In this study, starBase was utilized to predict the proper miRNAs that might bind with FOXD3-AS1. Then miR-127-3p was selected due to its remarkable interaction with FOXD3-AS1. As a miRNA, miR-127-3p was widely reported as a tumor suppressor in cancers. For example, miR-127-3p inhibits cell proliferation of oral squamous cell carcinoma by targeting KIF3B. 28 miR-127-3p is downregulated in osteosarcoma tissues and cells and inhibits cell growth via targeting ITGA6. 29 In ovarian cancer, miR-127-3p suppresses cell proliferation and invasion through targeting BAG5. 30 Current study testified that FOXD3-AS1 sponged miR-127-3p and overexpression of miR-127-3p could suppress the progression of melanoma.
FJX1, a member of mRNAs, has been certified to be closely associated with the progression of different diseases, particularly in cancers. For example, FJX1 is involved in nasopharyngeal carcinoma development. 31 FJX1 plays vital roles in endometriosis. 32 FJX1 promotes the angiogenesis of colorectal carcinoma. 33 In this study, the expression of FJX1 was negatively modulated by miR-127-3p in melanoma. Also, FJX1 was found to be highly expressed in melanoma cells. Furthermore, FOXD3-AS1, miR-127-3p, and FJX1 could bind to RISC. Rescue experiments depicted that upregulation of FJX1 countervailed the FOXD3-AS1 downregulation-mediated function on the progression of melanoma, indicating the involvement of FJX1 in the regulation of FOX3D-AS1 on melanoma progression.
To sum up, FOXD3-AS1 contributes to the progression of melanoma by targeting miR-127-3p/FJX1 axis, which shed new lights on researches concerning exploration of melanoma therapies.
Footnotes
Acknowledgment
The authors appreciate all the people involved in this study.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.