
Abstract
Background:
Prostate cancer (PCa) is the second most common cancer among men, threatening men's health and life. Long noncoding RNA Zinc-finger E-box binding homeobox 1 antisense gene 1 (ZEB1-AS1) and Cullin 4B (CUL4B) were reported to be connected with the tumorigenesis of PCa. However, it is unclear whether ZEB1-AS1 regulates the expression of CUL4B in PCa.
Materials and Methods:
The levels of ZEB1-AS1 and CUL4B in PCa tissues and cells were evaluated by quantitative real-time polymerase chain reaction. Protein levels of CUL4B, p21, CyclinD1, matrix metalloprotease 9 (MMP9), E-cadherin, phosphorylated-phosphatidylinositol 3 kinase (p-PI3K), PI3K phosphorylated protein kinase B (p-AKT), AKT, p-mTOR and mammalian target of rapamycin (mTOR) in PCa tissues or cells were assessed by Western blot analysis. The proliferation, migration, and invasion abilities of PCa cells were determined with 3-(4, 5-dimethylthiazol-2-YI)-2,5-diphenyltetrazolium bromide (MTT) or transwell assay. The interaction between ZEB1-AS1 or CUL4B and microRNA-342-3p (miR-342-3p) was predicted using starBase v2.0 database and confirmed by the dual-luciferase reporter assay.
Results:
ZEB1-AS1 and CUL4B were upregulated and miR-342-3p was downregulated in PCa tissues and cells. Both ZEB1-AS1 and CUL4B inhibition constrained proliferation, migration, and invasion of PCa cells. Moreover, the elevation of CUL4B reversed the effects of ZEB1-AS1 silencing on the proliferation, migration, and invasion of PCa cells. Importantly, ZEB1-AS1 modulated CUL4B expression by sponging miR-342-3p in PCa cells. Besides, ZEB1-AS1 mediated PI3K/AKT/mTOR signal pathway by miR-342-3p/CUL4B axis in PCa cells.
Conclusion:
ZEB1-AS1 modulated PCa progression through mediating PI3K/AKT/mTOR signaling by miR-342-3p/CUL4B axis, providing a possible strategy for the treatment of PCa.
Introduction
It was estimated that there were 1,276,106 cases of prostate cancer (PCa) and 359,000 related deaths worldwide in 2018, ranking the second most common cancer among men. 1 At present, prostate-specific antigen (PSA) examination of PCa is controversial. There is evidence that PSA examination can reduce the mortality of PCa, but the low specificity causes unnecessary prostate biopsy and overdiagnosis, resulting in more men suffering from severe side-effects. 2 –4
Although surgical methods and adjuvant chemotherapy have made significant progress, the incidence of PCa worldwide is equably enhancing. 5 However, PCa has a higher rate of recurrence or metastasis, resulting in reduced treatment outcomes and survival rates. 6 In consequence, it is of great significance to explore the pathogenesis of PCa for the development of specific biomarkers and treatment.
Long noncoding RNAs (lncRNAs) are a large class of outstanding nonprotein encoding RNAs, which exert a vital regulatory role in gene regulatory networks. 7 PCa meets with overdiagnosis and overtreatment due to the lack of specific biomarkers, and lncRNA is regarded as a potential biomarker and therapeutic target for PCa. 8 LncRNA E-box binding homeobox 1 antisense gene 1 (ZEB1-AS1), a carcinogenic regulator of a range of malignancies, had been shown to play a vital role in tumorigenesis and development, such as hepatocellular cancer, colorectal cancer, and glioma cancer. 9 –12 Furthermore, ZEB1-AS1 was revealed as an oncogene in PCa. 13 However, the role of ZEB1-AS1 in PCa and its related mechanisms have rarely been reported.
MicroRNAs (miRNAs) are a lumping family of post-transcriptional regulatory genes that exert crucial roles in organism development, viral infection, and tumorigenesis. 14,15 It had been reported that miRNA exerts important roles as a regulatory factor in the biological processes of progression of a variety of cancers. 16 –18 In addition, miRNA-342-3p (miR-342-3p) was revealed to be significantly reduced in triple-negative breast cancer, 19 gallbladder cancer, 20 nasopharyngeal cancer, 21 and nonsmall lung cancer. 22 However, the role of miR-342-3p and its mechanisms is rarely reported in PCa.
Cullin 4B (CUL4B) is a scaffold for the Cullin 4B-Ring E3 Ligase complex, which exerts vital roles in proteolysis. 23 It was reported that the mutation of CUL4B was related to brain malformation. 24 Moreover, CUL4B was connected with adjuvant-induced arthritis pathogenesis. 25 In addition, CUL4B was reported to be concerned with the occurrence and progression of diverse cancers, including PCa. 26 –29 At present, the role of CUL4B and its related mechanisms are still unclear in PCa.
From the above, this study evaluated the expression patterns of ZEB1-AS1 and CUL4B in PCa tissues and cells. Furthermore, the authors explored the role of ZEB1-AS1 and CUL4B in PCa cells. In addition, the regulation mechanism of the ZEB1-AS1/miR-342-3p/CUL4B axis in PCa cells was studied. This study provided a theoretical basis for the development of PCa treatment and specific biomarkers.
Materials and Methods
PCa specimen collection
From Lanzhou University Second Hospital, 30 PCa specimens and paired normal PCa tissues were collected for PCa research. This study was authorized by the Ethical Committee of Lanzhou University Second Hospital. None of the participants in the study had received radiation or chemotherapy, and informed consent was obtained before surgery.
Cell culture
The normal human prostate epithelial cells (RWPE-1) and PCa cell lines (DU145 and LNCaP) were obtained from American Tissue Culture Collection (Manassas, VA). The Roswell Park Memorial Institute (RPMI) 1640 medium (GIBCO, Uxbridge, United Kingdom) supplemented with fetal bovine serum (10%, FBS; Atlanta Biologicals, Atlanta, GA), streptomycin (1%; Baomanbio, Shanghai, China), and penicillin (1%; Baomanbio) was applied to culture all of the above cell lines. Moreover, all cells were cultivated and humidified with 5% CO2 at 37°C in an incubator.
Cell transfection
Small interfering RNA targeting ZEB1-AS1 (si-ZEB1-AS1), small interfering RNA targeting CUL4B (si-CUL4B), and negative control (si-con) were obtained from Genepharma (Shanghai, China). The pcDNA3.1 vector from Invitrogen (Carlsbad, CA) was used for the establishment of the pcDNA-ZEB1-AS1 overexpression vector (pcDNA-ZEB1-AS1) and pcDNA-CUL4B overexpression vector (pcDNA-CUL4B). MiRNA mimic targeting miR-342-3p (miR-342-3p, M12328) and scrambled mimic control (miR-con) were obtained from Ambion, Inc. (Austin, TX). Plasmids or oligonucleotides were transiently transfected into DU145 and LNCaP cells through Lipofectamine 2000 reagent (Invitrogen). The sequences were displayed as follows: si-ZEB1-AS1 (5′-UCAAUGAGAUUGAACUUCAGCUGGA-3′), si-CUL4B (5′-CCACCCAGAAGUCAUUAAUTT-3′) and si-con (5′-UUCUCCGAACGUGUCACGUTT-3′).
Quantitative real-time polymerase chain reaction
To assess the levels of ZEB1-AS1, miR-342-3p, and CUL4B, TRIzol Reagent (Invitrogen) was employed to acquire total RNA from PCa specimens and paired normal PCa tissues, as well as PCa cells. The PrimeScript RT reagent kit (Takara, Dalian, China) or One Step PrimeScript miRNA cDNA Synthesis Kit (Takara) was applied to synthesize the first-strand complementary DNA. The levels of ZEB1-AS1, miR-342-3p, and CUL4B were evaluated by the SYBR Premix Ex Taq™ II kit from Takara.
Primers were exhibited as follows: glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (F: 5′-GAAGGTGAAGGTCGGAGTC-3′, R: 5′-GAAGATGGTGATGGGATTTC-3′); U6 small nuclear RNA (snRNA) (F: 5′-GCTCGCTTCGGCAGCACA-3′, R: 5′-GAGGTATTCGCACCAGAGGA-3′); ZEB1-AS1 (F: 5′-TCCCTGCTAAGCTTCCTTCAGTGT-3′, R: 5′-GACAGTGATCACTTTCATACTT-3′); miR-342-3p (F: 5′-TCCTCGCTCTCACACAGAAATC-3′, R: 5′-TATGGTTGTTCACGACTCCTTCAC-3′), and CUL4B (F: 5′-CCTGGAGTTTGTAGGGTTTGAT-3′, R: 5′-GAGACGGTGGTAGAAGATTTGG-3′). The levels of ZEB1-AS1, miR-342-3p and CUL4B were figured using 2−ΔΔCt method, and U6 snRNA or GAPDH was served as an internal control.
Western blot analysis
The whole protein of PCa specimens and paired normal PCa tissues, as well as PCa cells was extracted with RIPA lysis buffer (Beyotime, Shanghai, China). Then, the BCA Assay Kit from Pierce (Rockford, IL) was applied to quantify the whole protein. After that, the whole protein was separated by 8%–12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE). Subsequently, the separated proteins were transferred to polyvinylidene difluoride (PVDF) membranes (Millipore, Billerica, MA). The PVDF membranes were blocked by Tris-buffered saline tween (TBST) buffer with 5% skim milk for 2 h. Following this, TBST was utilized to wash the PVDF membranes, and the membranes were incubated with primary antibodies overnight at 4°C. Then, the membranes were washed and incubated with goat antimouse or rabbit IgG (Abcam, Cambridge, MA).
Primary antibodies in this study were purchased from Abcam, including mouse-anti-CUL4B (1:500, ab67035), mouse-anti-GAPDH (1:2000, ab8245), rabbit-anti-p21 (1:500, ab227443), rabbit-anti-CyclinD1 (1:500, ab226977), rabbit-anti-matrix metalloprotease 9 (MMP9) (1:1000, ab38898), rabbit-anti-E-cadherin (1;1000, ab133597), mouse-anti-phosphatidylinositol 3 kinase (PI3K) (1:1000, ab86714), rabbit-anti-phosphorylated-phosphatidylinositol 3 kinase (p-PI3K) (1:1000, ab182651), rabbit-anti-AKT (1:2000, ab185633), rabbit-anti-p-AKT (1:1000, ab133458), rabbit-anti-mTOR (1:2000, ab2732), and rabbit-anti-p-mTOR (1:1000, ab109268). GAPDH was regarded as a loading control. The EasySee Western Blot kit from Transgen Biotech (Beijing, China) was employed for the detection of protein bands. The Quantity One software from Bio-Rad Laboratories (Hercules, CA) was employed for the assessment of protein bands.
3-(4,5-Dimethylthiazol-2-YI)-2,5-diphenyltetrazolium bromide assay
3-(4,5-Dimethylthiazol-2-YI)-2,5-diphenyltetrazolium bromide (MTT) assay was carried out for the assessment of cell proliferation. In short, transfected DU145 and LNCaP cells were seed in 96-well plates (Corning Costar, Corning, NY) with 2.5 × 103 cells in each well for 24, 48, and 72 h. Then, MTT (20 μL; Sigma) was replenished to each well and maintained for 4 h, and 150 μL dimethyl sulfoxide was added for the dissolution of the formazan crystals. The Microplate Absorbance Reader (Thermo Fisher Scientific, Waltham, MA) was employed for the evaluation of the color reaction at 490 nm.
Transwell assay
The invasion assay of transfected DU145 and LNCaP cells was evaluated through the transwell chamber (8 μm; Corning Costar) involving the matrigel matrix (BD Biosciences, San Jose, CA). The migration assay of transfected DU145 and LNCaP cells was similar to those of the invasion assay, but did not contain the matrigel matrix. In short, RPMI 1640 medium supplemented with FBS (10%) was added as a chemoattractant to the lower chamber. Contemporaneously, transfected DU145 or LNCaP cells (1.5 × 105) in RPMI 1640 medium without FBS were added to the upper chamber.
After culture for 48 h, the migrated or invaded cells were fixed by methanol (100%) on the lower surface of the membrane, followed by replenishing 0.1% crystal violet for staining of the migrated or invaded DU145 and LNCaP cells. At last, an inverted microscope (Olympus, Tokyo, Japan) was employed to count the migrated or invaded DU145 and LNCaP cells.
Dual-luciferase reporter assay
The starBase v2.0 database was utilized for the prediction of the binding sites between ZEB1-AS1 or CUL4B and miR-342-3p. After that, wild-type 3′-untranslated regions (UTR), CUL4B, mutant 3′-UTR CUL4B, wild-type ZEB1-AS1, and mutant ZEB1-AS1 sequences containing putative miR-342-3p binding sites were synthesized. Then, these sequences were inserted into the pGL3-control luciferase reporter vector (Promega, Madison, WI) for the construction of the luciferase reporter vectors CUL4B 3′-UTR-WT, CUL4B 3′-UTR-MUT, ZEB1-AS1 WT, and ZEB1-AS1 MUT. Following this, the luciferase reporter vector or control vector was co-transfected into DU145 and LNCaP cells with miR-342-3p or miR-con, respectively. In the end, the luciferase activities of luciferase reporter vectors were evaluated by dual-luciferase reporter assay kit (Promega).
Statistical analysis
SPSS 18.0 software (SPSS, Chicago, IL) was utilized for the performance of the statistical analysis. Data were exhibited as mean ± standard deviation. Differences with p < 0.05 were statistically significant. Student's t test or one-way variance analysis (ANOVA) was employed for comparing the differences between two or among multiple groups. The correlation between ZEB1-AS1 and CUL4B was analyzed using Spearman's correlation analysis.
Results
ZEB1-AS1 and CUL4B were elevated and positively correlated in PCa tissues
To inquire into the expression pattern of ZEB1-AS1 and CUL4B in PCa, quantitative real-time polymerase chain reaction (qRT-PCR) was first executed for the detection of the expression levels of ZEB1-AS1 and CUL4B in 30 PCa tissues and paired normal PCa tissues. The results exhibited that ZEB1-AS1 and CUL4B were remarkably augmented in PCa tissues compared with those in paired normal PCa tissues (Fig. 1A, B).
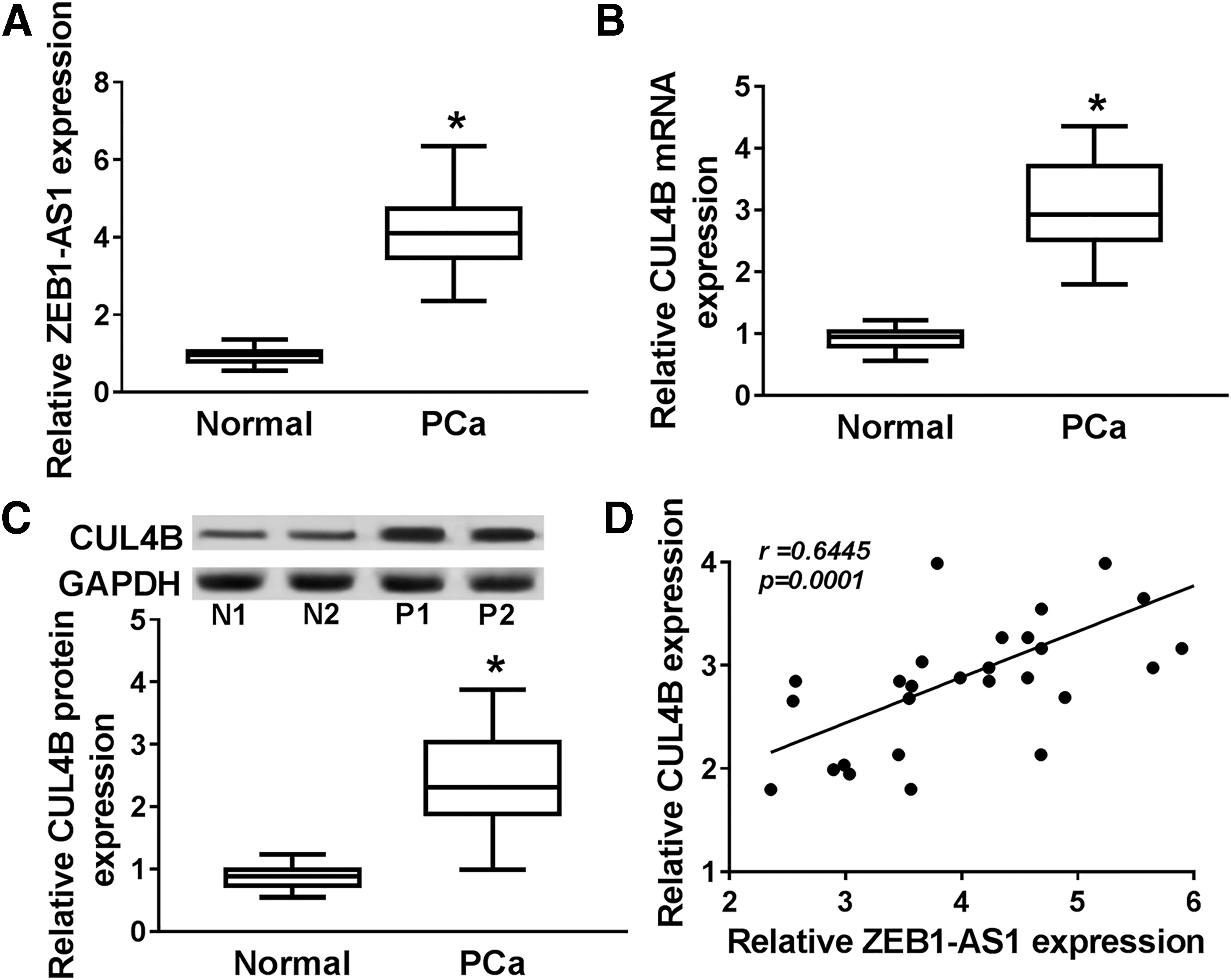
ZEB1-AS1 and CUL4B were elevated and positively correlated in PCa tissues.
Subsequently, the protein level of CUL4B in 30 PCa tissues and paired normal PCa tissues was evaluated by Western blot analysis. The results displayed that a prominent elevation of CUL4B protein was observed in PCa tissues compared with paired normal PCa tissues (Fig. 1C). The correlation between the expression of ZEB1-AS1 and CUL4B in PCa tissues was analyzed by Spearman's correlation analysis. The authors found that the expression of ZEB1-AS1 and CUL4B was positively correlated in PCa tissues (Fig. 1D). Therefore, ZEB1-AS1 and CUL4B were enhanced in PCa tissues, indicating that high expression of ZEB1-AS1 and CUL4B might be associated with the development of PCa.
Silencing of ZEB1-AS1 constrained proliferation, migration, and invasion of PCa cells
In consideration of the above results, the role of ZEB1-AS1 in PCa was further explored. In the first place, the expression level of ZEB1-AS1 in PCa cells and RWPE-1 cells was measured with qRT-PCR. As presented in Figure 2A, ZEB1-AS1 was evidently elevated in DU145 and LNCaP cells versus RWPE-1 cells. Subsequently, the si-ZEB1-AS1 (si-ZEB1-AS1#1 and si-ZEB1-AS1#2) or si-con was transfected into DU145 and LNCaP cells to silence the expression of ZEB1-AS1, respectively, and si-con served as the control group. Results of qRT-PCR indicated that a significant reduction of ZEB1-AS1 expression was discovered in DU145 and LNCaP cells transfected with si-ZEB1-AS1 compared with the control group, and the expression of ZEB1-AS1 was lower in the si-ZEB1-AS1#1 group than that in the si-ZEB1-AS1#2 group (Fig. 2B).

Effects of ZEB1-AS1 silencing on the proliferation, migration, and invasion of PCa cells.
Also, the authors selected si-ZEB1-AS1#1 for subsequent studies. In addition, the effects of ZEB1-AS1 silencing on cell proliferation, migration, and invasion in PCa cells were investigated. MTT assay was performed and the results showed that the reduction of ZEB1-AS1 expression dramatically repressed the proliferation of DU145 and LNCaP cells (Fig. 2C, D). Transwell migration and invasion assays exhibited that decreased ZEB1-AS1 expression conspicuously impeded the migration and invasion of DU145 and LNCaP cells (Fig. 2E, F).
In addition, the proliferation-associated proteins p21 and CyclinD1, as well as the migration and invasion-associated proteins MMP9 and E-cadherin, were evaluated to further validate the effects of ZEB1-AS1 downregulation on the proliferation, migration, and invasion of PCa cells. Results of Western blot analysis revealed that the reduction of ZEB1-AS1 expression in DU145 and LNCaP cells caused a significant decrease in protein levels of CyclinD1 and MMP9, but caused a marked increase in protein levels of p21 and E-cadherin (Fig. 2G, H). Together, these data indicated that the downregulation of ZEB1-AS1 suppressed cell proliferation, migration, and invasion in PCa cells.
Knockdown of CUL4B blocked proliferation, migration, and invasion of PCa cells
To explore the role of CUL4B in PCa, the mRNA and protein levels of CUL4B in PCa cells were evaluated using qRT-PCR or Western blot analysis. The results presented that mRNA and protein levels of CUL4B were distinctly increased in DU145 and LNCaP cells compared with RWPE-1 cells (Fig. 3A, B). Next, we examined the levels of CUL4B mRNA and protein in DU145 and LNCaP cells transfected with si-CUL4B or si-NC through qRT-PCR or Western blot analysis. The results displayed that the levels of CUL4B mRNA and protein were overtly reduced in DU145 and LNCaP cells after si-CUL4B transfection, indicating that the si-CUL4B could be used for subsequent studies (Fig. 3C, D).
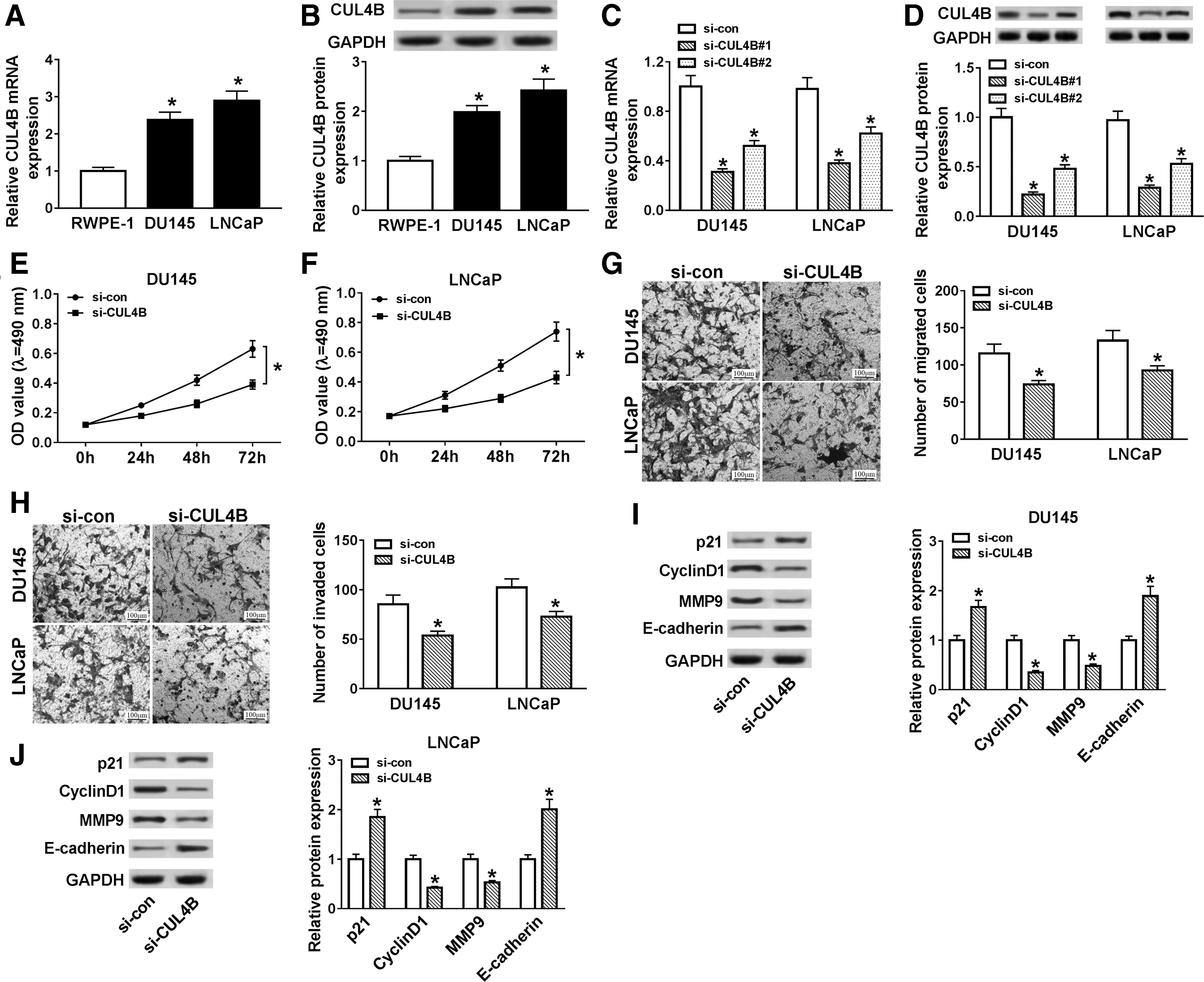
Effects of CUL4B inhibition on cell proliferation, migration, and invasion in PCa cells.
After that, the effects of CUL4B downregulation on the proliferation, migration, and invasion of PCa cells were further studied. The results of MTT assay indicated that the reduction of CUL4B apparently hampered the proliferation ability of DU145 and LNCaP cells (Fig. 3E, F). Moreover, transwell assay exhibited that the migration and invasion abilities of DU145 and LNCaP cells were dramatically hindered by CUL4B silencing (Fig. 3G, H). In addition, the protein levels of p21, CyclinD1, MMP9, and E-cadherin were detected with Western blot analysis. The authors found that the knockdown of CUL4B in DU145 and LNCaP cells remarkably reduced the protein levels of CyclinD1 and MMP9, and enhanced the protein levels of p21 and E-cadherin (Fig. 3I, J). Therefore, these results suggested that the downregulation of CUL4B impeded proliferation, migration, and invasion of PCa cells.
Overexpression of CUL4B weakened the effects of ZEB1-AS1 silencing on the proliferation, migration, and invasion of PCa cells
In view of the above results, the relationship between ZEB1-AS1 and CUL4B in PCa cells was further explored. At the outset, the effect of ZEB1-AS1 on the protein expression of CUL4B was detected by Western blot analysis. The results indicated that elevated ZEB1-AS1 expression distinctly enhanced the protein expression level of CUL4B in both DU145 and LNCaP cells. However, the reduction of ZEB1-AS1 obviously suppressed the protein level of CUL4B in DU145 and LNCaP cells (Fig. 4A, B). Moreover, the expression level of CUL4B protein was strikingly increased in both DU145 and LNCaP cells transfected with pcDNA-CUL4B compared with the control group (Fig. 4C).
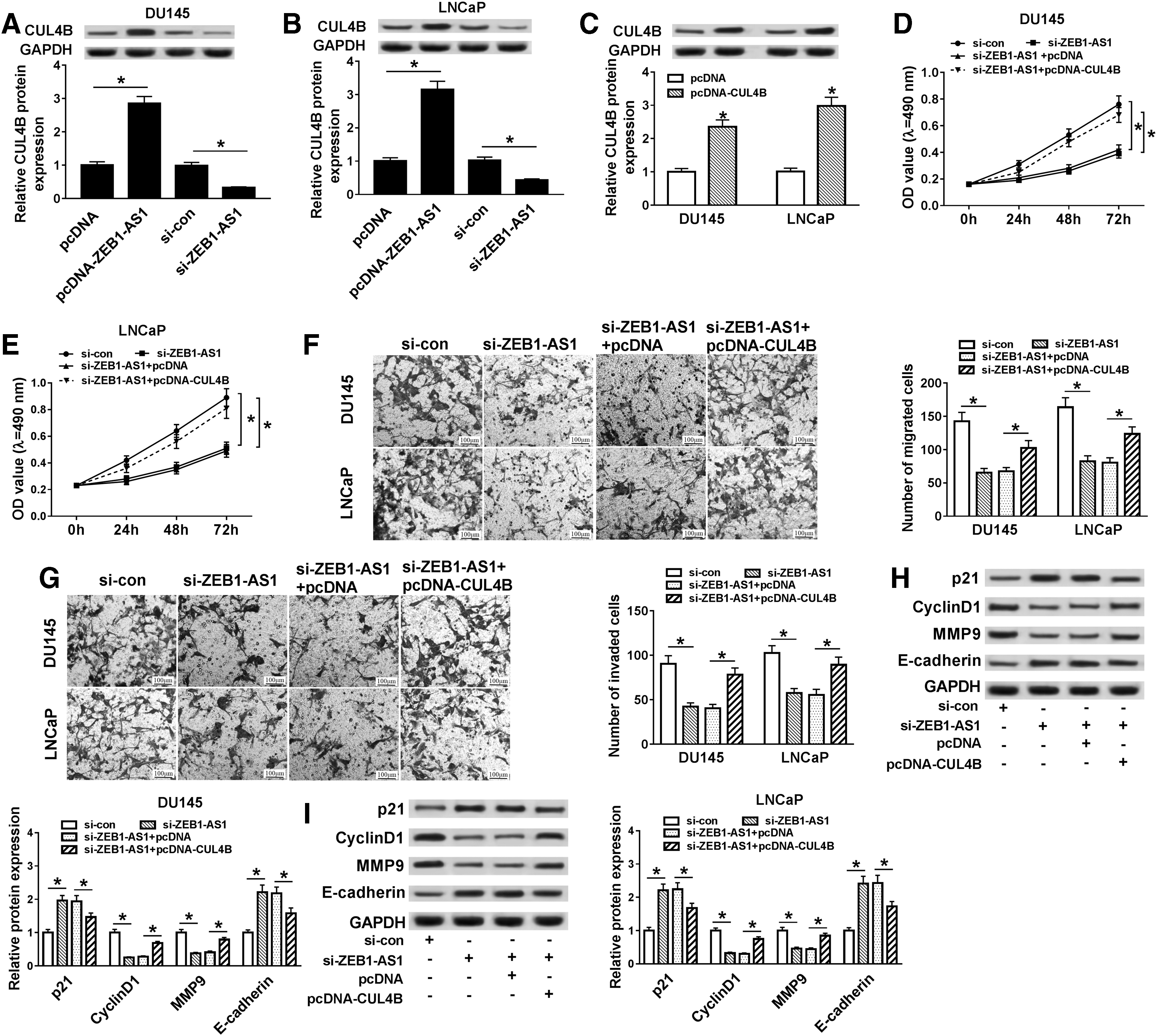
CUL4B augmentation weakened the effects of ZEB1-AS1 silencing on the proliferation, migration, and invasion of PCa cells.
Subsequently, it was further discussed whether ZEB1-AS1 affects cell proliferation, migration, and invasion by regulating the protein expression of CUL4B in DU145 and LNCaP cells. MTT assay revealed that the inhibition of cell proliferation caused by ZEB1-AS1 downregulation was reversed by the overexpression of CUL4B in DU145 and LNCaP cells (Fig. 4D, E). Consistently, transwell assay also indicated that elevation of CUL4B overturned the inhibition of migration and invasion of DU145 and LNCaP cells induced by ZEB1-AS1 reduction (Fig. 4F, G).
After that, Western blot analysis was performed to evaluate the proliferation-associated protein p21 and CyclinD1, as well as the expression of migration- and invasion-associated protein MMP9 and E-cadherin, to verify that ZEB1-AS1 affects PCa cell proliferation, migration, and invasion by regulating CUL4B expression. The results exhibited that the reduction in protein levels of CyclinD1 and MMP9 caused by ZEB1-AS1 knockdown was partly recovered by the introduction of CUL4B in DU145 and LNCaP cells. Concurrently, the enhancement of p21 and E-cadherin protein levels due to silencing of ZEB1-AS1 was partially reverted by elevating the CUL4Blevel (Fig. 4H, I). Taken together, these data indicated that the augmentation of CUL4B could weaken effects of ZEB1-AS1 silencing on the proliferation, migration, and invasion of PCa cells.
ZEB1-AS1 regulated CUL4B expression by binding to miR-342-3p in PCa cells
In the light of the above results, the authors further explored whether ZEB1-AS1 can act as a competing endogenous RNA (ceRNA) to regulate the expression of CUL4B in PCa cells. First, the starBase v2.0 database was utilized for the prediction of the possible sites where ZEB1-AS1 binds to miRNA. As presented in Figure 5A, miR-342-3p was predicted to have base pairing complementary to ZEB1-AS1. To validate this prediction, they constructed wild-type ZEB1-AS1 (ZEB1-AS1 WT) and mutant ZEB1-AS1 (ZEB1-AS1 MUT) pGL3 luciferase reporter vectors. The dual-luciferase reporter assay was then performed and the results displayed that the luciferase activity of ZEB1-AS1 WT co-transfected with miR-342-3p was strikingly reduced compared to the control group. However, there was no marked difference in luciferase expression of co-transfection with ZEB1-AS1 MUT and miR-342-3p (Fig. 5B, C).
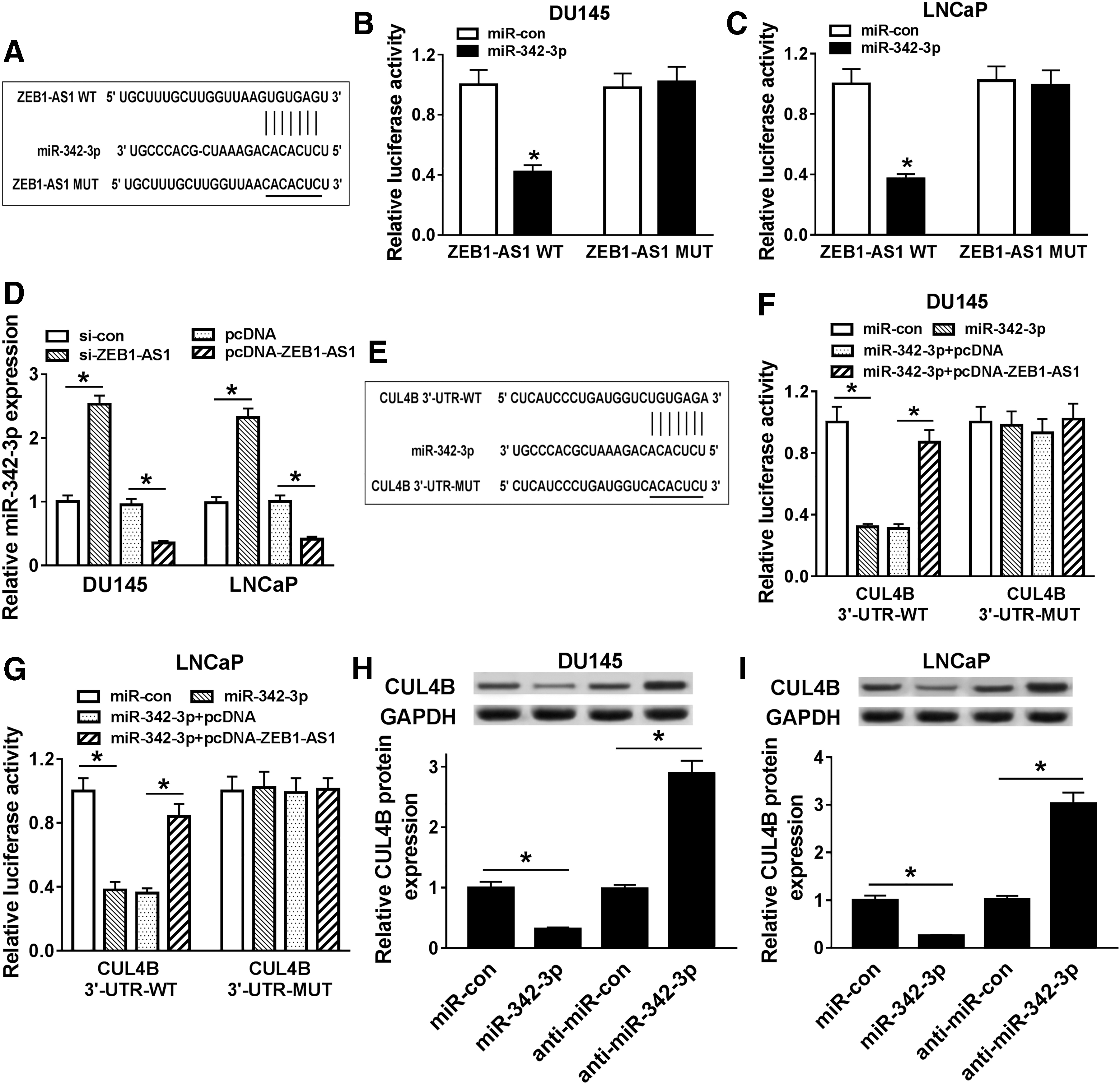
ZEB1-AS1 bound to miR-342-3p to regulate CUL4B in PCa cells.
Also, the effect of ZEB1-AS1 on the expression of miR-342-3p was assessed by qRT-PCR. The results exhibited that the expression of miR-342-3p was decreased in ZEB1-AS1-augmented DU145 and LNCaP cells, but was enhanced in ZEB1-AS1-silenced DU145 and LNCaP cells (Fig. 5D). In addition, the starBase v2.0 database also showed that miR-342-3p might bind to UTR of CUL4B mRNA (Fig. 5E). Following this, the authors established the wild-type CUL4B luciferase reporter vector (CUL4B 3′-UTR-WT) containing the predicted miR-342-3p binding region sites and the luciferase reporter vector for mutant CUL4B (CUL4B 3′-UTR-MUT). Results of dual-luciferase reporter assay revealed that co-transfection of miR-342-3p and CUL4B 3′-UTR-WT prominently repressed the luciferase activity in both DU145 and LNCaP cells compared the control group, while this suppression was restored by the introduction of ZEB1-AS1. However, for cells with CUL4B 3′-UTR-MUT, there was no apparent difference of luciferase activity in DU145 and LNCaP cells (Fig. 5F, G).
Besides, whether ZEB1-AS1 regulated CUL4B expression by miR-342-3p was further studied. Results of Western blot analysis presented that miR-342-3p elevation resulted in a reduction in CUL4B protein expression in both DU145 and LNCaP cells, and reduced miR-342-3p expression resulted in an increased protein expression of CUL4B in DU145 and LNCaP cells (Fig. 5H, I). In sum, ZEB1-AS1 could upregulate CUL4B expression by sponging miR-342-3p in PCa cells.
MiR-342-3p elevation curbed cell proliferation, migration, and invasion in PCa cells
To verify the effect of miR-342-3p in PCa, we first assessed miR-342-3p expression in PCa cells by using qRT-PCR. The authors discovered that miR-342-3p was conspicuously downregulated in DU145 and LNCaP cells compared to the RWPE-1 cells (Fig. 6A). Moreover, the expression of miR-342-3p in DU145 and LNCaP cells transfected with miR-342-3p was substantially enhanced (Fig. 6B). Next, the effects of miR-342-3p on cell proliferation, migration, and invasion in PCa cells were assessed. MTT assay revealed that miR-342-3p enhancement repressed the proliferation of DU145 and LNCaP cells (Fig. 6C, D). Furthermore, transwell assay demonstrated that upregulation of miR-342-3p suppressed cell migration and invasion in PCa cells (Fig. 6E, F).
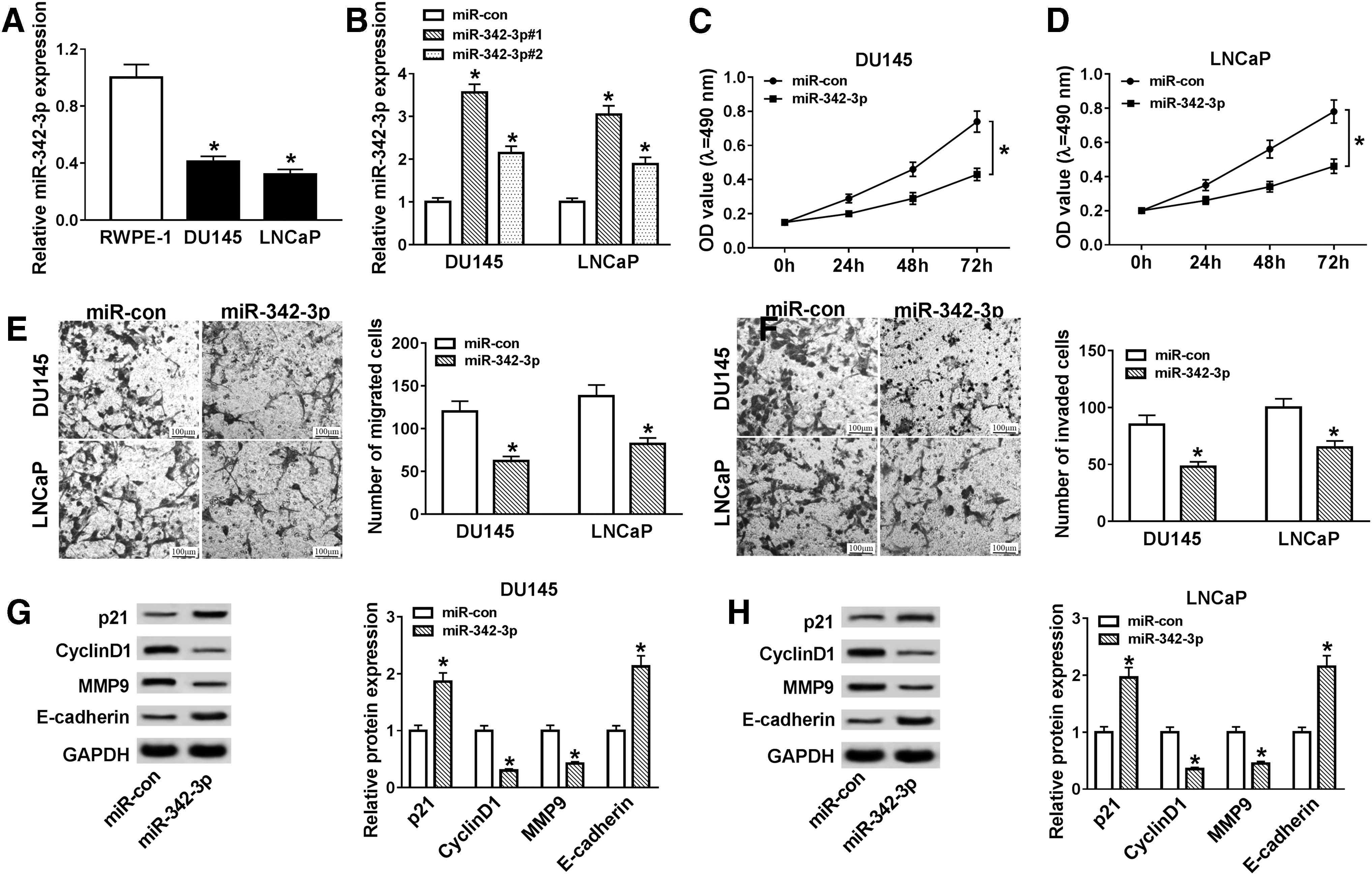
Effects of miR-342-3p on proliferation, migration, and invasion of PCa cells.
Besides, Western blot analysis disclosed that augmented miR-342-3p expression reduced the protein levels of CyclinD1 and MMP9 and increased the protein levels of p21 and MMP9 in DU145 and LNCaP cells (Fig. 6G, H). In sum, these findings suggested that miR-342-3p could impede cell proliferation, migration, and invasion in PCa cells.
ZEB1-AS1 regulated CUL4B expression to mediate PI3K/AKT/mTOR signaling in PCa cells
It has been reported that the occurrence of multiple cancers is associated with abnormal activation of the PI3K/AKT/mTOR signaling. 30 In view of the above results, we explored whether ZEB1-AS1 regulated the proliferation, migration, and invasion of PCa cells by CUL4B/PI3K/AKT/mTOR pathway. They observed that the reduction of ZEB1-AS1 markedly depressed the protein levels of p-PI3K, phosphorylated protein kinase B (p-AKT), and mammalian target of rapamycin (p-mTOR) in both DU145 and LNCaP cells, while these effects were overturned after CUL4B overexpression (Fig. 7A, B).
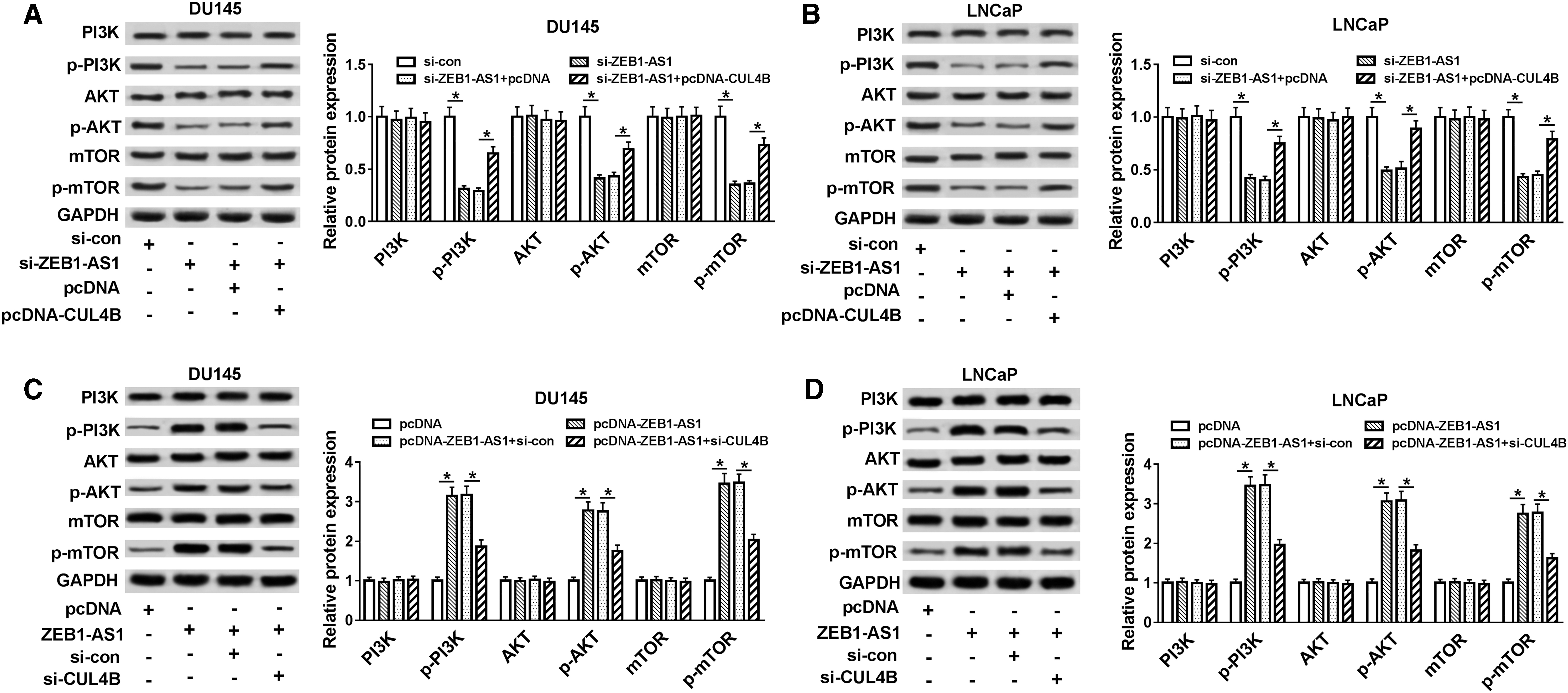
ZEB1-AS1 affected PI3K/AKT/mTOR signaling by regulating CUL4B expression.
Moreover, the protein levels of p-PI3K, p-AKT, and p-mTOR were markedly enhanced in DU145 and LNCaP cells transfected with pcDNA-ZEB1-AS1, while this elevation was overturned by CUL4B silencing (Fig. 7C, D). Taken together, these data revealed that ZEB1-AS1 mediated the PI3K/AKT/mTOR signaling by CUL4B in PCa cells.
Discussion
PCa has high morbidity and mortality, and poses a considerable threat to men's health. 31 Hence, the identification of novel molecular mechanisms connected with PCa progression was essential for the development of specific biomarker and therapeutic strategies. Increasing evidence indicated that abnormal expression of lncRNA was concerned with tumorigenesis. 32,33 Accumulated evidence has proved that ZEB1-AS1 was correlated with the pathogenesis of diverse tumors. For instance, the elevation of ZEB1-AS1 accelerated the metastasis of hepatocellular cancer and predicted a poor prognosis. 9 Moreover, ZEB1-AS1 was evidently enhanced in colorectal cancer, and reduced ZEB1-AS1 expression facilitated apoptosis and constrained proliferation of colorectal cancer cells. 10 Also, ZEB1-AS1 was revealed to be elevated in glioma tissues and cells, and decreased ZEB1-AS1 expression repressed the metastasis and growth of glioma cells in vitro and in vivo. 11 In this study, a prominent enhancement of ZEB1-AS1 was observed in PCa tissues and cells. Inhibition of ZEB1-AS1 retarded proliferation, migration, and invasion of PCa cells. Su et al. proclaimed that the upregulation of ZEB1-AS1 expedited migration and proliferation of PCa cells. 34 CyclinD1 is a key cell cycle regulator that modulates G1 and S phase transitions. 35 Moreover, MMP9 was involved in tumor aggressiveness. 36 P21 could promote cell cycle arrest in a variety of stimuli. 37 Also, E-cadherin plays a vital role in epithelial cell behavior and cancer suppression. 38 Herein, inhibition of ZEB1-AS1 reduced the protein levels of CyclinD1 and MMP9 and elevated the protein levels of p21 and E-cadherin. The authors' results were consistent with a study by Xu et al., 32 indicating that augmented ZEB1-AS1 expression played a carcinogenic role in PCa.
It has been reported that ZEB1-AS1 could acts as a ceRNA binding to miRNA to regulate the biological function of miRNA. 10 –12 For example, Meng et al. claimed that ZEB1-AS1 could directly modulate miR-200c/141 in glioma cells. 11 To investigate whether ZEB1-AS1 acts as a ceRNA in PCa cells, starBase v2.0 database was employed and the results exhibited that ZEB1-AS1 possessed binding sites for miR-342-3p. Previous studies presented that miR-342-3p exerted an anticancerous role in gallbladder cancer, nonsmall cell lung cancer, and nasopharyngeal cancer. 20 –22 For instance, elevated miR-342-3p expression repressed cell colony formation, proliferation, and invasion in nasopharyngeal cancer. 21 Also, miR-342-3p upregulation impeded cell migration and proliferation in nonsmall cell lung cancer. 22 In this study, the authors found that miR-342-3p was a target for ZEB1-AS1, and ZEB1-AS1 positively regulated miR-342-3p expression in PCa cells. And miR-342-3p overexpression repressed cell proliferation, migration, and invasion of PCa cells. Consistent with the above studies, these results suggested that miR-342-3p exerted antitumor role in PCa.
In a general way, miRNAs play roles in different cancers by targeting the target gene. Because of this, the authors proved that CUL4B was a target gene of miR-342-3p by starBase v2.0 database and dual-luciferase reporter assay. Moreover, CUL4B was enhanced in PCa tissues and cells, and silencing of CUL4B blocked cell proliferation, migration, and invasion in PCa cells. Literature of Qi et al. stated that CUL4B was increased in PCa tissues, and elevated CUL4B expression predicted a poor prognosis. 28 Also, Zhao et al. also stated that CUL4B was augmented in PCa tissues and cells, and the elevation of CUL4B accelerated cell aggressiveness and proliferation in PCa cells. 39 This study proved that CUL4B was an oncogene in PCa and was consistent with studies of Qi et al. and Zhao et al. In addition, they discovered that ZEB1-AS1 regulated CUL4B expression by binding to miR-342-3p in PCa cells by Western blot analysis. Augmentation of CUL4B expression attenuated the effects of ZEB1-AS1 downregulation on the proliferation, migration, and invasion of PCa cells.
Activation of the PI3K/AKT/mTOR signaling pathway was discovered in diverse cancers, including PCa. 40 Liang et al. reported that GPCR48/LGR4 facilitated PCa progression by activating the PI3K/AKT pathway. 41 Besides, GOLM1 expedited the progression of PCa by activating the signaling of PI3K/AKT/mTOR. 42 Also, enhanced TIPE2 inhibited cell invasion, proliferation, and migration by suppressing PI3K/AKT signaling in PCa cells. 43 In this study, the authors found that ZEB1-AS1 mediated the PI3K/AKT/mTOR signaling through CUL4B in PCa cells. Nevertheless, the roles and mechanisms of ZEB1-AS1 and CUL4B in this study were only studied in PCa cell model in vitro. In the future, the role and the regulatory mechanism of ZEB1-AS1 can be further confirmed by establishing a xenograft model.
Conclusions
In conclusion, ZEB1-AS1 and CUL4B were remarkably augmented in PCa tissues and cells. Furthermore, knockdown of ZEB1-AS1 suppressed proliferation, migration, and invasion of PCa cells through ZEB1-AS1/miR-342-3p/CUL4B axis-mediated PI3K/AKT/mTOR signaling.
Authors' Contributions
T.M. and H.C. conceived, designed, and revised this study. P.W., N.Y., and J.B. analyzed the data and wrote the article. All authors read and approved the final article.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.