
Abstract
Objective:
TP73-AS1 has been reported as an overexpressed oncogenic lncRNA in several types of cancer. However, these analyses of The Cancer Genome Atlas data set revealed downregulation of TP73-AS1 in acute myeloid leukemia (AML). In this study, we aimed to study the molecular mechanism between TP73-AS1 and cell proliferation in AML.
Methods:
Bone marrow (BM) samples were obtained from 50 AML patients and 50 healthy controls. Cell transient transfections were performed to analyze gene interactions. Dual-luciferase reporter assay, quantitative polymerase chain reaction and Western blot were used to study the gene expressions. Cell proliferation was analyzed by CCK-8 method.
Results:
TP73-AS1 was confirmed to be downregulated in AML. TP73-AS1 was predicted to interact with miR-21, while overexpression of TP73-AS1 and miR-21 did not affect the expression of each other. Instead, overexpression of TP73-AS1 led to the upregulation of phosphatase and tensin homologue (PTEN), a downstream target of miR-21. Cell proliferation analysis showed that overexpression of TP73-AS1 and PTEN led to a decreased proliferation rate of AML cells. Overexpression of miR-21 played an opposite role and reduced the effects of overexpressing TP73-AS1 and PTEN.
Conclusion:
TP73-AS1 may regulate the miR-21/PTEN axis to affect cell proliferation in AML.
Introduction
Acute myeloid leukemia (AML) is the major subtype of acute leukemia and accounts for more than 80% of all acute leukemia cases. 1 –3 Increased incidence of AML is observed with aging, from 1.3 per 100,000 people among adults younger than 60 years to 12.2 per 100,000 people among adults older than 65 years. 3 With the development of anticancer therapies, survival of younger AML patients has been significantly improved. However, only 30% of AML patients who are older than 65 years survive greater than 1 year after the initial diagnosis. 4,5 Therefore, novel therapeutic approaches are of great importance.
Previous studies on the biology of AML have revealed that genetic alterations play critical roles in the development and progression of AML. 6 –8 Studies on the molecular pathogenesis of AML have provided novel insights into the development of novel therapies, such as targeted therapy. 6 –8 It has been observed that the occurrence of AML is frequently accompanied by the dysregulation of phosphatase and tensin homologue (PTEN). 9,10
PTEN is a well-studied tumor suppressor that suppresses tumor growth mainly by inactivating the PI3K-Akt pathway, which is one of the major survival pathways in cancer biology. 11 In effect, the well-characterized oncogenic miR-21 plays its roles in cancer biology mainly by targeting PTEN. 12 The bioinformatic analyses showed that miR-21 may interact with TP73-AS1, which is a characterized oncogenic lncRNA in several types of cancer, such as liver cancer. 13 By analyzing The Cancer Genome Atlas (TCGA) data set, the authors observed the downregulation of TP73-AS1 in AML. Therefore, this study was performed to explore the involvement of TP73-AS1 and its interactions with miR-21 in AML.
Materials and Methods
Participants and bone marrow
This study passed the review of the Ethics Committee of the First Affiliated Hospital of Kunming Medical University (no. FAH20160423896532KMU).
Bone marrow (BM) samples were obtained from 50 AML patients (30 males and 20 females, 51–75 years old, mean age 63.6 ± 5.1 years) and 50 healthy controls (30 males and 20 females, 51–75 years old, mean age 63.8 ± 5.3 years) who were admitted at the aforementioned hospital between May 2016 and May 2019. AML with maturation (M2) is a subtype of AML. In this study, 40 patients were diagnosed as M2. All AML patients are newly diagnosed cases and no other severe clinical disorders were diagnosed. No therapies were initiated before the collection of BM. BM mononuclear cells (BMMNCs) were separated from BM using the Lymphocyte Separation Medium (TBD Sciences). All samples of BMMNCs were stored in liquid nitrogen before use.
AML cell line and transient transfections
The AML human Kasumi-6 cell line (M2) from ATCC was used in this study. Cells were cultivated in Roswell Park Memorial Institute (RPMI)-1640 medium containing 10% fetal bovine serum and 100 U/mL penicillin/streptomycin at 37°C with 5% CO2. Cells used for transfections were harvested at 75%–85% confluence.
Expression vectors of TP73-AS1 and PTEN were constructed by RiboBio (Guangzhou, China) using pcDNA3.1 vector as the backbone. Negative control (NC) miRNA and miR-21 mimic were also synthesized by RiboBio. Lipofectamine 2000 (Invitrogen) was used to transfect siRNA (50 nM) or vector (10 nM) into 106 cells. Transfection efficiency was measured by quantitative polymerase chain reaction (qPCR) at 24 h post-transfection. Untransfected cells were used as control (C) cells. Cells transfected with NC miRNA or empty pcDNA3.1 vector were NC cells. Cells were collected at 24 h post-transfection to perform the following experiments.
Luciferase activity assay
pGL3 Luciferase Reporter Vector (Basic; Promega Corporation) was used as the backbone to construct TP73-AS1 luciferase vector. Transfections were performed to cotransfect TP73-AS1 vector + NC miRNA or TP73-AS1 vector + miR-21 mimic into 106 cells. Cells were harvested at 48 h post-transfection. Luciferase activity assay was performed using the Dual-Luciferase Reporter Assay System (Promega Corporation). Renilla luciferase activity was normalized to Firefly luciferase expression.
RNA extractions and RT-qPCR
Total RNAs were extracted from tissue samples and cultivated cells using the GenElute™ Total RNA Purification Kit (Sigma-Aldrich). RNA samples were digested by DNA eraser (Takara) to remove genomic DNAs. Reverse transfections were performed using the PrimeScript® RT reagent kit (TaKaRa). qPCRs were prepared using the QuantiFast SYBR Green PCR Kit (QIAGEN) to measure the expression levels of TP73-AS1 and PTEN. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as endogenous control. High Pure miRNA Isolation Kit (Sigma-Aldrich) was used to extract miRNAs. The expression levels of mature miR-21 were measured using the All-in-One™ miRNA qRT-PCR Reagent Kit (GeneCopoeia). Fold changes of gene expression were calculated using 2−ΔΔCT methods. All PCRs were repeated three times.
Western blot
Total proteins were extracted from cells harvested at 24 h post-transfection using RIPA buffer (Cell Signaling Technology). Protein concentrations were measured using the BCA Protein Assay Kit (Cell Signaling Technology). After protein denaturation at 95°C for 10 min, 12% sodium dodecyl sulfate/polyacrylamide gel was used to separate proteins, followed by gel transfer to polyvinylidene fluoride membranes. After blocking in phosphate-buffered saline containing 5% fat-free milk at room temperature for 2 h, membranes were incubated with rabbit primary antibodies of GAPDH (ab9845; Abcam) and PTEN (ab31392; Abcam) at 4°C for 18 h, followed by incubation with the HRP goat anti-rabbit (IgG) secondary antibody (ab6721; Abcam) at room temperature for 2 h.
Signals were produced using enhanced chemiluminescence (Santa Cruz Biotechnology), and data were processed using the ImageJ v1.46 software.
Cell proliferation assay
Cell proliferation was assessed at 24 h post-transfection using the Cell Counting Kit-8 kit (Dojindo). Briefly, each well of a 96-well plate was added with 0.1 mL cell suspension containing 104 cells. Cells were cultivated under aforementioned conditions and 0.01 mL WST-8 solution was added into each well at 4 h before cell collection. Cells were collected every 24 h for a total of 96 h. Absorbance was measured at 450 nm using a spectrophotometer.
Statistical analysis
Data of three biological replicates are expressed as mean ± standard deviation and were analyzed by the SPSS 18.0 software (SPSS, Inc.). Unpaired t-test was used to compare differences between two groups. Differences among multiple groups were compared by analysis of variance (ANOVA) (one-way) and Tukey test. p < 0.05 was statistically significant.
Results
TP73-AS1 was downregulated in AML
The TCGA data set was the first to explore the differential expression of TP73-AS1 in AML. It was observed that TP73-AS1 was significantly downregulated in AML in comparison with that in nontumor tissues (12.01 vs. 24.51). Downregulation of TP73-AS1 in AML was further confirmed by detecting the expression levels of TP73-AS1 in BMMNCs from both AML patients (n = 50) and healthy controls (n = 50) (Fig. 1). Unpaired t-test showed that the expression levels of TP73-AS1 were significantly lower in AML patients than that in the controls (p < 0.001).
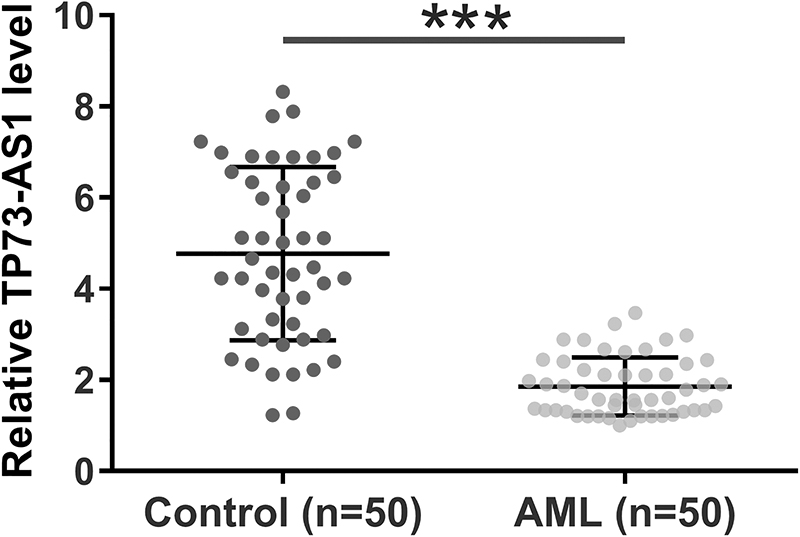
TP73-AS1 was downregulated in AML. The downregulation of TP73-AS1 in AML was analyzed by measuring the expression levels of TP73-AS1 in bone marrow mononuclear cells from both AML (n = 50) and controls (n = 50). Unpaired t test was used for data comparison. Polymerase chain reactions were repeated three times and data were expressed by mean values.***p < 0.001. AML, acute myeloid leukemia.
TP73-AS1 can directly interact with miR-21
The interactions between TP73-AS1 and human miRNAs in the miRBase database were predicted using the online RNA-RNA interaction prediction program IntaRNA. Among human miRNAs, TP73-AS1 was predicted to form strong base-pairing with miR-21 (Fig. 2A). Dual-luciferase reporter assay showed that compared with Kasumi-6 cells transfected with TP73-AS1 vector and miRNA NC (NC group), cells transfected with TP73-AS1 vector and miR-21 mimic had a significantly lower relative luciferase activity (Fig. 2B, p < 0.05).
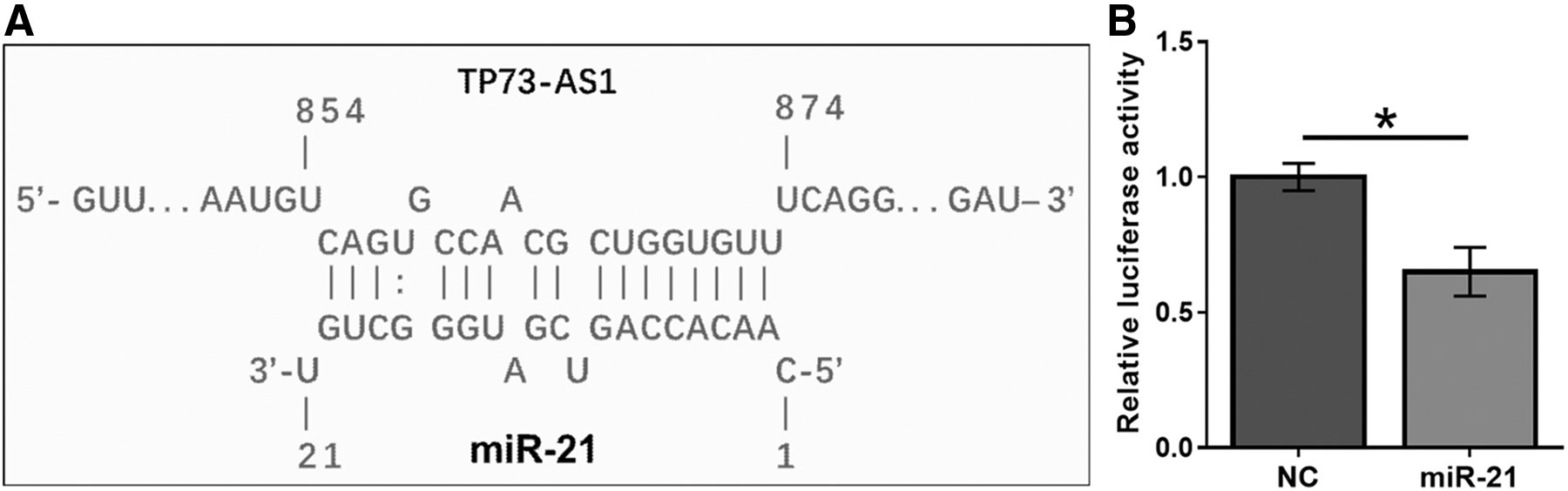
TP73-AS1 can directly interact with miR-21. The interactions between TP73-AS1 and human miRNAs from miRBase were analyzed using an online RNA-RNA interaction prediction program named IntaRNA. The potential base-paring formed by TP73-AS1 and miR-21 was presented
TP73-AS1 upregulates PTEN, a target of miR-21
Kasumi-6 cells were transfected with TP73-AS1 expression vector or miR-21 mimic to further assess the interaction between TP73-AS1 and miR-21. Overexpression of TP73-AS1 and miR-21 was confirmed by qPCR at 24 h post-transfection (Fig. 3A, p < 0.05). The effects of overexpressing TP73-AS1 and miR-21 on the expression of each other were also evaluated by qPCR at 24 h post-transfection. Compared with the NC and C groups, overexpression of TP73-AS1 and miR-21 did not affect the expression of each other (Fig. 3B, p < 0.05). Western blot and qPCR were performed to evaluate the effects of overexpressing TP73-AS1 on the expression of PTEN at both protein and mRNA levels, respectively. Compared with the C and NC groups, overexpression of TP73-AS1 led to upregulated PTEN at both protein (Fig. 3B) and mRNA (Fig. 3C) levels (p < 0.05).
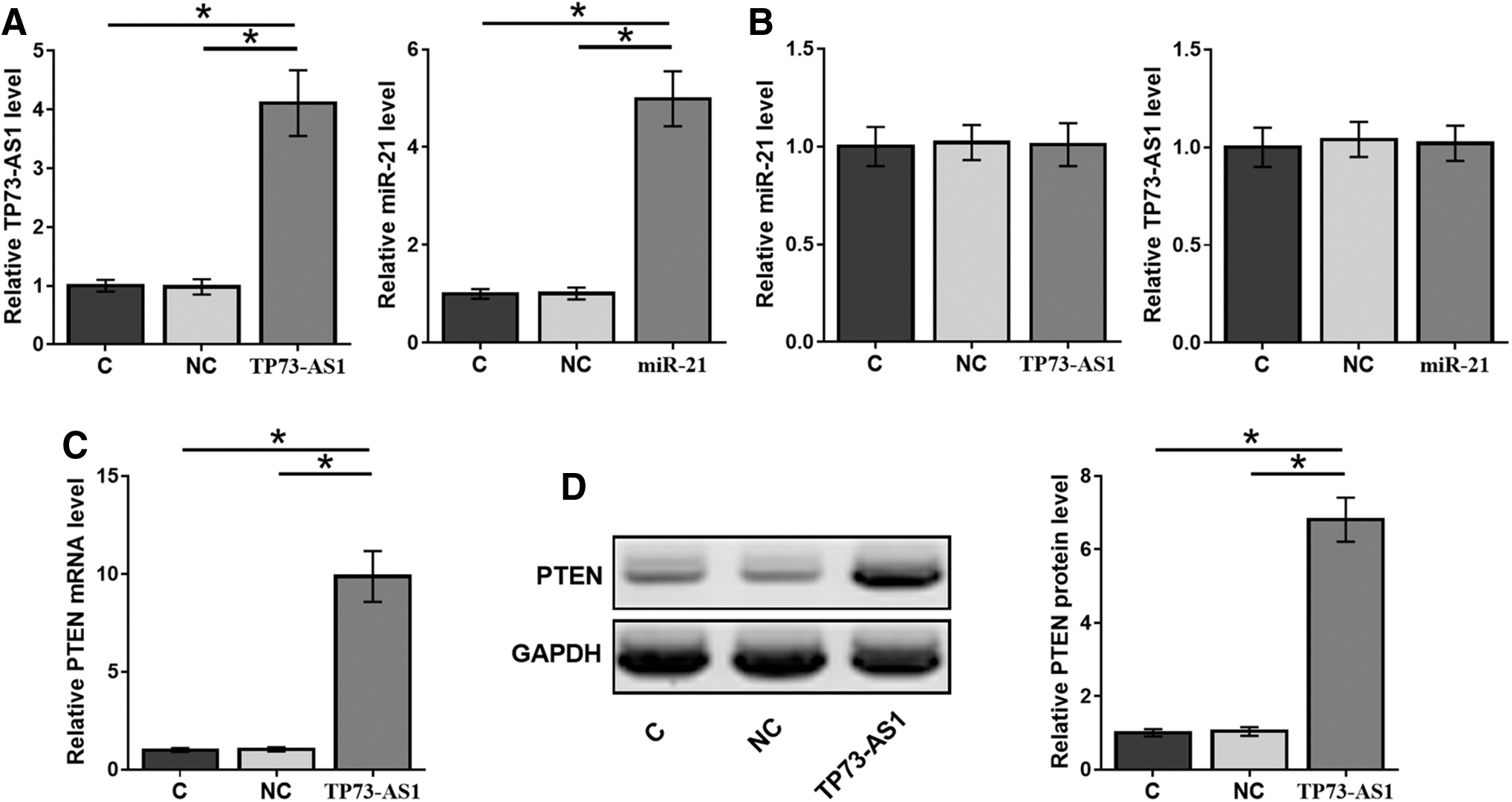
TP73-AS1 upregulates PTEN, a target of miR-21. Kasumi-6 cells were transfected with TP73-AS1 expression vector or miR-21 mimic to further analyze the interaction between TP73-AS1 and miR-21. Overexpression of TP73-AS1 and miR-21 was confirmed by qPCR at 24 h post-transfection
TP73-AS1 regulated miR-21/PTEN axis to suppress the proliferation of Kasumi-6 cells
The effects of overexpressing TP73-AS1, miR21, and PTEN on the proliferation of Kasumi-6 cells were assessed by CCK-8 assay. Compared with the C group, overexpression of TP73-AS1 and PTEN led to decreased proliferation rates of AML cells. Overexpression of miR-21 played an opposite role and reduced the effects of overexpressing TP73-AS1 and PTEN (Fig. 4, p < 0.05).
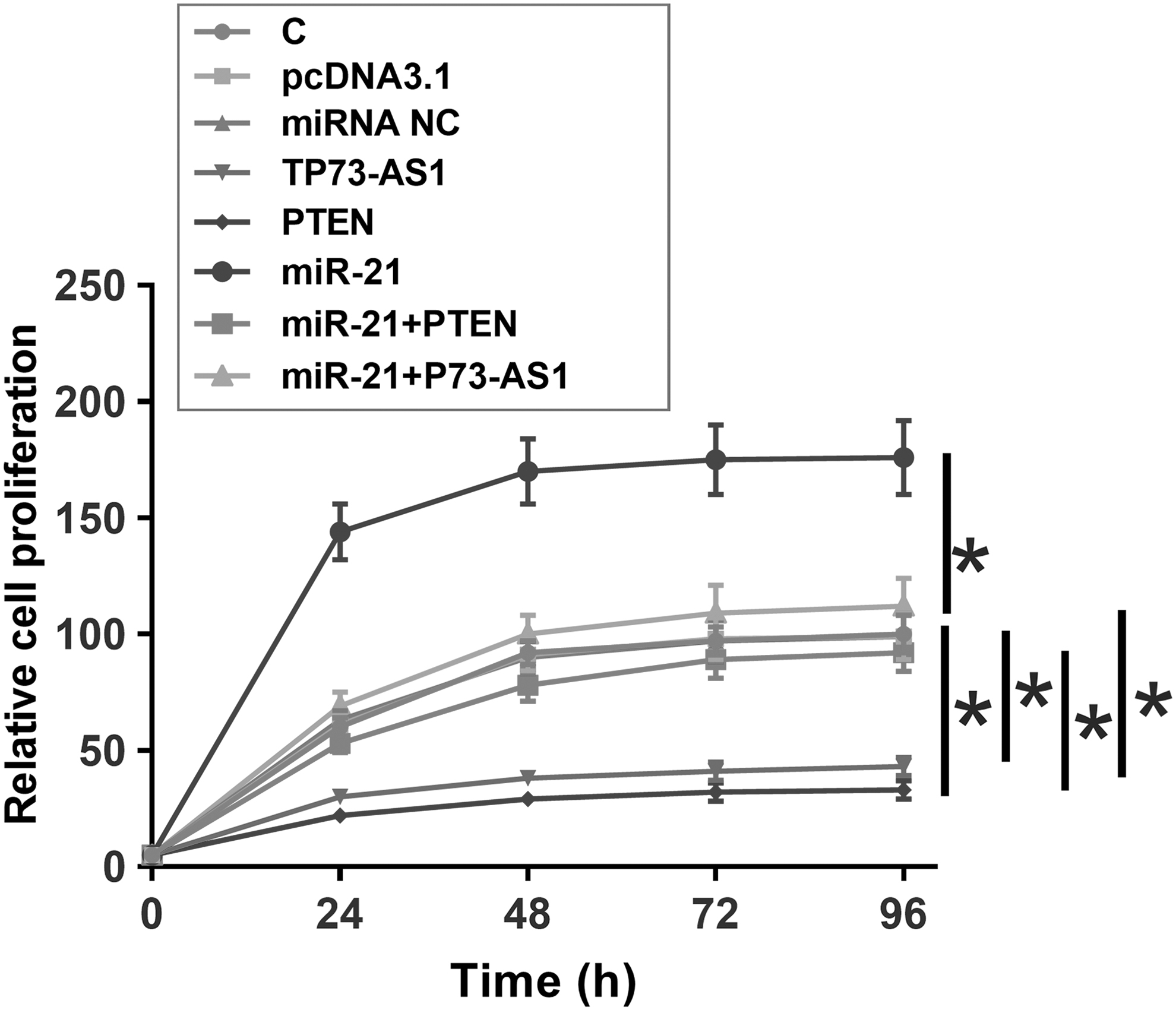
TP73-AS1 regulated miR-21/PTEN axis to suppress the proliferation of Kasumi-6 cells. The effects of TP73-AS1, miR21, and PTEN overexpression on the proliferation of Kasumi-6 cells were analyzed by CCK-8 assay. Experiments were repeated three times and data were expressed as mean values. *p < 0.05. PTEN, phosphatase and tensin homologue.
Discussion
The roles of TP73-AS1 have been reported in several types of cancer. To the best of the authors' knowledge, this study is the first to report the involvement of TP73-AS1 in AML. The authors found that TP73-AS1 could regulate miR-21/PTEN to regulate AML cell proliferation.
Previous studies have investigated the functions of TP73-AS1 in many types of cancers, such as liver cancer, esophageal squamous cell carcinoma, breast cancer, and lung cancer. 13 –16 TP73-AS1 is upregulated in these cancers and promotes cancer development by regulating cancer cell behaviors, such as promoting the proliferation of cancer cells. 13 –16 Knockdown of TP73-AS1 induces the apoptosis of cancer cells to suppress cancer development. 16 However, by analyzing the TCGA data set, the authors observed downregulation of TP73-AS1 in various cancers, such as colon cancer, nonsmall-cell lung cancer, and liver cancer. The inconsistency between TCGA data set and previous studies might be due to the discrepancy in the involved population. Future studies are needed to further elucidate the differential expression of TP73-AS1 in different types of cancer.
By analyzing the TCGA data set, the authors observed the downregulation of TP73-AS1 in AML, which was further confirmed by measuring its expression levels in AML patients and healthy controls included in this study. These results also showed a decreased proliferation rate of AML cells after the overexpression of TP73-AS1. These data suggest that TP73-AS1 is a tumor-suppressive lncRNA in AML.
The main role of miRNAs is to downregulate the expression of target genes. 17 This study showed the direct interaction between TP73-AS1 and miR-21, while overexpression of miR-21 did not affect the expression of TP73-AS1. Instead, overexpression of TP73-AS1 led to upregulation of PTEN, which is a well-established target of miR-21. Therefore, TP73-AS1 is likely an endogenous sponge of miR-21 to upregulate PTEN, thereby inhibiting the proliferation of AML cells. However, it might involve other mechanisms underlying the interactions between TP73-AS1 and miR-21. Further studies are needed to fully elucidate this regulation network.
In conclusion, TP73-AS1 is downregulated in AML and may regulate the miR-21/PTEN axis to suppress AML cell proliferation.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
This work was supported by the Fund of Yunnan Provincial Health Science and Technology Plan (grant no. 2016NS052).