
Abstract
Background:
Cervical cancer (CC) is one of the most common cancers among women worldwide. Circular RNAs (circRNAs) are recently identified as important gene regulators with critical roles in cancer biology. In this study, we explored the effects of circ_0000388 on the malignant phenotypes of CC cells and its mechanism.
Materials and Methods:
Circ_0000388 expression and miR-337-3p expression in CC tissue samples were measured using quantitative real time polymerase chain reaction. CCK-8 was adopted to assess the effect of circ_0000388 on CC cell line proliferation. TUNEL assay was employed to probe the effect of circ_0000388 on apoptosis. Wound healing assay and transwell assay were conducted to detect the effect of circ_0000388 on migration and invasion. Further, interaction among circ_0000388, miR-337-3p, and TCF12 (transcription factor 12) was determined by bioinformatics analysis, RT-PCR, Western blot, RNA immunoprecipitation assay, and luciferase reporter assay.
Results:
Circ_0000388 expression in CC clinical samples was upregulated and this was correlated with unfavorable pathological indexes. Circ_0000388 remarkably enhanced the proliferation and metastasis of CC cells. Circ_0000388 overexpression dramatically impeded miR-337-3p expression and it was identified as a sponge of miR-337-3p. Furthermore, circ_00003888 also enhanced the TCF12 expression, while the effect could be reversed by co-transfection with miR-377-3p.
Conclusions:
Circ_0000388 was a novel oncogenic circRNA in CC, and promoted cancer progression via regulating miR-337-3p and TCF12, and could be potentially used as a diagnostic biomarker and therapy target.
Introduction
Cervical cancer (CC) has become the fourth most common cancer in women worldwide, whereas, in Central and East Africa, the morbidity of CC in women has already ranked first in all cancers. 1 Although the combination of surgery, radiotherapy, and chemotherapy has significantly improved the survival time of CC patients, for CC patients who have progressed to the advanced stage, the 5-year survival rate is far from satisfactory. 2 Therefore, it is especially important to explore the molecular mechanisms of CC development and find sensitive early markers and more effective therapeutic targets.
Circular RNA (circRNA) belongs to noncoding RNA (ncRNA). Structurally, circRNA is a kind of RNA without 5′, 3′ ends and polyadenylation tails, which is not easily degraded by RNase. Functionally, circRNAs can regulate gene expression by affecting transcription, mRNA stability, or by affecting translation by acting on RNA-binding proteins and microRNA (miRNA). 3 Due to its stable structure, it is potential to be used as a marker of a variety of diseases. It has been found that circRNA is abnormally expressed in a variety of human tumors and plays an important role in the tumorigenesis and development of malignant tumors. 4 Meanwhile, a variety of circRNAs have been found to dramatically affect the progression of CC. For instance, circ_0023404, which is highly expressed in CC, interacts with miR-5047 and facilitates CC metastasis and improves its drug resistance by modulating VEGFA expression and autophagy 5 ; circ_0000263 high expression can enhance the proliferation and migration of CC cells and accelerate cancer progression 6 ; circ_0000745 can enhance the proliferation, migration, and invasion of CC cells. 7 However, at present, there is little research on the function of circ_0000388, and it remains largely unknown whether it takes part in CC progression.
miRNAs are ncRNAs with 19–25 nucleotides in length. Accumulating researches have found that miRNAs play important roles in diverse human tumors. For instance, the miR-146a/b expression is increased in papillary thyroid carcinoma and has a significant effect on promoting the growth and migration of cancer cells. 8 MiR-214 expression and miR-218 expression are decreased in breast cancer, and it has been proved that it can repress the proliferation of cancer cells and facilitate apoptosis, and slow down the progression of breast cancer. 9 Mounting researches imply that miR-337-3p serves as a tumor suppressor in diverse tumors. For instance, miR-337-3p targets JAK2 thereby suppressing the proliferation and invasion of liver cancer cells. 10 In gastric cancer, transfection of miR-337-3p mimics has been validated to notably repress the invasion of gastric cancer cells, and the downregulation of miR-337-3p expression is closely related to the metastasis of GC. 11 Additionally, studies have indicated that miR-337-3p can also restrain the CC progression to a certain extent and improve the prognosis of cancer, 12 but the upstream and downstream regulation mechanisms of miR-337-3p are far from being elucidated.
Transcription factor 12 (TCF12) is a member of the basic helix-loop-helix transcription factors, which is a protein with DNA binding ability. TCF12 is widely expressed in diverse tissues and organs of the human body and modulates cell differentiation. 13,14 There is increasing evidence that TCF12 has a dual role of tumor suppression or cancer promotion in different human cancers. In gastric cancer, highly expressed TCF12 enhances the proliferation, migration, and invasion of GC cells by activating PI3K/AKT signaling pathway. 15 In oral cancer, TCF12 can remarkably repress the activity of oral squamous cell carcinoma cells through regulating the transcription of FAM213A. 16 In addition, in CC, it has been found that TCF12 can enhance the migration and invasion of HeLa and C33A cells, and participate in promoting cancer progression, 17 but the upstream regulation mechanism of TCF12 has not been elucidated clearly.
There are wide-ranging effects of circRNA on miRNA expression and activity, and miRNAs have been shown to interfere with the post-transcriptional translation process by binding to the 3′ UTR end of specific mRNAs. 3,18 In view of this, we are curious about the role of circRNA-miRNA-mRNA regulatory networks in CC. Through CircInteractome database, we found that miR-337-3p was one of the downstream targets of circ_0000388, while data from TargetScan database project that miR-337-3p can target on TCF12. Based on the above data, we made a hypothesis that circ_0000388 could play an essential role in the progression of CC by targeting the miR-337-3p/TCF12 axis. Therefore, this work will probe the role of circ_0000388, miR-337-3p, and TCF12 in CC and its mutual regulation mechanism to further explain the molecular basis of the development of CC and explore new therapeutic targets for CC.
Materials and Methods
Clinical samples
CC tissue samples and adjacent normal tissues were collected from 40 CC patients in Linyi Central Hospital from April 2017 to April 2018. The samples have complete clinical and pathological records. The study protocol was approved by the Ethics Committee of Linyi Central Hospital.
Cell culture and transfection
Human CC cells (HeLa, SiHa, Caski, C-33A, and MS751 cells) and normal human cervical epithelial cells were purchased from the Cell Center of the Chinese Academy of Sciences (Shanghai, China). The cells were cultured in RPMI 1640 medium (Thermo Fisher Scientific, MA) containing 10% fetal bovine serum (FBS; Thermo Fisher Scientific) and 1% penicillin/streptomycin (Invitrogen, Carlsbad, CA) and cultured in an incubator at 37°C in CO2 (volume fraction of 5%). The cells in the logarithmic growth phase were trypsinized and subcultured with 0.25% trypsin (Thermo Fisher HyClone, UT). Following that, the shRNA for circ_0000388, overexpressing pcDNA3.1-circ_0000388 and their corresponding negative controls, miR-337-3p mimics, inhibitors, and miR negative control was synthesized (RiboBioCo., Guangzhou, China). Cell transfection was performed with Lipofectamine® 2000 (Invitrogen, Shanghai, China) in accordance with the manufacturer's instructions.
Quantitative real time polymerase chain reaction
The circ_0000388 or miR-337-3p expression was detected by quantitative real-time polymerase chain reaction (PCR). TRIzol lysate (Invitrogen) was used to suspend and lyse the cells. Afterward, the RNA in the cell lysis were extracted by chloroform-isoamyl alcohol method. Spectrophotometry was used to detect the purity and concentration of RNA samples. In accordance with the instructions, 2 μg RNA is reversely transcribed into cDNA with PrimeScript RT Reagent Kit (Invitrogen). Then, 1 μL cDNA sample was taken to prepare 50 μL quantitative PCR reaction system containing SYBR Green Premix Ex Taq II (Takara, Dalian, China). Melting curves were plotted for evaluating the specificity of PCR amplification. The internal reference genes were GAPDH or U6, and the relative expression was calculated by using 2−ΔΔCT method. The primers used in PCR of this study were shown in Table 1.
Primer Sequences Used in Polymerase Chain Reaction for This Study
GAPDH, glyceraldehyde 3-phophate dehydrogenase; TCF12, transcription factor 12.
CCK-8
Cell viability was assessed using the CCK-8 assay. Cells were seeded in 96-well plates at a density of 1 × 103 cells/well for 24 h, and 10 μL of CCK-8 reagent (Dojindo Molecular Technologies, Kumamoto, Japan) was added to each well in accordance with the manufacturer's instructions. The value of optical density (OD) at 450 nm was measured on a spectrophotometer (Bio-Rad, CA) after incubation for 1 h in a 37°C incubator. The same procedures were conducted on 48, 72, and 96 h, respectively. Finally, the proliferation curve was plotted according to the OD value.
Cell scratch assay
First, cells were inoculated in a six-well plate supplemented with 2 mL of complete medium to each well, and when the cells covered the bottom of the wells, the scratch was made with a vertical tip and then the cells were washed twice with phosphate-buffered saline (PBS). Subsequently, the complete medium was replaced by serum-free medium. Then, the cells were observed under the inverted microscope and recorded as 0 h. Afterward, the culture was continued at 37°C in 5% CO2, and the plate was taken out after 24 h, and the scratch healing was observed at the same observation point.
Transwell assay
Cell migration and invasion were assessed by transwell assay. The Transwell chamber (Corning, NY) was covered with 200 mg/mL Matrigel (BD, San Jose) and incubated overnight. Cells were then added to the upper chamber with serum-free medium. The medium (500 μL) containing 10% FBS was added in the lower chamber as chemotactic agent. After 24 h of culture, all noninvasive cells were cleared. The Matrigel membrane was removed and fixed with paraformaldehyde and then stained with a crystal violet. At last, the number of cells was counted using inverted microscope (Olympus, Tokyo, Japan). In the migration assay, Matrigel was not used and the other procedures were same as the invasion assay.
Western blot
The cells were harvested and incubated with precooled RIPA lysate for 20 min. After centrifugation, the supernatant was taken, and the protein was quantified by BCA protein quantification kit, and the protein concentration was adjusted and boiled to denature for 5 min. Total proteins were separated by SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) and transferred to polyvinylidene fluoride membrane. Then the membrane was blocked at room temperature with 3% bovine serum albumin in TBST solution. After that, the diluted TCF12 antibody (rabbit anti-human polyclonal antibody, 1:1000, ab245540; Abcam) was added and maintained at 4°C overnight. Then the membrane was washed three times with TBST for 3 min each time. Subsequently, diluted secondary antibody (goat anti-rabbit IGg, 1:2000, ab205718; Abcam) was added, incubated at room temperature for 40 min, and then the membrane was washed by TBST again. At last, the membrane was added with chemiluminiscence using hypersensitive ECL (Guangzhou Xiangbo Biotechnology Co., Ltd.) to develop the bands. Glyceraldehyde 3-phophate dehydrogenase (GAPDH) was used as internal reference. The GAPDH primary antibody was anti-GAPDH (murine anti-human monoclonal antibody, 1:1000, ab8245; Abcam), the secondary antibody was goat anti-mouse IgG (1:2000, ab205719; Abcam).
TUNEL assay
The medium was removed and the cells were washed once with PBS. The cells were fixed with immunostaining fixative for 30 min, and washed once with PBS. The immunostaining washing solution was added and the cells were incubated for 2 min in an ice bath. Subsequently, 50 μL of TUNEL assay solution (Invitrogen) was added to the sample and the sample was incubated at 37°C for 60 min in the dark. The sample was washed three times with PBS. After being sealed with an anti-fluorescence quenching liquid, the fluorescence was observed under a fluorescence microscope, and the excitation light was 450 to 500 nm, and the emission light was 515 to 565 nm (green fluorescence). A random field of five fields was selected for each sample to calculate the apoptotic rate: apoptotic rate = apoptotic cells/total cells × 100%.
Dual-luciferase reporter gene assay
All luciferase reporter vectors (circ_0000388-WT, circ_0000388-MUT, TCF12-WT, TCF12-MUT) were constructed by Promega (Madison, WI). First, HeLa and SiHa cells (4.5 × 104) were seeded in 48-well plates and cultured to 70% confluence. The above luciferase reporter vector, miR-337-3p mimics or negative control was then co-transfected into HeLa and SiHa cells using Lipofectamine 2000 (Invitrogen). After 48 h of transfection, luciferase activity was determined in accordance with the manufacturer's instructions.
RNA immunoprecipitation assay
The RNA immunoprecipitation (RIP) assay was performed using the Magna RIP RNA Binding Protein Immunoprecipitation Kit (Millipore). Approximately 1 × 107 cells were resuspended in RIP lysis buffer plus protease and RNase inhibitor (Epicentre). Subsequently, 100 mL of cell lysate was incubated with 5 mg of control IgG or AGO2 antibody-coated beads at 4°C overnight, respectively. After treatment with proteinase K, the immunoprecipitated RNA was extracted by RNeasy Min Elute Cleanup Kit (Qiagen, German) and subjected to reverse transcription. The abundance of circ_0000388 was then detected by quantitative real time polymerase chain reaction (qRT-PCR).
Statistical analysis
All data in this study were processed using SPSS 20.0 statistical analysis software (SPSS, Inc., Chicago, IL); measurement data were expressed as mean ± standard deviation (x ± s), and Student's t-test was used for comparison analysis between groups. The count data or percentage (%) were compared using chi-square analysis; p < 0.05 was considered statistically significant.
Results
Circ_0000388 was related to CC process
To preliminarily determine the role of circ_0000388 in CC, we first detected the circ_0000388 expression in CC tissues and normal cervical tissues by qRT-PCR, and found that in comparison with normal tissues, circ_0000388 expression in CC tissues was significantly elevated (Fig. 1A). We further analyzed the circ_0000388 expression and the pathological features of patients. The findings implied that the circ_0000388 expression was notably correlated with the FIGO stage, lymph node metastasis, and depth of invasion (Table 2). Additionally, a significant increase in the circ_0000388 expression was also observed in human CC cell lines in comparison with normal human cervical epithelial cells (Fig. 1B). The above results implied that there was a correlation between circ_0000388 and the tumorigenesis of CC.
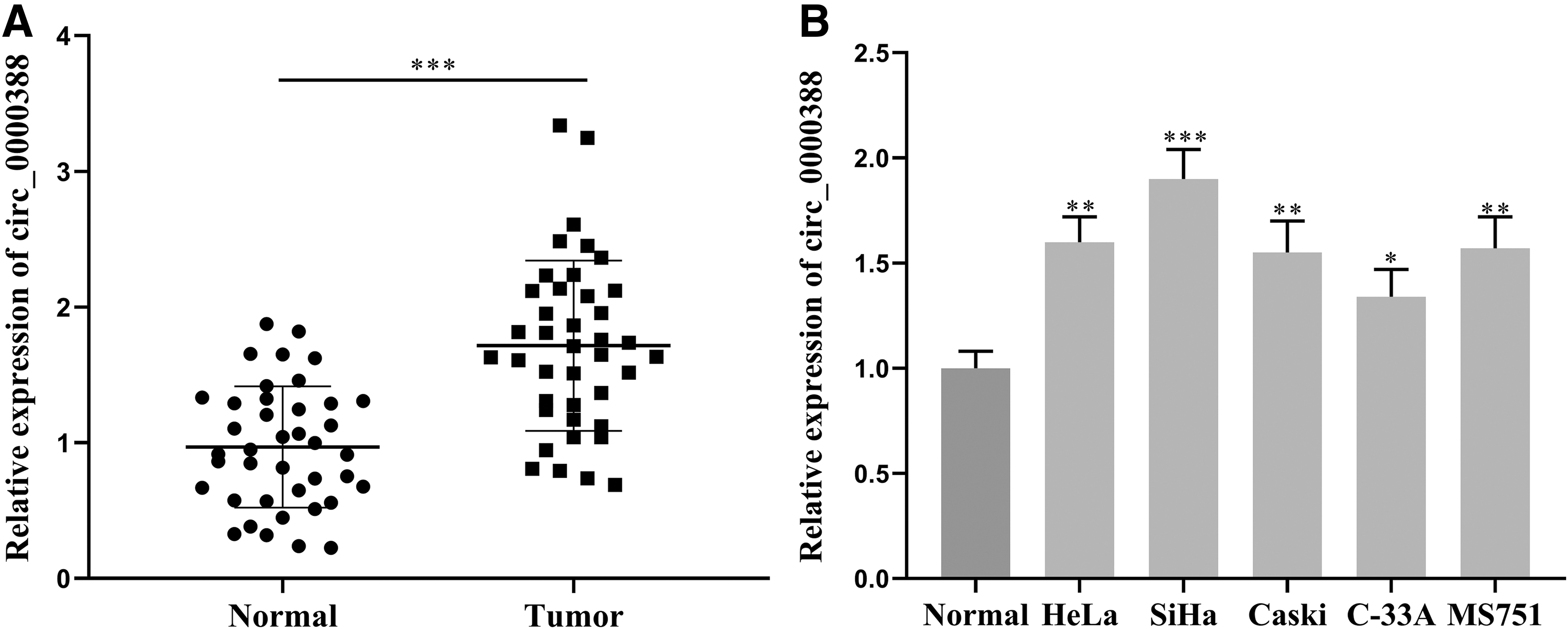
Circ_0000388 expression in CC tissues and cell lines.
Relationship Between Characteristics of Cervical Cancer Patients and circ_0000388 Expression
The effect of circ_0000388 on CC cells
To further probe the function of circ_0000388, we successfully constructed circ_0000388 overexpressing and knockdown cell lines in HeLa and SiHa cell lines, respectively (Fig. 2A). Through CCK-8 assay, we found that circ_0000388 overexpression remarkably enhanced the proliferation of HeLa cells, and knockdown of circ_0000388 dramatically repressed the proliferation of SiHa cells (Fig. 2B, C). Cell scratch assay and Transwell assay were employed to detect cell migration and invasion, and the results reflected that circ_0000388 overexpression notably enhanced the migration and invasion of CC cells, and knocking down circ_0000388 gave the opposite result (Fig. 2D, E). Through TUNEL assay, we demonstrated that circ_0000388 upregulation markedly suppressed the apoptosis in HeLa cells, and knockdown of circ_0000388 remarkably increased the apoptosis rate of SiHa cells (Fig. 2F). Collectively, we concluded circ_0000388 had a significant effect on the proliferation, migration, invasion, and apoptosis of CC cells.

Effect of circ_0000388 on CC cells.
MiR-337-3p was low-expressed in cervical CC tissues and cell lines
To preliminarily clarify the role of miR-337-3p in CC, we also detected the miR-337-3p expression in CC tissues and cell lines by qRT-PCR. In comparison with normal tissues, the miR-337-3p expression was dramatically decreased in CC tissues (Fig. 3A), and miR-337-3p expression in CC cell lines in comparison with normal human CC epithelial cells was notably decreased (Fig. 3B). Additionally, further studies found a negative correlation between the miR-337-3p expression and circ_0000388 expression in CC tissues (r 2 = 0.3641, Fig. 3C). In conclusion, miR-337-3p may be associated with CC development and linked to circ_0000388.

Expression of miR-337-3p in CC tissues and cell lines.
The effect of miR-337-3p on CC cells
To further probe the role of miR-337-3p in the regulation of the biological function of CC cells, we successfully constructed miR-337-3p overexpression and inhibition models in HeLa and SiHa cell lines, respectively (Fig. 4A). Cell proliferation was detected by CCK-8 assay, and miR-337-3p mimics and inhibitors remarkably impeded and facilitated the proliferation of CC cells in comparison with the control group, respectively (Fig. 4B, C). In scratch healing assay and Transwell assay, miR-337-3p mimics significantly repressed the migration and invasion of HeLa cells, and miR-337-3p inhibitors dramatically enhanced the migration and invasion of SiHa cells (Fig. 4D, E). Furthermore, the results of the TUNEL assay implied that miR-337-3p mimics enhanced HeLa cell apoptosis, while miR-337-3p had an opposite effect on SiHa cells (Fig. 4F). In summary, miR-337-3p significantly repressed proliferation, migration and invasion and promoted apoptosis of CC cells.

Effect of miR-337-3p on CC cells.
Interaction between circ_0000388 and miR-337-3p
Accumulating researches indicate that circRNA could exert its biological functions via targeting miRNAs. 19 Since we had found the opposite characteristics of circ_0000388 and miR-337-3p in expression and function, we were also interested in whether the two can interact with each other. Therefore, we detected the downstream target of circ_0000388 through bioinformatics analysis and found that there was a binding site between it and miR-337-3p (Fig. 5A). The dual-luciferase reporter gene assay demonstrated that miR-337-3p significantly reduced circ_0000388 wild-type luciferase activity, but had no significant effect on luciferase activity of the mutant circ_0000388 (Fig. 5B). RIP assay also implied that miR-337-3p were enriched in Ago2 immunoprecipitation compared to IgG immunoprecipitation (Fig. 5C), which confirmed the direct binding relationship between circ_0000388 and miR-337-3p. We also examined the effect of circ_0000388 on the miR-337-3p expression by qRT-PCR, and found that overexpression and knockdown of circ_0000388 expression remarkably suppressed and promoted miR-337-3p in CC cells respectively (Fig. 5D). Hence, circ_0000388 could targetedly inhibit miR-337-3p expression.
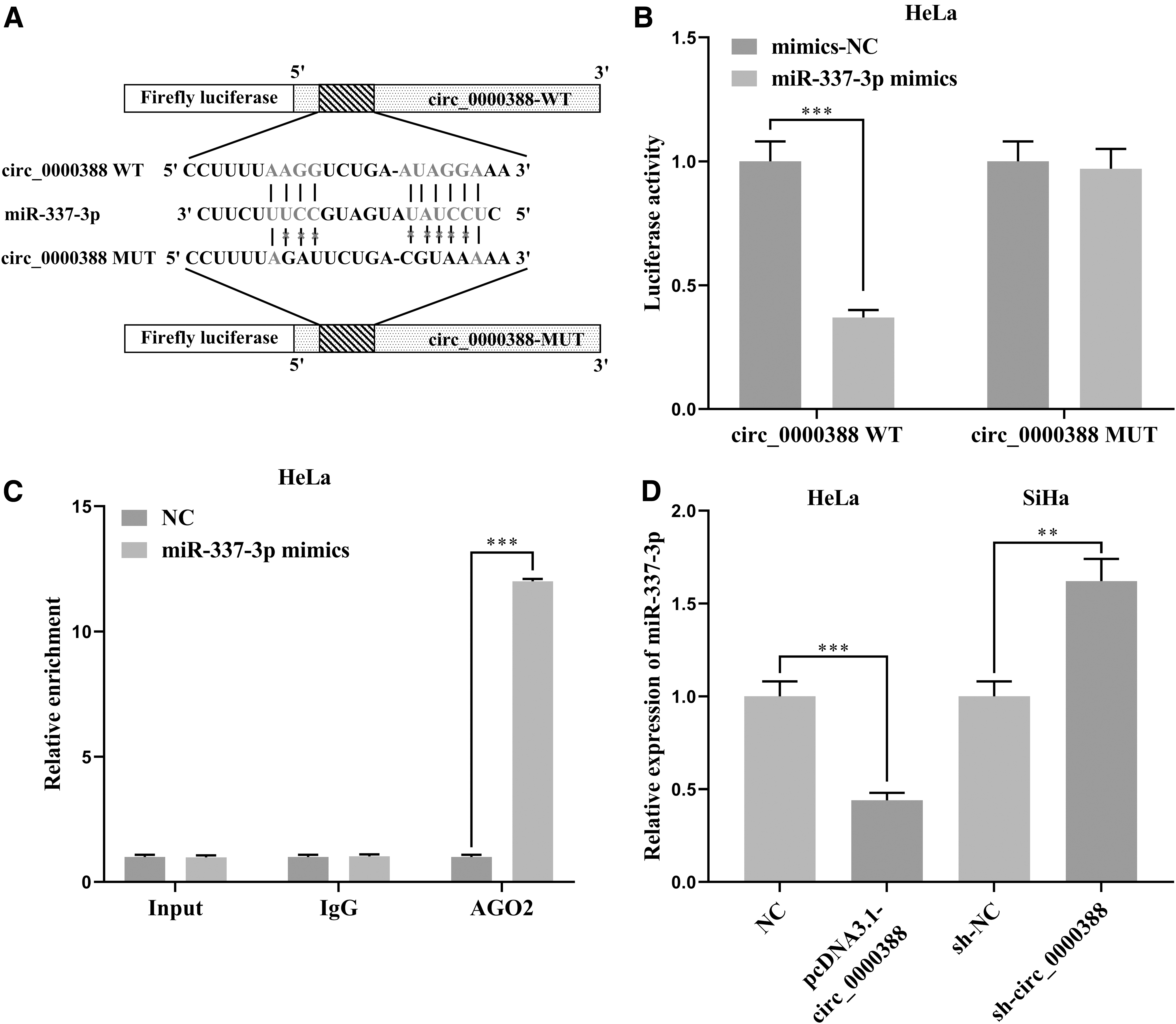
Interaction between circ_0000388 and miR-337-3p.
MiR-337-3p reversed the cancer-promoting effect of circ_0000388
To further verify the interaction between circ_0000388 and miR-337-3p, we transfected miR-337-3p mimics into HeLa cells with overexpressed circ_0000388. The CCK-8 assay was performed again and it was found that miR-337-3p mimics attenuated the effect of circ_0000388 on facilitating HeLa cell proliferation ( Fig. 6A). The results of the scratch assay and the Transwell assay indicated that the migration and invasion of HeLa cells were remarkably impeded by miR-337-3p in comparison with the circ_0000388 overexpression (Fig. 6B, C). In the TUNEL assay, the apoptotic rate of the pcDNA3.1-circ_0000388+miR-337-3p mimics group was markedly higher than that of the pcDNA3.1-circ_0000388 group (Fig. 6D). These results further confirmed the interaction between circ_0000388 and miR-337-3p.
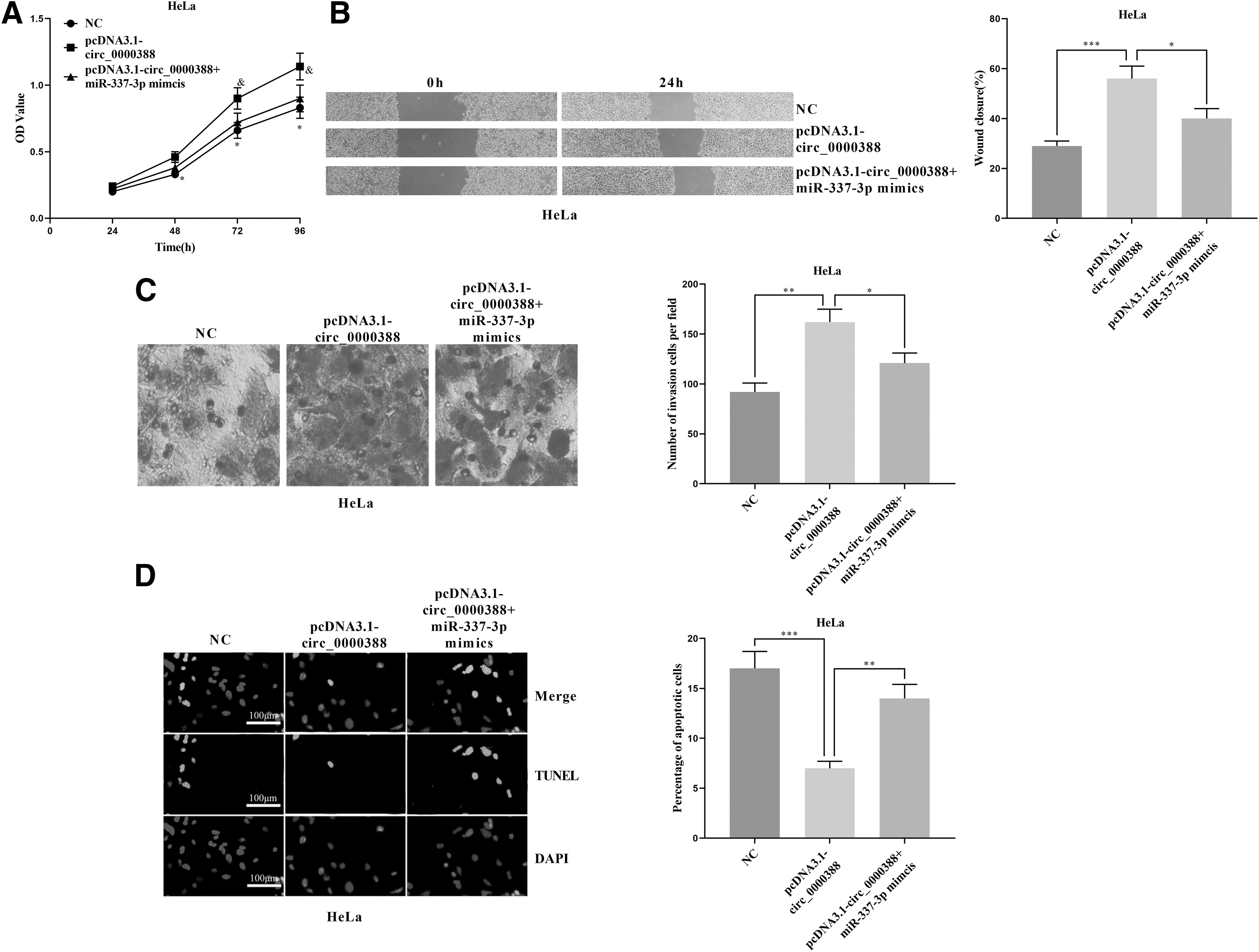
MiR-337-3p reversed the effect of circ_0000388 on CC cells.
Circ_0000388 indirectly modulated TCF12 through miR-337-3p
To further probe the downstream mechanism of miR-337-3p, we predicted the downstream target of miR-337-3p through TargetScan and found that there were 3 binding sites between miR-337-3p and the 3′UTR of TCF12 (Fig. 7A). Dual-luciferase reporter gene assay validated that all of these three binding sites were effective in CC cells (Fig. 7B, C). Furthermore, miR-337-3p mimics notably impeded the TCF12 mRNA expression and its protein, whereas miR-337-3p inhibitors had the opposite effect (Fig. 7D, E). In HeLa cells with circ_0000388 overexpression, we observed a significant increase in TCF12 mRNA and its protein, whereas miR-337-3p mimics reversed this promotion (Fig. 7F, G). The above results reflected that TCF12 could be targeted and repressed by miR-337-3p and indirectly modulated by circ_0000388.

Circ_0000388 indirectly regulated TCF12 by miR-337-3p.
Discussion
In recent years, the biological functions of a variety of circRNAs in cancer have been gradually studied. These abnormally expressed circRNAs not only modulate the development of various tumors but also can serve as the indicators of the prognosis. 20 –25 For instance, circ_0081001 expression in osteosarcoma is significantly increased, and the highly expressed circ_0081001 is thought to indicate poor prognosis 23 ; circ_0039411 has been validated to be highly expressed in papillary thyroid carcinoma, and it enhances the proliferation, migration, and invasion of cancer cells 24 ; the circ_0002138 expression in colorectal cancer tissues is significantly decreased and it markedly impedes the proliferation of cancer cells. 25 In the study, we explored the expression characteristics and biological functions of circ_0000388. We found that circ_0000388 was remarkably increased in CC tissues and cell lines and was closely related to CC progression. We further investigated the role of circ_0000388 in CC through CCK-8, cell scratch, transwell and TUNEL assays, and found that overexpressed circ_0000388 remarkably enhanced the proliferation, migration, and invasion of CC cells, whereas inhibiting their apoptosis; conversely, the opposite result was observed when circ_0000388 was knocked down. The above results implied that circ_0000388 had a significant effect on promoting CC progression.
Besides, we identified miR-337-3p as a tumor suppressor in CC. We demonstrated that miR-337-3p mimics remarkably restrained the proliferation, migration, and invasion of CC cells. The opposite result was observed when the miR-337-3p inhibitors were transfected into CC cells, which was contrary to the effect of circ_0000388. CircRNA can also serve as endogenous competitive RNA to sponge specific miRNA, thereby repressing the biological function of the latter. This interaction has become one of the hotpots of recent oncology research. 3 For instance, in CC, circ_0023404 can sponge miR-136 to regulate TFCP2/YAP aixs to exert its cancer-promoting effect. 26 In our study, miR-337-3p was found to have a binding site between circ_0000388 and miR-337-3p. The overexpressing circ_0000388 significantly suppressed the miR-337-3p expression, and miR-337-3p expression was notably increased after knocking down circ_0000388. In other previous studies, miR-337-3p has also been reported to be modulated by circRNA. For instance, in chronic lymphocytic leukemia, miR-337-3p is targeted and repressed by circ_0132266. 27 The regulatory mechanism of miR-337-3p found in the study was similar to that.
TCF12 has been confirmed to modulate biological processes including immune response, cancer metastasis, cognitive function, and so on. 28 –31 It can reduce the E-cadherin expression via repressing inhibitor of differentiation-1 to accelerate cancer progression. 32 Additionally, the inhibition of TCF12 can restrain the activation of the PI3K/AKT signaling pathway. 15 The PI3K/Akt pathway is a crucial signaling pathway involved in cell proliferation and epithelial–mesenchymal transition during tumorigenesis, and in CC, this pathway has been validated to have a cancer-promoting effect. 33 In the study, TCF12 was verified as a downstream target of miR-337-3p, and negatively regulated by it, but positively regulated by circ_0000388. Our study proved that circ_0000388 was a sponge for miR-337-3p, and promoted the expression of TCF12, which partly clarify the dysregulation of TCF12 in CC.
Taken together, we demonstrate that circ_0000388 can enhance the proliferation, migration, and invasion and repress the apoptosis of CC cells via regulating miR-377-3p/TCF12 axis. Even though research on circRNA in cancer is still in its infancy, and its clinical application has a long way to go, our research reveals a novel mechanisms in the CC progression, which provides implications for the development of new and effective therapeutics. In the future, in vivo studies are required to further validate our demonstrations. Additionally, our study demonstrates the correlation between circ_0000388 and the FIGO stage of CC, which implies that circ_0000388 can probably be a diagnosis and prognosis marker of CC. In the following work, clinical samples should be collected from multiple medical centers to validate this potential.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Authors' Contributions
Conceived and designed the experiments: X.-j.J., Q.-h.M., and Y.L.; Performed the experiments: Q.-h.M., Y.L., and C.K.; Statistic analysis: Q.-h.M., Y.L., and X.M.G.; Wrote the article: Q.-h.M. and Y.L. All authors read and approved the final article.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.