
Abstract
Background:
Hepatocyte nuclear factor 1 homeobox A-antisense RNA 1 (HNF1A-AS1) is a long noncoding RNA and controls human tumor development and progression. However, its expression and role in breast cancer, the most overwhelmingly occurring malignancy in women globally, remain poorly illuminated.
Materials and Methods:
Expression of HNF1A-AS1, miRNA (miR)-20a-5p, and tripartite motif containing 32 (TRIM32) was detected using quantitative real-time polymerase chain reaction and Western blotting. Cell proliferation, apoptosis, migration, and invasion were measured by cellTiter 96 AQueous one solution cell proliferation assay kit, flow cytometry, and transwell assays, respectively. Epithelial-mesenchymal transition (EMT) was evaluated by Western blotting, analyzing E-cadherin, N-cadherin, and vimentin expression. Mice xenograft model was generated to investigate tumor growth in vivo. The target binding among miR-20a-5p, HNF1A-AS1, and TRIM32 was confirmed by dual-luciferase reporter assay.
Results:
Expression of HNF1A-AS1 and TRIM32 was upregulated and miR-20a-5p was downregulated in breast cancer tumors and cell lines. Deletion of HNF1A-AS1 induced cell apoptosis rate, but suppressed cell proliferation, EMT, migration, and invasion in MDA-MB-231 and MCF-7 cells. Furthermore, HNF1A-AS1 downregulation impeded tumor growth in vivo. Interestingly, miR-20a-5p overexpression elicited the similar suppressive effects in MDA-MB-231 and MCF-7 cells, which was partially reversed by TRIM32 upregulation; besides, miR-20a-5p silencing could abolish the antitumor role of HNF1A-AS1 deletion. Notably, HNF1A-AS1 positively modulated TRIM32 expression through acting as a molecular “sponge” for miR-20a-5p.
Conclusions:
Knockdown of HNF1A-AS1 suppressed breast carcinogenesis presumably through targeting miR-20a-5p/TRIM32 axis, suggesting that HNF1A-AS1 might be a promising therapy target for breast cancer.
Introduction
Breast cancer is considered the leading cause of new cancer diagnoses in women worldwide. 1 According to gene expression profiling, breast cancer is classified into four clinically relevant subtypes: luminal like, human epidermal growth factor receptor 2 (HER2) enriched, basal like, and normal like. 2 Up to now, radio therapy and chemotherapy are widely used as the first-line treatment in breast cancer 3 ; however, acquired resistance and metastasis are the main undesired consequences after these approaches. Among the emerging therapies in cancer, gene therapy has been caught much attention. 4,5 Therefore, it is urgent and imperative to elucidate novel molecular mechanisms correlated with breast cancer development.
MicroRNAs (miRNAs) are a class of endogenous noncoding transcripts and consisted of approximately 22 nucleotides. Numerous evidence has proposed the dysregulation of miRNAs as a hallmark for cancer initiation and metastasis. 6 Many cancer-related miRNAs have been identified in the biology of breast cancer, and circulating miRNAs probably are promising diagnostic and therapeutic targets for breast cancer. 7,8 miRNA-20a-5p (miR-20a-5p) is a crucial molecule in many biological processes in cancer progression. In breast cancer, it has been announced to be involved in autophagy, angiogenesis, malignant behaviors, and chemotherapeutic and radiosensitive response. 9 –12 However, the molecular mechanism of miR-20a-5p has not been completely expounded upon yet.
The miRNA-long non-coding RNA (lncRNA) interaction is a pivotal pattern for the adjustment of miRNA-related gene expression. lncRNAs are a type of endogenous transcripts with longer than 200 nucleotides and act as a competing endogenous RNA (ceRNA) to sponge target miRNAs. Previous studies have confirmed that several lncRNAs regulate miR-20a-5p concentration in human cancers, 13 –15 including breast cancer. 16 Hepatocyte nuclear factor 1 homeobox A-antisense RNA 1 (HNF1A-AS1) is a novel lncRNA. 17 In human cancers, HNF1A-AS1 participates in controlling tumor development and progression. 18 Nevertheless, the expression and role of this lncRNA in breast cancer are poorly clarified so far. Besides, there is no reported evidence to define the interaction between HNF1A-AS1 and miR-20a-5p as well.
Tripartite motif containing 32 (TRIM32) is a member of TRIM family and possesses E3 Ubiquitin ligase activity, which then contributes to tumorigenesis. 19 The biological role of TRIM32 remains to be well annotated. In this study, the authors measured expression of HNF1A-AS1, miR-20a-5p, and TRIM32 in breast cancer tissues and cell lines. The effects of HNF1A-AS1 deletion and miR-20a-5p overexpression were further investigated in malignant phenotypes of breast cancer cells in vitro and in vivo. Notably, the interplay among the three was identified.
Materials and Methods
Clinical samples
The study was conducted with the approval of the Ethical and Scientific Committees of Xi'an NO.3 Hospital and with informed consent signed by every participator. A total of 30 patients (n = 30; average age, 46 years; tumor stage I/II, n = 21 and III/IV n = 9, Supplementary Table S1) with primary breast cancer were recruited in Xi'an NO.3 Hospital. Then, breast cancer tissues and the paired adjacent normal tissues were obtained from this cohort of patients during surgery. The normal tissues were acquired at least 5 cm away from breast cancer tissues. The excluded patients received antitumor therapies before, such as surgery, radiotherapy, and chemotherapy. All the tissue samples were snap-frozen in liquid nitrogen for further detection.
Cell culture
Human breast cancer cell lines MDA-MB231 (HTB-26) and MCF-7 (HTB-22) were purchased from American Type Culture Collection (Manassas, VA), as well as a normal breast cell line MCF-10A (CRL-10137). All cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Hyclone, Logan, UT) containing 10% fetal bovine serum (FBS; Hyclone). The culture methods were consistent with the suggestion of ATCC.
Quantitative real-time polymerase chain reaction
The frozen tissues and cultivated cells were lysed in E.Z.N.A. Total RNA Kit I (OMEGA, Norcross, Georgia) and the total RNA was extracted following the standard procedures. High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) was used to synthesize cDNA and RevertAid First Strand cDNA Synthesis kit (Thermo Scientific, Pittsburgh, PA) was utilized to amplify gene expression with special primers. The semiquantitation was performed with 2−ΔΔCT method on Light Cycler 96 (Roche, Basel, Switzerland). Glyceraldehyde-phosphate dehydrogenase (GAPDH) and U6 small nuclear RNA (U6) acted as the endogenous controls.
The primers were listed as follows: HNF1A-AS1, 5′-TCAAGAAATGGTGGCTAT-3′ (sense) and 5′-GCTCTGAGACTGGCTGAA-3′ (anti-sense) 17 ; miR-20a-5p, 5′-CCCCTAAAGTGCTTATAGTGC-3′ (sense) and 5′-GTGCAGGGTCCGAGGT-3′ (anti-sense); TRIM32, 5′-GGCCACTGTACACTCCCTGT-3′ (sense) and 5′-AGCCAAAGAGCCTGTGAAGA-3′ (anti-sense); GAPDH, 5′-GGGAGCCAAAAGGGTCAT-3′ (sense) and 5′-GAGTCCTTCCACGATACCAA-3′ (anti-sense); and U6, 5′-GCGCGTCGTGAAGCGTTC-3′ (sense) and 5′-GTGCAGGGTCCGAGGT-3′ (anti-sense).
Cell transfection
Oligonucleotides small interfere RNA (siRNA) against HNF1A-AS1 (si-HNF1A-AS1), miR-20a-5p mimics and miR-20a-5p inhibitors (anti-miR-20a-5p) were synthesized by GENEWIZ (Suzhou, China), as well as their negative controls si-NC, miR-NC mimic and anti-miR-NC. (Invitrogen, Carlsbad, CA) was employed to amplify the full length of HNF1A-AS1 (NR_024345) and coding domain sequence of TRIM32. Briefly, 50 nM of oligonucleotides or 2 μg vectors was transfected into MDA-MB231 and MCF-7 cells (about 60% confluence) by Lipofectamine 2000 (Invitrogen). In rescue experiments and dual luciferase reporter assay, half nucleotides were co-transfected in cells. The special sequence of siRNAs were as follows: si (sh)-HNF1A-AS1, 5′-GGGUGAGCAGCUGUUUGCAAGACUA-3′ 20 ; and si (sh)-NC, 5′-CACCTGCATTCAAACTCGGACTGTT-3′. Every transfection group was performed three times for further analysis.
Cell proliferation assay
A cellTiter 96 AQueous one solution cell proliferation assay kit (MTS; Promega, Madison, MI) was used to detect cell proliferation. Transfected cells were reseeded in a 96-well plate (Corning, Inc., Corning, NY) at a density of 4000 cells/well. After transfection for 0, 24, 48, and 72 h, 20 μL of reagent was added into each well and incubated for another 2 h. Absorbance at wavelength of 490 nm was determined using a microplate reader (Molecular Devices, Shanghai, China) and cell growth curve was drawn according to the optical density values. The reactions were performed in sextuplicate for each group.
Flow cytometry
After transfection for 24 h, cells were collected to evaluate cell apoptosis by Annexin V-FITC Apoptosis Detection kit (Sigma-Aldrich, St. Louis, MO). The cells were resuspended in 500 μL of binding buffer, treated with 5 μL Annexin V-FITC and 5 μL propidium iodide (PI) for 30 min in the dark, and then analyzed on FACSCalibur flow cytometer (BD Biosciences, Franklin Lakes, NJ). The data analysis was performed on CellQuest program (BD Biosciences) and apoptosis rate was recorded as percentage of apoptotic cells in Annexin V+/PI− and Annexin V+/PI+ quadrants. Apoptosis rate in each group was analyzed on flow cytometry for three times in three independent experiments.
Transwell assays
The abilities of cell migration and invasion of MDA-MB-231 and MCF-7 cells were measured by transwell assays after transfection for 24 h. For invasion analysis, transwell chamber (8 μm pore size; Corning, Inc.) coated with matrigel (BD Biosciences) was loaded in a 24-well plate (Corning, Inc.). The lower chamber was 500 μL of DMEM with 10% FBS (Hyclone) and the upper chamber was 200 μL of cell suspension (1 × 105) in DMEM without FBS (Hyclone). The plates were incubated for 24 h and the cells invading the bottom chamber were stained with crystal violet and being photographed under a microscope in five predetermined fields ( × 200). Three independent experiments were carried out. The migration analysis was depending on transwell chamber (Corning, Inc.) without matrigel, and the other operations were the same as invasion analysis.
Western blotting
Total protein was harvested from transfected MDA-MB-231 and MCF-7 cells at 24 h using RIPA lysis buffer (Thermo Scientific). The extraction (20 μg) was loaded for the standard procedures of Western blotting assay. The primary antibodies, including TRIM32 (#131223; 1:5000), E-cadherin (#231303; 1:1000), N-cadherin (#98952; 1:1000) and vimentin (#137321; 1:2000), were purchased from Abcam (Cambridge, UK), and GAPDH (#181602; 1:5000) was the loading control. The protein bands were detected using an enhanced chemiluminescence system. Western blotting was performed in triplicate for every sample and the relative protein expression was semiquantified on Image J.
Dual-luciferase reporter assay
The wild type (WT) and mutant (MUT) of HNF1A-AS1 and TRIM32 3′ untranslated region (3′ UTR) containing the complementary binding site of miR-20a-5p were separately inserted into pGL4 (Promega) dual luciferase vector. MDA-MB-231 and MCF-7 cells were co-transfected with miR-20a-5p mimics or miR-NC mimics with either HNF1A-AS1/MTDH-WT or HNF1A-AS/MTDH-MUT. After 24-h transfection, the firefly luciferase activity and Renilla luciferase activity (served as the internal control) were performed using GloMax LUMINOMETER (Promega). All operations were carried out at least three times.
Xenograft model
MCF-7 and MDA-MB-231 cells were infected by lentiviral particles encoding shRNA against HNF1A-AS1 (sh-HNF1A-AS1; Neuron Biotech, Shanghai, China) or the negative control (sh-NC) using Polybrene reagent (Sigma-Aldrich). A total of 24 nude mice (male; 6-week-old BALB/c) were obtained from Model Animal Research Center of Nanjing University. Then, the stably transfected MCF-7 cells (3 × 106) and MDA-MB-231 cells (3 × 106) were subcutaneously injected into right flanks (n = 6) of nude mice, respectively. The xenograft tumors were examined and measured every week. The longest tumor axis (L) and the shortest tumor axis (S) were evaluated by a caliper and the volume of tumor was determined by the following formula: 0.5 × LS 2 . The tumor weight was evaluated with an electronic balance. Finally, the tumors were isolated in the fifth week after the mice were euthanatized.
The animal experiment was in accordance of the rules of the Ethics Committee of Xi'an NO.3 Hospital. The feeding and operations were carried out in accordance with National Institutions of Health Guide for Care and Use of Laboratory Animals.
Statistical analysis
The data were presented as mean ± standard deviation and the statistical analysis was performed on GraphPad Prism 5 (GraphPad, San Diego, CA). Two-tailed Student's t-test and one-way analysis of variance were performed for comparisons between groups. The significant difference was acknowledged as p-value <0.05. Pearson correlation analysis was used to estimate the correlation between the expression levels of HNF1A-AS1, miR-20a-5p, and TRIM32.
Results
HNF1A-AS1 was highly expressed in breast cancer tissues and cell lines
To investigate whether HNF1A-AS1 is deregulated in breast cancer, the authors recruited a cohort of patients (n = 30) with breast cancer, and the clinicopathological features are presented in Supplementary Table S1. According to quantitative real-time polymerase chain reaction (qRT-PCR) analysis, the relative expression level of HNF1A-AS1 was dominantly higher in tumor samples than that in the paired adjacent normal tissues (Fig. 1A). Besides, human breast cancer cell lines MDA-MB-231 and MCF-7 exhibited higher level of HNF1A-AS1 comparing to normal breast cell line MCF-10A (Fig. 1B). These findings showed a dysregulation of HNF1A-AS1 in breast cancer, suggesting a potential oncogenic role of HNF1A-AS1.
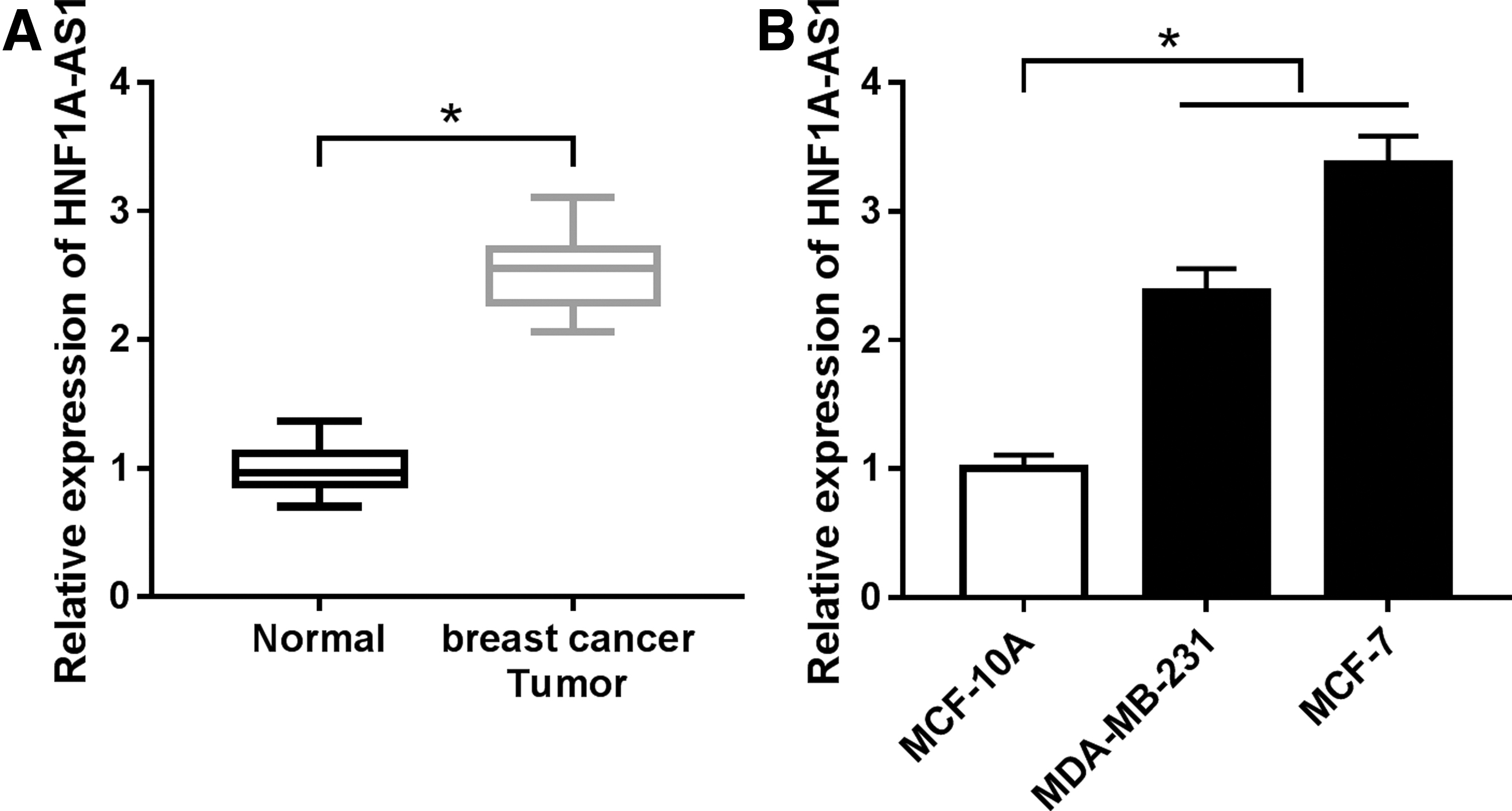
lncRNA HNF1A-AS1 (HNF1A-AS1) was highly expressed in breast cancer tissues and cell lines. qRT-PCR analysis was carried out to test relative expression of HNF1A-AS1 in
Deletion of HNF1A-AS1 suppressed the aggressiveness of breast cancer cells in vitro
Based on the observation above, the authors performed a series of loss-of-function experiments. MDA-MB-231 and MCF-7 cells were transfected with si-HNF1A-AS1 or si-NC. qRT-PCR showed a dramatic inhibition of HNF1A-AS1 level after si-HNF1A-AS1 transfection (Fig. 2A, B), suggesting an effective transfection efficiency in both MDA-MB-231 and MCF-7 cells. With HNF1A-AS1 deletion, cell proliferation was significantly decreased at 72 h as described by MTS assay (Fig. 2C, D), while flow cytometry (FCM) analysis presented a promotion on apoptosis of MDA-MB-231 and MCF-7 cells after transfection with si-HNF1A-AS1 for 24 h (Fig. 2E, F).

Deletion of HNF1A-AS1 suppressed the aggressiveness of breast cancer cells in vitro. MDA-MB-231 and MCF-7 cells were transfected with siRNA against HNF1A-AS1 (si-HNF1A-AS1) or its negative control (si-NC).
In terms of metastatic ability, transwell assays depicted an obvious loss of migrated cells and invaded cells in HNF1A-AS1-silenced MDA-MB-231 and MCF-7 cells at 24 h (Fig. 2G, H). Furthermore, expression of epithelial-mesenchymal transition (EMT)-related markers was identified. As Figure 2I and J showed, E-cadherin expression was highly induced, whereas N-cadherin and vimentin expression were distinctively descended by 24 h transfection of si-HNF1A-AS1. These results showed that deletion of HNF1A-AS1 could suppress the malignant phenotypes of MDA-MB-231 and MCF-7 cells, indicating HNF1A-AS1 knockdown might result in impairment of breast cancer cell aggressiveness.
HNF1A-AS1 physically targeted miR-20a-5p
To further investigate the molecular mechanism of HNF1A-AS1 in breast carcinogenesis, bioinformatics tools were utilized to predict the potential pairing bases between HNF1A-AS1 and miRNAs. DIANA tool LncBase Predicted v.2 (
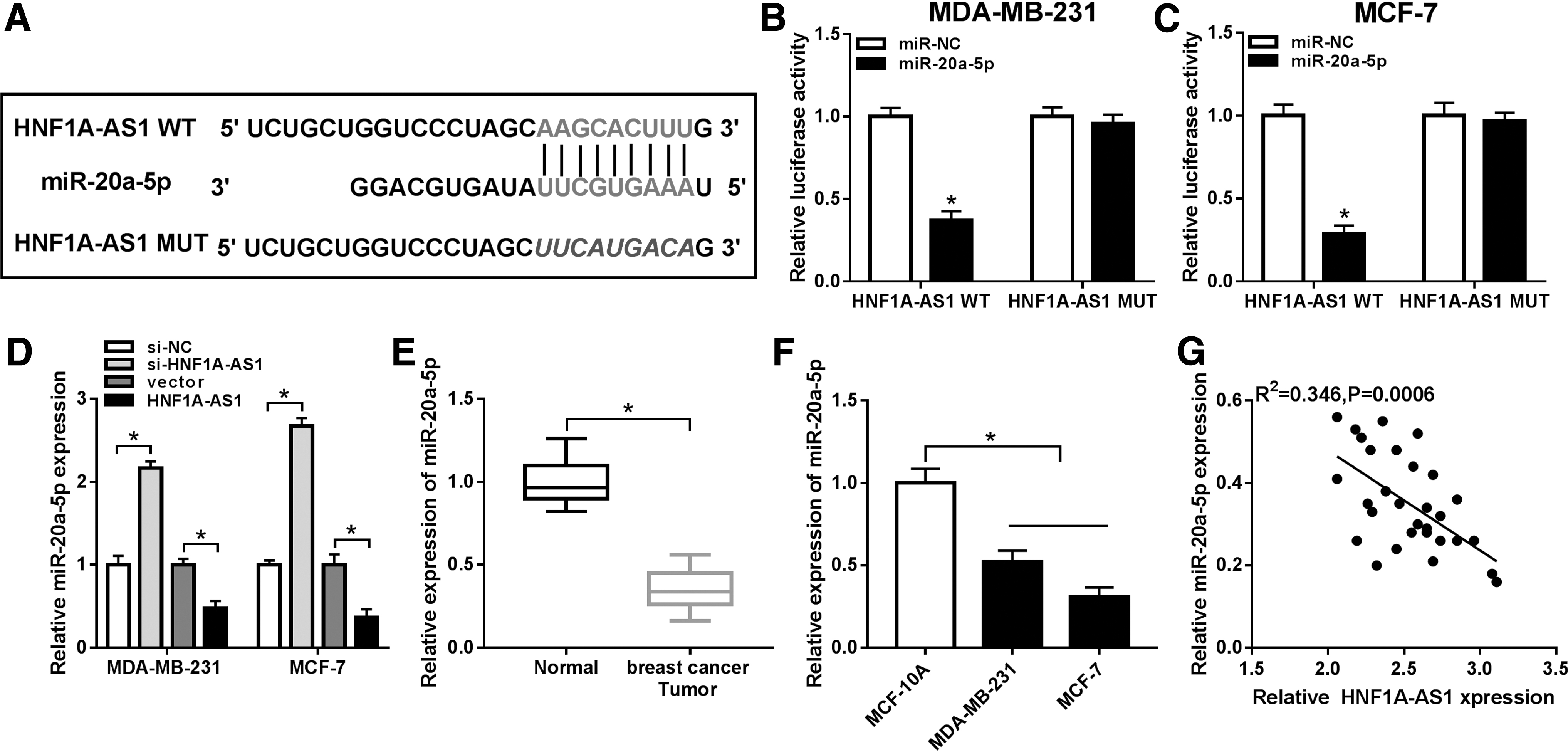
HNF1A-AS1 physically targeted miRNA-20a-5p (miR-20a-5p).
Moreover, expression level of miR-20a-5p was lower with HNF1A-AS1 overexpression vector transfection, and higher with si-HNF1A-AS1 transfection (Fig. 3D). In addition, qRT-PCR data revealed that miR-20a-5p was lowly expressed in breast cancer tumors and cell lines MDA-MB-231 and MCF-7 compared to the paired normal tissues and cell line MCF-10A, respectively (Fig. 3E, F). Notably, Pearson correlation analysis identified a negative correlation between HNF1A-AS1 and miR-20a-5p expression in these breast cancer subjects (R 2 = 0.346, p = 0.0006; Fig. 3G). These results indicated HNF1A-AS1 could control miR-20a-5p expression by direct binding.
Elevated miR-20a-5p mediated the suppression of HNF1A-AS1 deletion on breast carcinogenesis
Rescue experiments were carried out to figure out whether miR-20a-5p expression affected the biological role of HNF1A-AS1 in breast carcinogenesis. MDA-MB-231 and MCF-7 cells were transfected with si-HNF1A-AS1, together with either anti-miR-20a-5p or its control. As shown in Figure 4A and B, co-transfection with anti-miR-NC did not alter the promotion of si-HNF1A-AS1 on miR-20a-5p expression level; meanwhile, co-transfection with anti-miR-20a-5p distinctively lowered miR-20a-5p level in HNF1A-AS1-silenced cells. With co-transfection with anti-miR-20a-5p, MTS assay showed that the descended cell proliferation was improved when HNF1A-AS1 was deleted in MDA-MB-231 and MCF-7 cells (Fig. 4C, D), while FCM analysis presented an inhibition on si-HNF1A-AS1-mediated apoptosis (Fig. 4E, F).
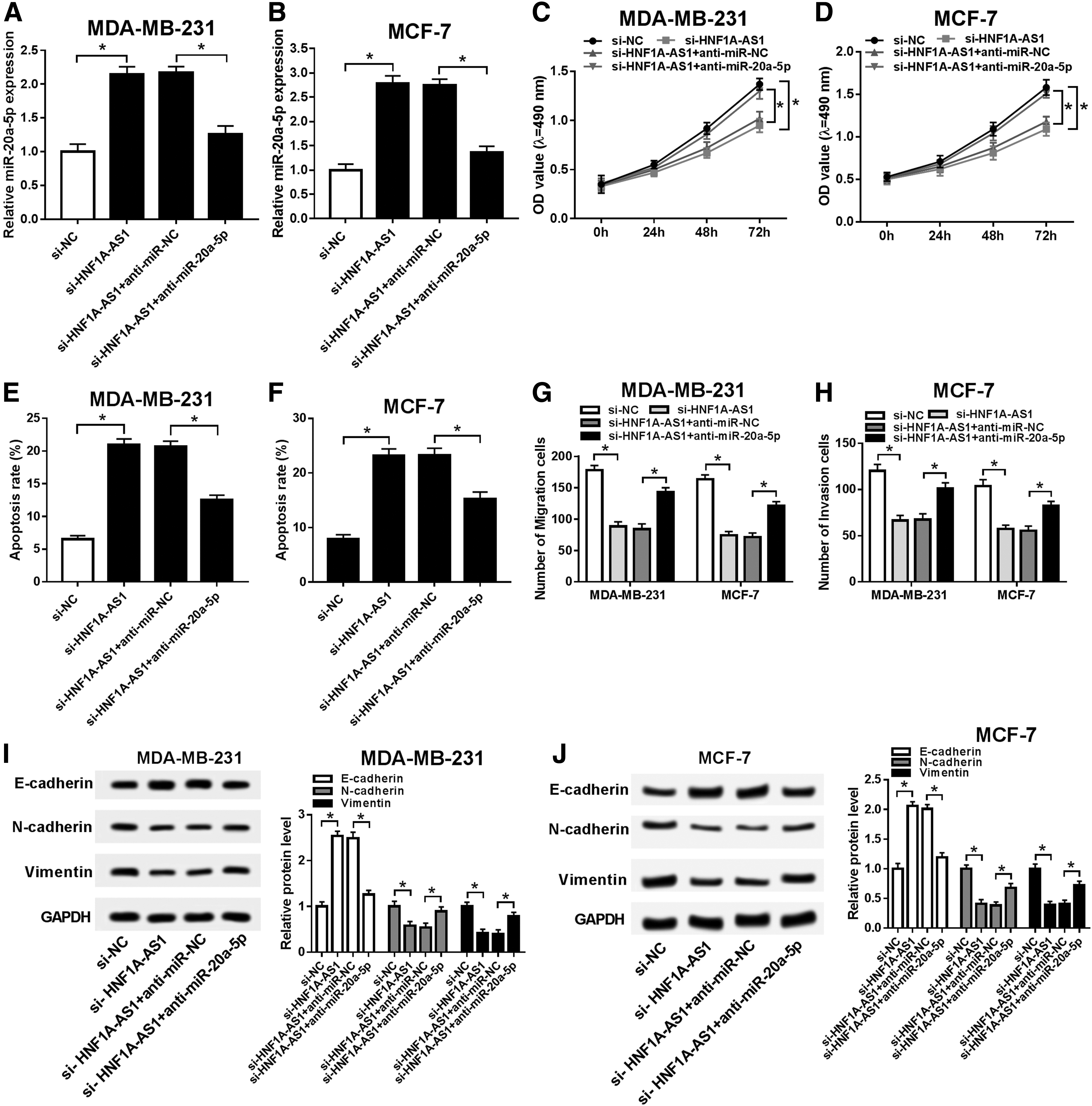
Elevated miR-20a-5p mediated the repression of HNF1A-AS1 deletion on breast carcinogenesis. MDA-MB-231 and MCF-7 cells were transfected with si-HNF1A-AS1 alone or together with miR-20a-5p inhibitors (anti-miR-20a-5p).
In terms of metastatic ability, transwell assays depicted obvious recovery of cell migration and invasion due to miR-20a-5p downregulation in HNF1A-AS1-silenced MDA-MB-231 and MCF-7 cells (Fig. 4G, H). Furthermore, E-cadherin expression was significantly reduced, whereas N-cadherin and vimentin expression were distinctively rescued by co-transfection of si-HNF1A-AS1 and anti-miR-20a-5p (Fig. 4I, J). These results showed miR-20a-5p downregulation could partially abolish the repression of HNF1A-AS1 deletion on breast carcinogenesis, indicating HNF1A-AS1 deletion exerted its antitumor role depending on elevating miR-20a-5p.
TRIM32 was a direct target of miR-20a-5p
Targetscan bioinformatics tools were utilized to explore the downstream targets of miR-20a-5p. The authors chose the conserved complementary site of miR-20a-5p in TRIM32 transcript on position of 1735–1741 (Fig. 5A). The sequence CACUUUA in TRIM32 3′ UTR WT was substituted with AUUCCAC. Vectors containing TRIM32 3′ UTR WT or MUT were transfected into MDA-MB-231 and MCF-7 cells combined with miR-20a-5p mimics or miR-NC mimics. Dual-luciferase reporter assay showed that only the relative luciferase activity of TRIM32 3′ UTR WT vector was significantly reduced with miR-20a-5p ectopic expression (Fig. 5B, C).
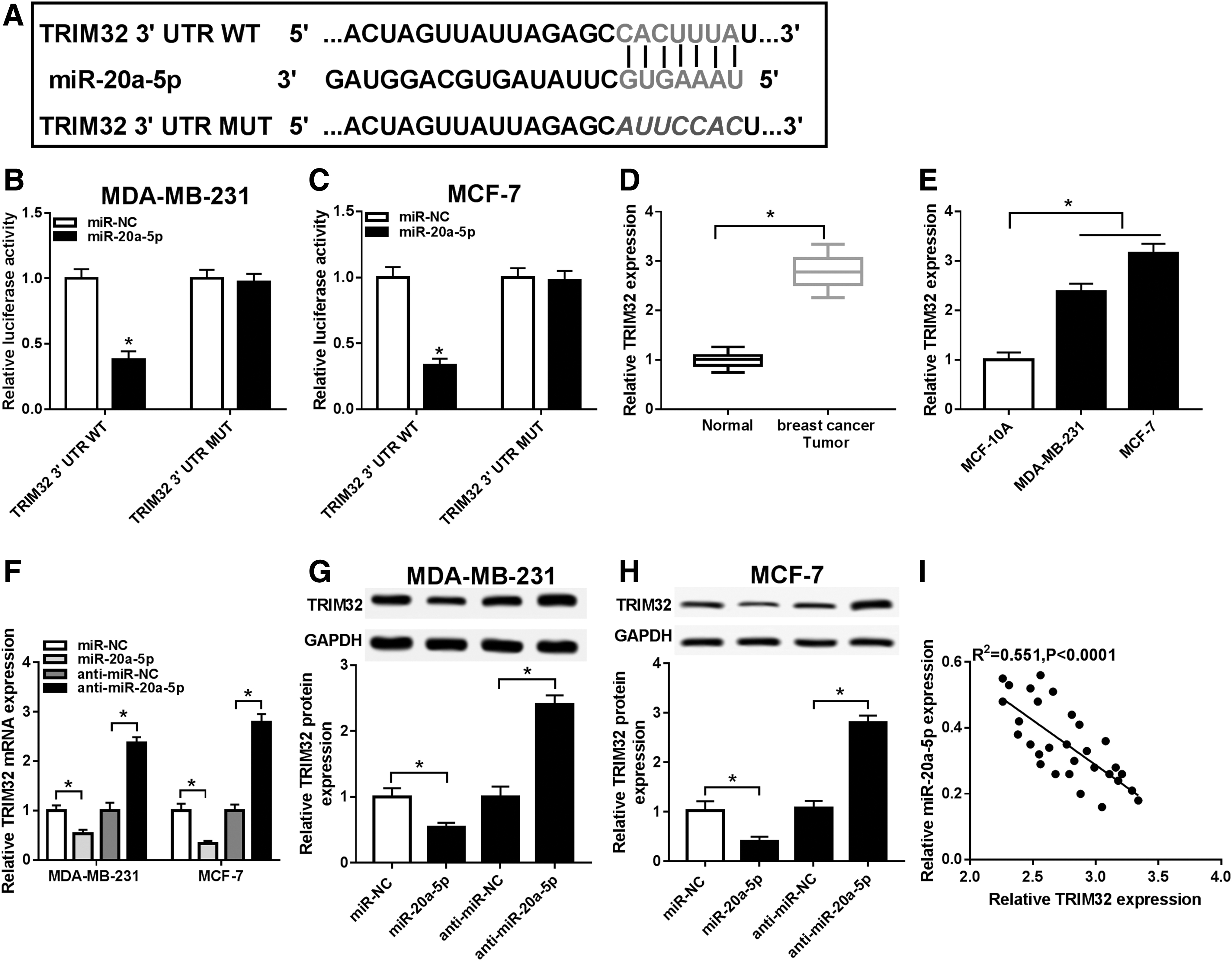
TRIM32 was a direct target of miR-20a-5p.
In addition, qRT-PCR data revealed that TRIM32 mRNA expression was highly expressed in breast cancer tumors and cell lines MDA-MB-231 and MCF-7 compared to the paired normal tissues and cell line MCF-10A, respectively (Fig. 5D, E). Moreover, expression of TRIM32 mRNA level (Fig. 5F) and protein level (Fig. 5G, H) was lower with miR-20a-5p mimics transfection, and higher with anti-miR-20a-5p transfection. Notably, Pearson correlation analysis identified a negative correlation between miR-20a-5p and TRIM32 mRNA expression in these breast cancer subjects (R 2 = 0.551, p < 0.0001; Fig. 5I). These results indicated TRIM32 was targeted and downregulated by miR-20a-5p in breast cancer cells.
Reduced TRIM32 mediated the inhibition of miR-20a-5p on breast carcinogenesis
MDA-MB-231 and MCF-7 cells were transfected with miR-20a-5p alone or together with TRIM32 overexpression vector, followed by determination of the malignant phenotypes. As a result, co-transfection with pcDNA-TRIM32 distinctively enhanced TRIM32 mRNA level (Fig. 6A, B) and protein level (Fig. 6C, D) in miR-20a-5p-overexpressed cells. With co-transfection with pcDNA-TRIM32, MTS assay showed that the depressed cell proliferation was improved by TRIM32 restoration in miR-20a-5p-upregulated MDA-MB-231 and MCF-7 cells (Fig. 6E, F), while FCM analysis presented an inhibition of pcDNA-TRIM32 on miR-20a-5p-mediated apoptosis (Fig. 6G, H).
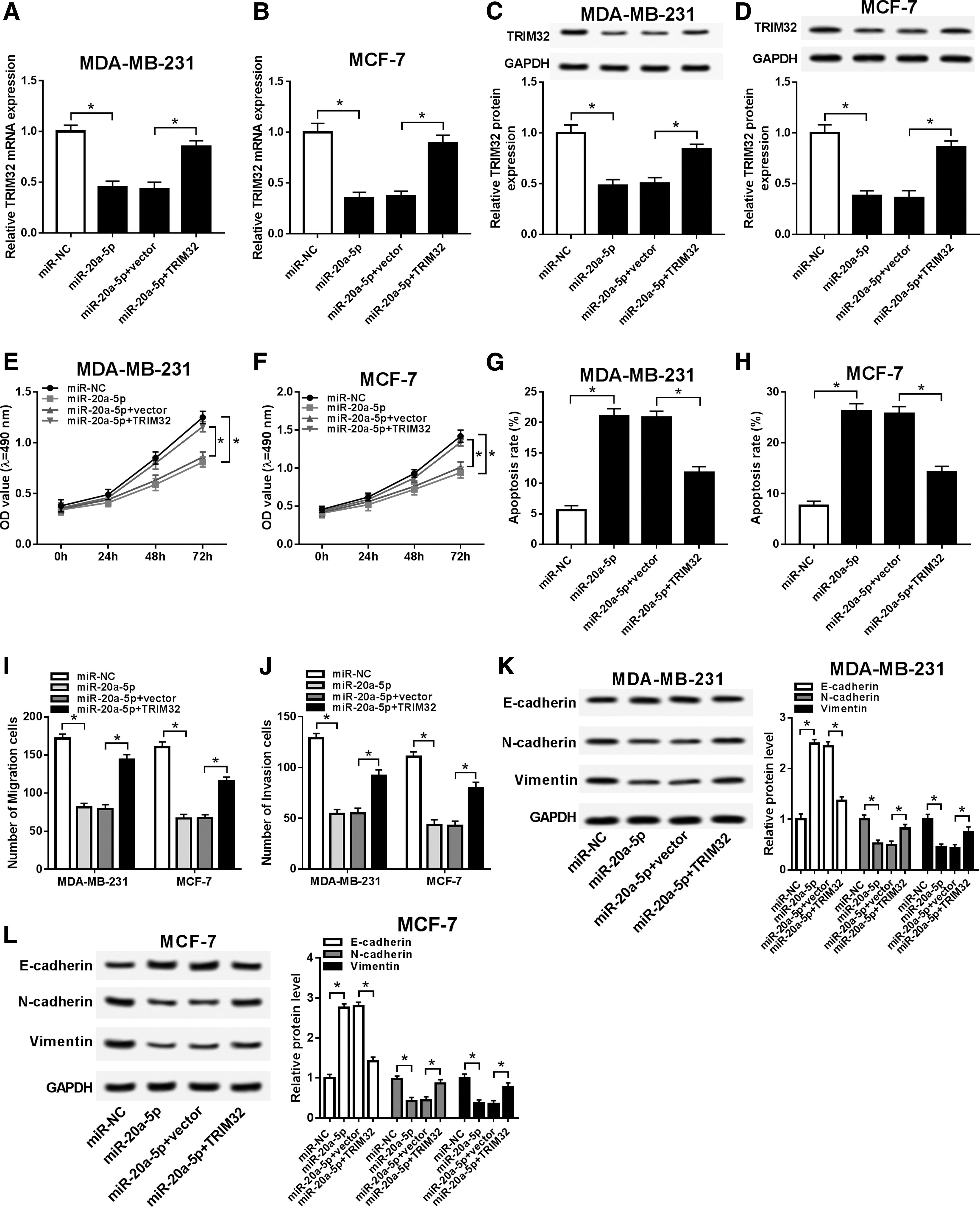
Reduced TRIM32 mediated the inhibition of miR-20a-5p on breast carcinogenesis. MDA-MB-231 and MCF-7 cells were transfected with miR-20a-5p alone or together with TRIM32 overexpression vector (TRIM32).
Transwell assays depicted an obviously increased cell migration and invasion due to TRIM32 restoration in miR-20a-5p mimics-transfected MDA-MB-231 and MCF-7 cells (Fig. 6I, J). Furthermore, E-cadherin expression was significantly reduced, whereas N-cadherin and vimentin expression were distinctively promoted by co-transfection of miR-20a-5p mimics and pcDNA-TRIM32 (Fig. 6K, L). These results demonstrated TRIM32 restoration could partially abolish the repression of miR-20a-5p on breast carcinogenesis, indicating miR-20a-5p exerted its antitumor role depending on attenuating its target TRIM32.
HNF1A-AS1 positively regulated TRIM32 expression through sponging miR-20a-5p
The authors had confirmed that miR-20a-5p could directly interact with HNF1A-AS1 and TRIM32, then whether there was a reciprocity and mutual benefit between HNF1A-AS1 and TRIM32 was unclear. Thus, the correlation of their expression levels in tumors was analyzed by Pearson correlation analysis. As shown in Figure 7A, TRIM32 mRNA expression was dramatically correlated with HNF1A-AS1 expression in this cohort of patients in a manner of linear regularity (R 2 = 0.437, p < 0.001). Moreover, the downregulatory effect of HNF1A-AS1 silencing on TRIM32 expression was broken up by anti-miR-20a-5p transfection in MDA-MB-231 and MCF-7 cells, as evidenced by qRT-PCR and Western blotting (Fig. 7B–D). This outcome demonstrated HNF1A-AS1 could positively regulate TRIM32 expression through sponging miR-20a-5p.

HNF1A-AS1 deletion downregulated TRIM32 expression through sponging miR-20a-5p.
Knockdown of HNF1A-AS1 suppressed tumor growth presumably through upregulating miR-20a-5p and downregulating TRIM32
To verify the functions of HNF1A-AS1/miR-20a-5p/TRIM32 axis in breast cancer cells in vivo, xenograft mice model was stimulated by MCF-7 cells or MDA-MB-231 cells stably expressing sh-HNF1A-AS1 or sh-NC. The nude mice generated tumors in both groups after inoculation; however, HNF1A-AS1 knockdown could slow down tumor growth (Fig. 8A, G), as well as reduced tumor weight (Fig. 8B, H). After the xenograft tumors were harvested on day 35, expression of HNF1A-AS1, miR-20a-5p, and TRIM32 was monitored. Expectedly, HNF1A-AS1 and TRIM32 levels were significantly lower in MCF-7 cell-mediated mice tumors (Fig. 8C, E, F, I, K, L), whereas miR-20a-5p level was higher (Fig. 8D, J). These results indicated knockdown of HNF1A-AS1 suppressed breast cancer tumor growth in vivo, presumably through targeting miR-20a-5p/TRIM32 axis.
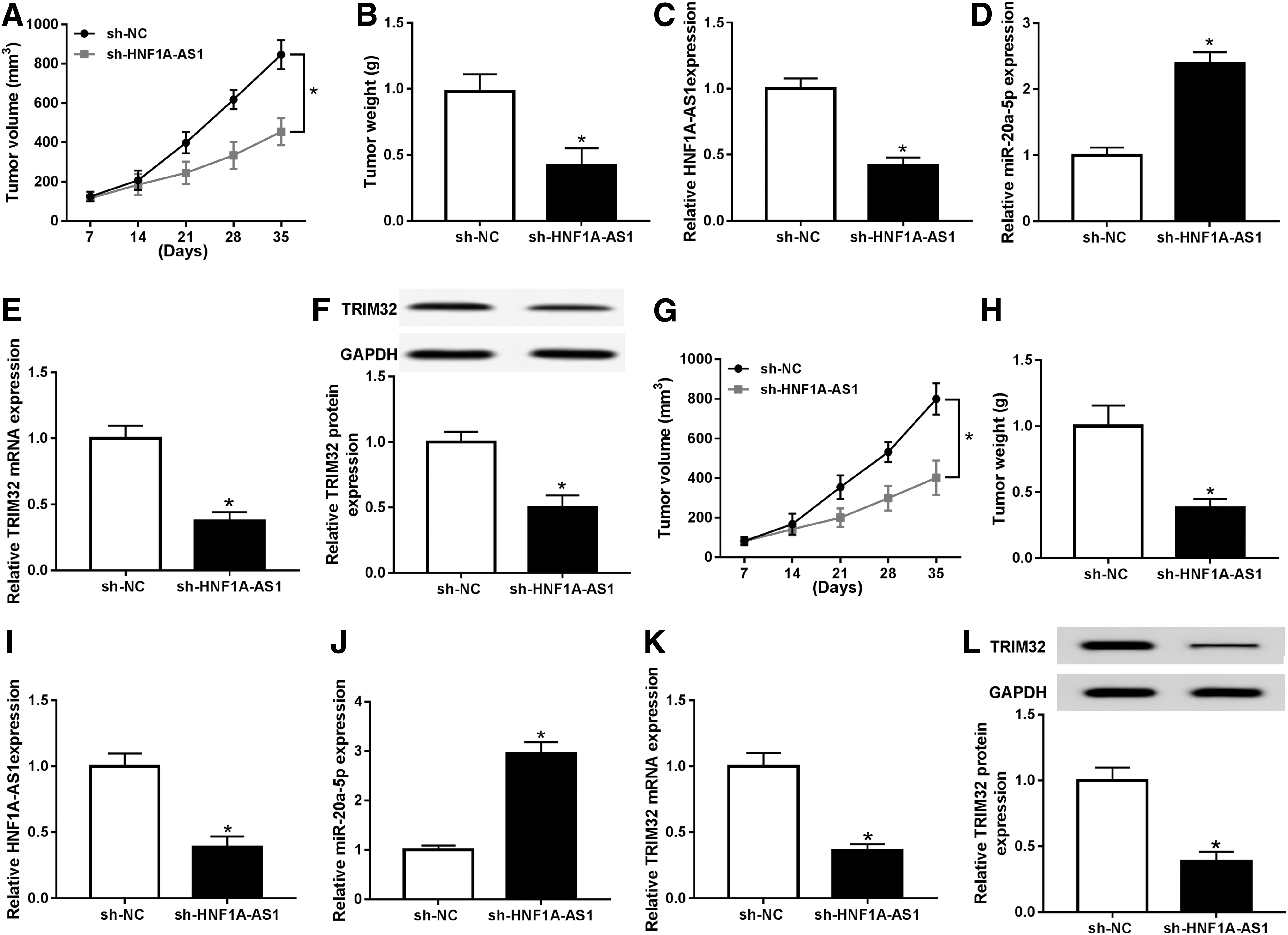
Knockdown of HNF1A-AS1 suppressed tumor growth presumably through upregulating miR-20a-5p and downregulating TRIM32. Xenograft tumors were generated by subcutaneous injection of MCF-7 cells (3 × 106) or MDA-MB-231 cells (3 × 106) stably expressing shRNA against HNF1A-AS1 (sh-HNF1A-AS1) or its negative control (sh-NC) into the right flanks of BALB/c nude mice (n = 6).
Discussion
Since identified, HNF1A-AS1 had been proposed to function as an oncogene in different human cancers and correlate with poor survival. 21 –23 Moreover, HNF1A-AS1-associated ceRNA network had been widely complicated in malignant tumor progressions, such as proliferation, cell cycle, apoptosis, autophagy, migration, and invasion. 24,25 Unfortunately, the role of this lncRNA remained little studied in breast cancer, as well as its expression status. Herein, the authors attempted to explore the expression of HNF1A-AS1 in breast cancer tissues and cell lines, then the biologic role of its dysregulation was further defined out in breast cancer cells both in vitro and in vivo. Notably, they determined a promising pathway underlying the role of HNF1A-AS1 in breast carcinogenesis through acting as a molecular sponge of miR-20a-5p.
In this study, the authors observed a higher level of HNF1A-AS1 in both breast cancer tumors and cell lines (MDA-MB-231 and MCF-7), consistent with previous studies. 21 –26 This finding suggested that HNF1A-AS1 might also play an oncogenic role in breast cancer. Later, loss-of-function experiments were carried out in MDA-MB-231 and MCF-7 cell-transfected si-HNF1A-AS1 or its negative control. As a result, the oncogenic phenotypes, including proliferation, EMT, migration, and invasion, were suppressed, but apoptosis rate was enhanced due to HNF1A-AS1 deletion, which were in agreement with results from Liu et al. 26 and Cai et al. 23
However, the expression of HNF1A-AS1 in serum from breast cancer patients and the association between HNF1A-AS1 level and clinicopathologic features such as distance metastasis, tumor stage, and overall survival were left to be further detected, 23 as well as the involvement of HNF1A-AS1 in signaling pathways such as Notch and Wnt. 26,27 Subsequently, the influence of HNF1A-AS1 on tumorigenesis in vivo was investigated researched, and it was discovered that tumor growth of MCF-7 cells and MDA-MB-231 cells in nude mice was restrained in sh-HNF1A-AS1 groups. These data supported the results reported by Fang et al. 27 and Ding et al. 28 Collectively, they confirmed the HNF1A-AS1/miR-20a-5p/TRIM32 pathway presumably underlying breast carcinogenesis (Supplementary Fig. S1), suggesting HNF1A-AS1 probably as a widespread oncogene in human malignant tumors.
In this study, the authors demonstrated that miR-20a-5p expression level could be sponged by HNF1A-AS1 in MDA-MB-231 and MCF-7 cells. Actually, among the many predicted miRNAs for HNF1A-AS1, miR-20a-5p was downregulated in breast cancer and was the most sensitive one when HNF1A-AS1 was knocked down (Supplementary Table S2). Thus, they chose miR-20a-5p for the candidate target gene for HNF1A-AS1 in this study. Similarly, Zhao et al. 16 also declared a direct interplay between miR-20a-5p and lncRNA HOTAIR in regulation of breast cancer cell behaviors. Combined these two results, they brought forward a notion that there might exist a miR-20a-5p-related ceRNA network in modulating breast carcinogenesis.
Moreover, Zhao et al. 29 also pointed three lncRNAs XIST, H19, and MALAT1 directly binding to miR-20a-5p. Meanwhile, they further revealed a higher expression of miR-20a-5p in both breast cancer tissues and serum, which supported findings from Bai et al., 30 whereas conflicted with outcomes from Si et al. 11 and us. According to qRT-PCR analysis, the authors discovered that the basic expression of miR-20a-5p was depressed in breast cancer tissues and cell lines (MDA-MB-231 and MCF-7) compared to normal adjacent tissues and normal breast cell line MCF-10A.
Functionally, high expression of miR-20a-5p could impair cell proliferation, EMT, migration, and invasion in MDA-MB-231 and MCF-7 cells, while its silencing could improve HNF1A-AS1 knockdown-mediated inhibition of these oncogenic progressions. And the role of miR-20a-5p mentioned above agreed with previous study. 16 However, expression and role of this miRNA in breast cancer were not identical in varying researches, and the mechanism of this inconformity had not been discussed. All in all, these studies illustrated a key role of miR-20a-5p in tumorigenesis of breast cancer through being sponged by lncRNAs.
TRIM32 was frequently overexpressed in different types of human tumors such as neuroblastoma, lung cancer, gastric cancer, and breast cancer. 31 –34 With bioinformatics analysis, TRIM32 and many other functional genes were predicted to be potential targets of miR-20a-5p, and the authors further confirmed the target binding between miR-20a-5p and TRIM32 because TRIM32 was the most sensitive candidate gene with miR-20a-5p overexpression (Supplementary Table S3). Expression of TRIM32 was markedly upregulated in breast cancer tissues and cell lines (MDA-MB-231 and MCF-7), which was in favor of previous studies. 34,35
Functionally, increased TRIM32 could display oncogenic role that partially rescues cell proliferation, EMT, migration, invasion, and apoptosis inhibition in miR-20a-5p-overexpressed MDA-MB-231 and MCF-7 cells. Besides, Zhao et al. 34 claimed a positive relationship between TRIM32 expression and chemosensitivity through nuclear factor kappaB (NF-κB) signaling pathway.
In terms of the molecular mechanism of TRIM32 in breast cancer, Liu et al. 35 also declared p53 could transcriptionally induce TRIM32 expression, and TRIM32 could, in turn, promote p53 degradation. Thus, TRIM32 participated in apoptosis, cell cycle, and senescence in tumorigenesis. Furthermore, HNF1A-AS1-mediated ceRNA network had been demonstrated to be complicated with a feedback loop of p53 in colon cancer. 27 Therefore, the authors put forward a hypothesis that HNF1A-AS1/miR-20a-5p/TRIM32 axis contributed to breast cancer development, presumably in a p53-dependent manner (Supplementary Fig. S2). Nevertheless, this should be further tested using additional experimentation.
Besides, the detail mechanism of HNF1A-AS1/miR-20a-5p/TRIM32 axis, including signaling pathways such as NF-κB, was not investigated in this study. Also, ABI1 and NF-κB inhibitor Piasy were declared to be target genes of TRIM32 for ubiquitination, and then degraded 36,37 (Supplementary Fig. S2). In terms of target gene of miR-20a-5p, that multiple TRIM proteins such as TRIM3, 8, 32, 36, 37, 44, 66, and 71 were predicted to have complementary binding site of miR-20a-5p and this is of value to be verified in the future.
Conclusions
Taken together, the authors showed HNF1A-AS1 was highly expressed in breast cancer tumors and cell lines, and its deletion could repress the malignant behaviors of breast cancer cells both in vitro and in vivo through directly upregulating miR-20a-5p and downregulating TRIM32, a downstream target gene of miR-20a-5p. These results provide a novel HNF1A-AS1/miR-20a-5p/TRIM32 pathway in breast carcinogenesis, and suggest HNF1A-AS1 could be a potential target for the occurrence, development, and treatment of human breast cancer.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.