
Abstract
Background:
Long noncoding RNAs could serve as a candidate target for prostate cancer (PCa) diagnosis and treatment. The current study aimed to investigate the role and functions of SNHG1 in PCa cells.
Materials and Methods:
Abnormal expression of SNHG1, survival analysis, and target gene were determined or predicted by bioinformatics techniques. Gene expressions at transcriptional and translational levels were determined by Quantitative Real-time PCR and Western blotting, respectively. Cell viability, growth, and apoptosis rate were detected by Cell Counting Kit-8, colony formation assay and flow cytometry.
Results:
The results showed that SNHG1 was highly expressed in PCa tissues, which was accompanied by decreased miR-377-3p expression and poor overall survival rate, and that miR-377-3p was predicted as the target of SNHG1 in PCa cells. Moreover, SNHG1 counteracted the effects of miR-377-3p on inhibiting cell growth and promoting apoptosis of PCa cells. Furthermore, miR-377-3p counteracted the effects of AKT2 on promoting cell viability, growth, and suppressing apoptosis of PCa cells. In addition, AKT2 expression was proved to be regulated by miR-377-3p.
Conclusions:
The SNHG1/miR-377-3p/AKT2 regulatory axis in PCa cells was disclosed. The upregulated AKT2 might be a result of dysregulated interaction balance between the expressions of miR-377-3p and SNHG1. Based on such discoveries, the intervention of SNHG1/miR-377-3p/AKT2 axis could be further explored in the treatment of PCa.
Introduction
Prostate cancer (PCa) is a frequently diagnosed cancer in male urogenital system all over the world. 1 PCa has the second highest incidence among all malignant tumors in men, 2 and the number of PCa cases diagnosed in China is increasing annually. 3 According to the latest data from the National Cancer Center of China, PCa has become the most common tumor in male urogenital and reproductive system since 2008, with ninth highest incidence among all male malignancies. 4 One of the most important risk factors for developing PCa is transmissibility. 5 At present, as a molecular marker for the diagnosis of PCa, serum prostate-specific antigen (PSA) has been widely used in clinical practice. 6 However, due to the influence of benign prostatic hyperplasia, inflammation, age, drug and iatrogenic factors, PSA specificity, and sensitivity remain controversial. 7 Radiotherapy, androgen deprivation therapy and chemotherapy are the main treatments for PCa, and are used according to the condition of each patient. 8 –11 However, none of the currently available approaches was effective in treating PCa, due to the lack of regulatory network of PCa-related genes and long noncoding RNAs (lncRNAs).
lncRNAs are a transcriptional product of RNA polymerase II (RNAP II, Pol II) and is ubiquitous in the nucleus or cytoplasm of eukaryotic cells. 12 –17 The structure of lncRNAs is similar to that of mRNAs, but they do not have protein-coding function. 12 –17 lncRNAs are found abnormally expressed in various conditions, affecting disease progressions, including cancers. 12 –17 Hence, the abnormality of the expression of lncRNA could indicate and reflect cancer stage, tumor-associated signaling pathways, and the efficacy of anticancer therapy. 12 –17 Today, with the widespread use of high-throughput sequencing and bioinformatics analysis, a large number of PCa-related lncRNAs are screened. 16 Studies have revealed the role of aberrantly expressed lncRNAs in PCa and its effects on the cell proliferation, invasion, metastasis, apoptosis, and suggested that lncRNAs could be explored as molecular candidate target for PCa diagnosis and treatment. 18 –21
Recently, a study found that SNHG1 expression was related to tumorigenesis and prognosis. 22 However, few studies explored the role of SNHG1 in PCa. Thus, the current study investigated the role of SNHG1 in PCa cells and determined if miRNA or gene SNHG1 interacted to exert its function.
Materials and Methods
Bioinformatics and sample collection
Based on the mean or median expression level of SNHG1 in the Cancer Genome Atlas (TCGA) database, differences in the expression of SNHG1 in the database between normal tissues and PCa tissues and survival analysis of patients were analyzed. Tissue samples from patients diagnosed with PCa were extracted during surgery following the Examination Ethics of the Affiliated Yantai Yuhuangding Hospital of Qingdao.
Cell culture
We purchased 22Rv1, LNCaP cells and were transported from American Type Culture Collection. All the cells were incubated in RPMI 1640 (21875091; Thermo Fisher, Waltham) medium with 10% fetal bovine serum (F8192; Sigma-Aldrich, MO) in 5% CO2 at 37°C.
Design
For determining the interaction of SNHG1 and miR-377-3p, 22Rv1 cells were divided into six groups, namely, Control, pc-Control, pc-SNHG1, mimic negative control (NC), mimic, and pc-SNHG1 + mimic. The cells in the Control group were treated by culture medium only, whereas those in rest of the groups were respectively transfected with pc-SNHG1 vector (GenePharma, Shanghai, China), pc-SNHG1 (GenePharma), miR-377-3p mimic NC (GenePharma), miR-377-3p mimic (GenePharma), and pc-SNHG1 with miR-377-3p mimic. LNCaP cells were divided into six groups, namely, Control, si-Control, si-SNHG1, inhibitor NC, inhibitor, and si-SNHG1 + inhibitor groups. The cells in the Control group were treated by culture medium only, whereas those in rest of the groups were respectively transfected with si-SNHG1 vector (GenePharma), si-SNHG1 (GenePharma), miR-377-3p inhibitor NC (GenePharma), miR-377-3p inhibitor (GenePharma), and si-SNHG1 with miR-377-3p inhibitor.
To determine the connection between AKT2 and miR-377-3p, 22Rv1 cells were divided into five groups, namely, Control, NC, AKT2, AKT2 + mimic, and mimic groups. The cells in the Control group were treated by culture medium only, whereas those in rest of the groups were respectively transfected with AKT2 vector (GenePharma), AKT2 (GenePharma), AKT2 with miR-377-3p mimic, and miR-377-3p mimic. LNCaP cells were divided into five groups, namely, Control, siNC, siAKT2, siAKT2 + inhibitor, and inhibitor groups. The cells in the Control group were treated by culture medium only, whereas those in rest of the groups were respectively transfected with siAKT2 vector (GenePharma), siAKT2 (GenePharma), siAKT2 with miR-377-3p inhibitor, and miR-377-3p inhibitor. The experiments were conducted in accordance with the instructions of Lipofectamine 3000 (L3000015; Thermo Fisher).
Quantitative polymerase chain reaction
The mRNA expressions of SNHG1, miR-377-3p, and AKT2 were determined by quantitative polymerase chain reaction (qPCR). Total RNAs were extracted using TRIzol reagent (15596018; Thermo Fisher) from PCa and adjacent tissues and 22Rv1 and LNCaP cells (2 × 105 cells). cDNAs of SNHG1, miR-377-3p, and AKT2 were synthesized by the PrimeScript RT Reagent Kit (TaKaRa Biotechnology Co., Ltd., Dalian, China). One microliter forward primer (10 μM), reverse primer 1 μL (10 μM), 8 μL cDNA template, 10 μL 2 × SYBER Green master mix (4913850001; Roche, Shanghai, China), and 5 μL ddH2O were mixed together. The PCRs were prepared and performed in Bio-Rad IQ5 thermocycler (Bio-Rad, CA) as follows: at 95°C for 90 s, at 95°C for 20 s, at 65°C for 20 s, at 72°C for 40 s, for 40 cycles. Relative transcriptional levels were calculated by 2−ΔΔCt method. The primers used were listed in Table 1.
Primers Used in the Study
lncRNA, long noncoding RNA.
Luciferase assay
The interactions between SNHG1 and miR-377-3p or between AKT2 and miR-377-3p were predicted by starBase Version 3.0 or LncBase Predicted Version 2.0. For comparison, mutant of SNHG1 or AKT2 was generated by the Quick-Change Site-Directed Mutagenesis Kit (Stratagene, CA). pRL-TK (Promega, WI) was transfected with miR-377-3p mimic by Lipofectamine 3000 (L3000015; Thermo Fisher). Relative luciferase activity was measured according to the assessment of absolute luciferase activity by the Dual-Glo Luciferase Assay Kit (Promega).
Cell counting kit (CCK-8)
The cell viability was determined at 24, 48, and 72 h after the treatments. Briefly, 2 × 105 cells were subjected to Cell Counting Kit-8 (CCK-8) test following the instruction of CCK-8 (96992-100TESTS-F; Sigma-Aldrich). Eventually, optical density in each group was read on microplate reader (Multiskan; Thermo, Waltham) at 450 nm.
Colony formation assay
LNCaP and 22Rv1cells were digested, counted, and prepared to the concentration of 1 × 106 cells/mL. The cell suspension was then added to a six-well culture plate. The cells were cultured at 37°C with 5% CO2 in an incubator for 14 d. Culture was terminated when the number of macroscopic clones reached ∼50–150. The cells were then washed twice by phosphate-buffered saline (PBS) and treated by 1 mL methanol for 15 min, rinsed in running water, and then stained by 1 mL 0.2% Crystal Violet for 30 min. When staining solution was dried in a fume hood, the number of colony formation was counted under a microscope.
Apoptosis assessment
LNCaP and 22Rv1cells were digested, collected, and centrifuged at 1000 r/min for 4 min at 4°C, and adjusted to 1 × 106 cells/mL. The cells were then resuspended and washed twice with 1 mL cold PBS and then by 200 μL binding buffer. Next, the cells were treated by fluorescein isothiocyanate-labeled Annexin V 10 and 5 μL Propidium Iodide for 15 min at room temperature (Annexin V-FITC Kit; Sigma-Aldrich), and then resuspended in 300 μL binding buffer. Cell apoptosis rate was detected immediately by flow cytometry (BD FACSVerse™; Becton Dickinson, San Jose).
Western blot
The translational level of AKT2 was determined by Western blot. LNCaP and 22Rv1cells were digested and diluted to 1 × 106 cells/mL, and total proteins in cells were extracted by RIPA lysate (R0278; Sigma-Aldrich) and protease inhibitor (S8830; Sigma-Aldrich). Next, the proteins were separated on 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis at 100 V for 1.8 h. The proteins were further transferred to a polyvinylidene fluoride membrane and blocked by 5% nonfat milk for 1 h. AKT2 antibody (55 kD, 1:1000, ab175354; Abcam, San Francisco,) and β-actin antibody (ab8226, 42 kD, 1:1000; Abcam) were mixed in the milk and incubated with the membrane at 4°C overnight. Furthermore, after washing the membrane by PBS with 0.1% Tween, the proteins were probed by Goat anti-mouse secondary antibody (1:2000, ab205719; Abcam), and developed by ECL (#6883; SignalFire™ ECL Reagent).
Statistics
The data in this study were analyzed by GraphPad prism version 8.0. Comparison between two individual groups was performed by Student t test, and multiple comparisons were conducted by one-way analysis of variance or two-way analysis of variance. Comparisons within two groups in multiple groups were conducted by Tukey's test. Survival analysis was performed by Kaplan–Meier method and examined by Log-rank test. The correlation between the expressions of SNHG1 and miR-377-3p was evaluated by Pearson method. p < 0.05 was considered as statistically significant.
Results
SNHG1 was highly expressed in PCa tissue accompanied by a decrease of miR-377-3p expression
The expressions of SNHG1 and miR-377-3p and their relationship were determined and evaluated by qPCR and bioinformatics techniques. We observed the overexpression of SNHG1 in cancer tissue compared with normal tissue, and this was related to a more unfavorable prognosis (Fig. 1A, B). Furthermore, in the tissues derived from PCa patients, SNHG1 expression was higher than that in adjacent tissue (Fig. 1C, **p < 0.05). However, miR-377-3p expression was found decreased in cancer tissues compared with in adjacent tissue, showing a negative correlation with SNHG1 expression (Fig. 1D, E, **p < 0.05). The results indicated that increased SNHG1 expression might contribute to poor prognosis of PCa, and that miR-377-3p may be involved in the regulatory pathway of SNHG1.
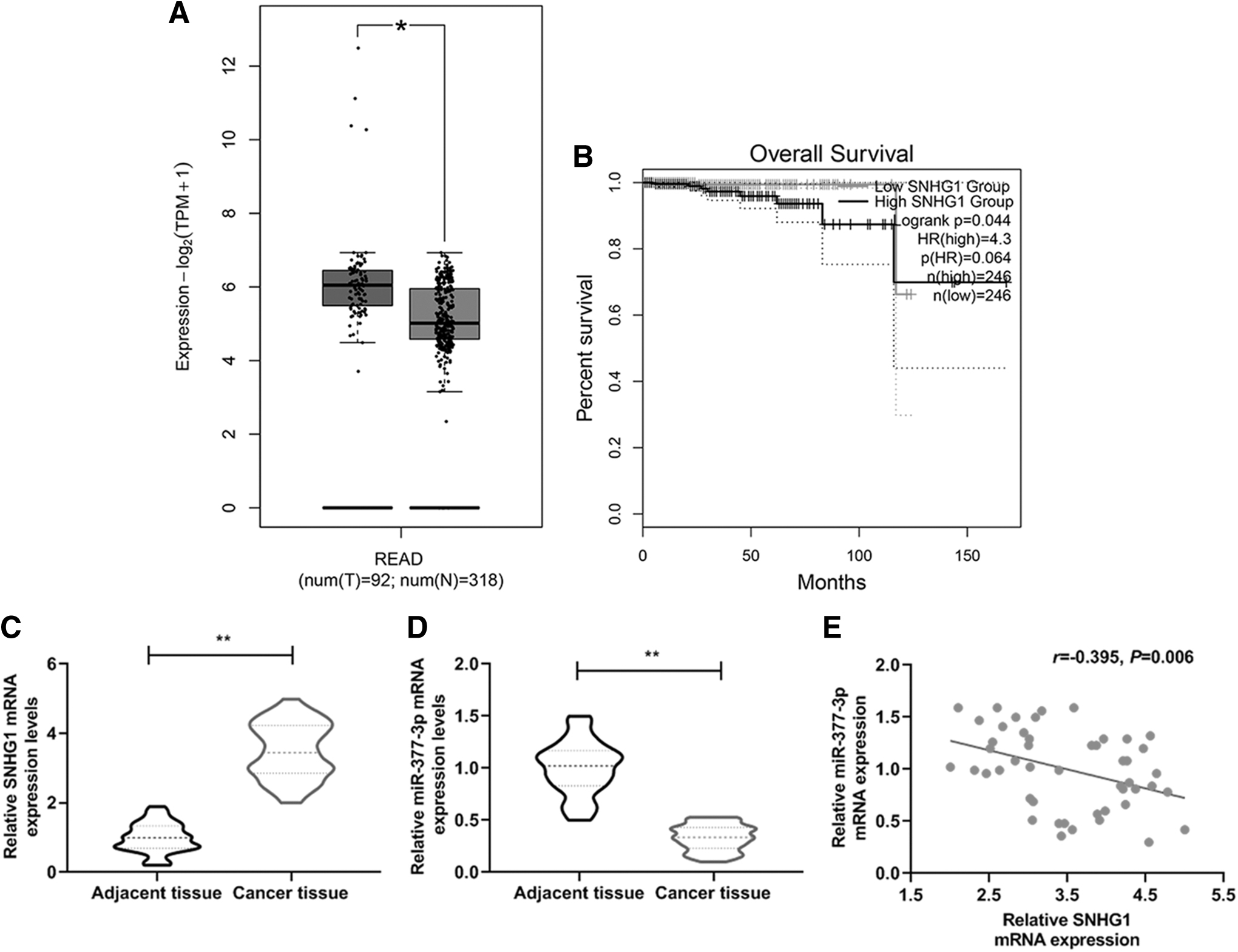
SNHG1 was highly expressed in prostate cancer tissue accompanied by the decreasing of miR-377-3p expression and overall survival rate.
miR-377-3p was the target of SNHG1 in PCa cells
The relationship between the expressions of SNHG1 and miR-377-3p was further confirmed by bioinformatics techniques and luciferase assay, and the expressions of SNHG1 and miR-377-3p in the presence or absence of one another were determined. In this study, we discovered the complementary sequence in both genes (Fig. 2A), and the relative luciferase activity was lowered than that in Blank group when miR-377-3p was transfected into 22Rv1 (Fig. 2B, **p < 0.01), or into LNCaP cells (Fig. 2C, **p < 0.01) with wild-type SNHG1. Furthermore, in 22Rv1 or LNCaP cells, the relative SNHG1 mRNA expression in pc-SNHG1 group was significantly higher than that in pc-Control group, accompanied by the increase of SNHG1 mRNA expression in the pc-SNHG1 + mimic group compared with the mimic group (Fig. 2D, **p, ΩΩ p < 0.01). The relative SNHG1 mRNA expression in si-SNHG1 group and si-SNHG1 + inhibitor group was lower compared with si-Control group and inhibitor group, respectively (Fig. 2E, ΔΔ p, ΨΨ p < 0.01). The relative miR-377-3p mRNA expression in pc-SNHG1 group was significantly lower than that in pc-Control group, accompanied by the decrease of miR-377-3p mRNA expression in the pc-SNHG1 + mimic group compared with mimic group (Fig. 2F, **p, ΩΩ p < 0.01). The relative miR-377-3p mRNA expression in si-SNHG1 group and si-SNHG1 + inhibitor group was higher compared with si-Control group and inhibitor group, respectively (Fig. 2G, ΔΔ p < 0.01, ΨΨ p < 0.05). Thus, it was proved that miR-377-3p could interact with SNHG1 in PCa cells.
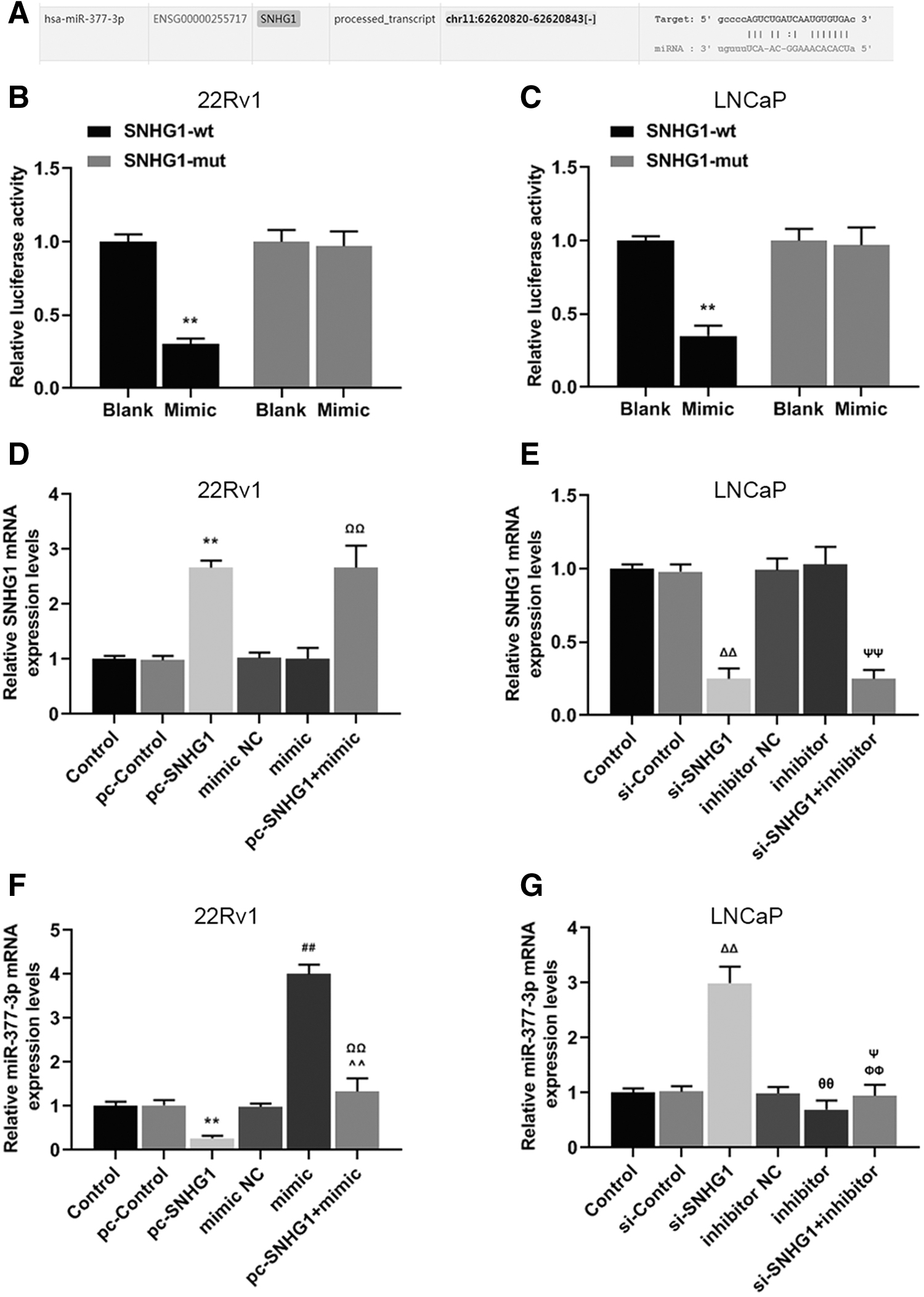
miR-377-3p was the target of SNHG1 in prostate cancer cells.
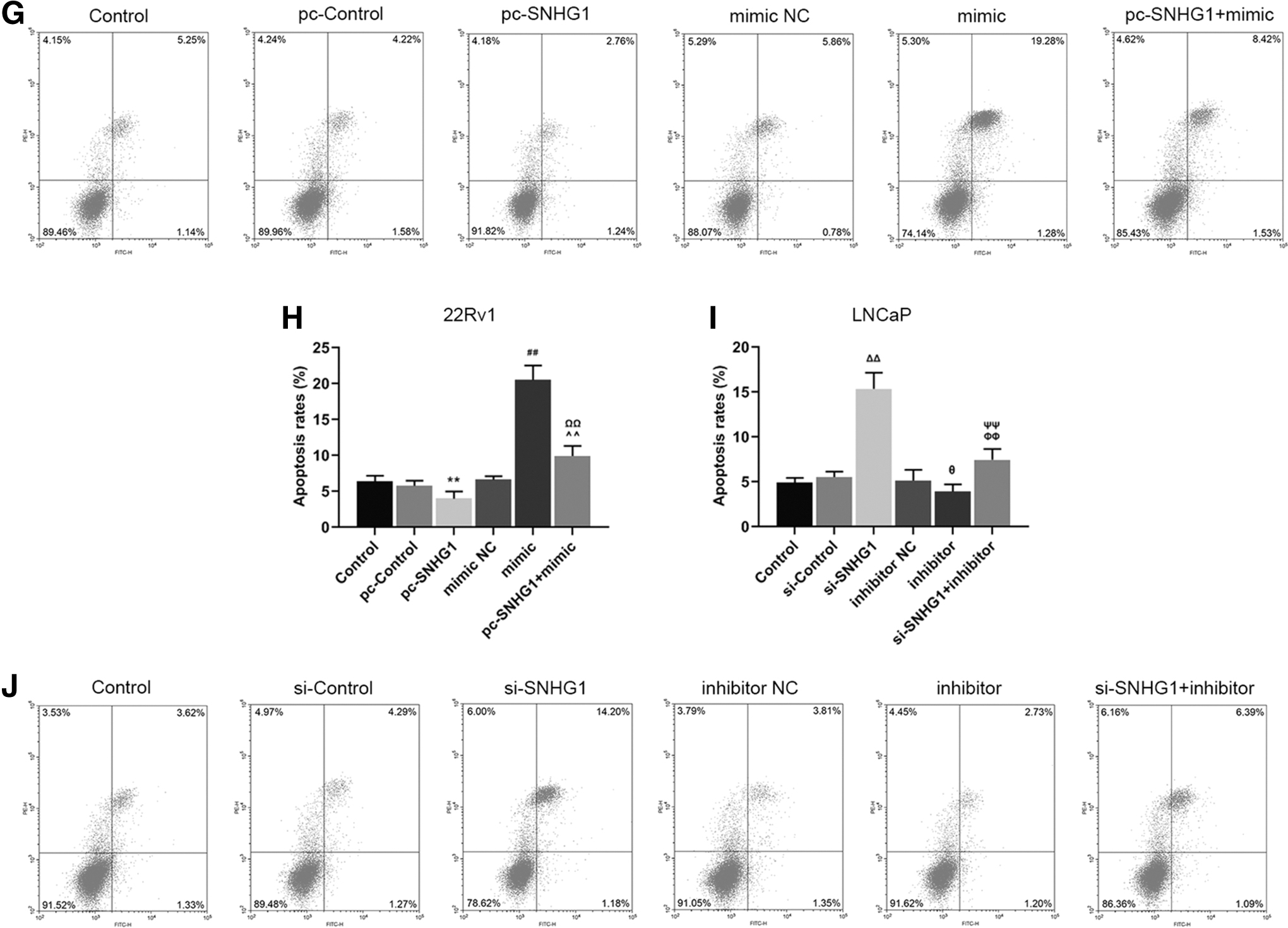
SNHG1 counteracted the effects of miR-377-3p on inhibiting viability of PCa cells
To determine the effects of SNHG1 and miR-377-3p on PCa promotion, we measured the cell viability, growth, and apoptosis rates of 22Rv1 and LNCaP cells with regulated expression levels of SNHG1 and miR-377-3p. It was found that the relative cell viability rate of 22Rv1 in pc-SNHG1 group was higher than that in pc-Control group, but lower in mimic group compared with that in mimic NC group after 48 and 72 h of culturing (Fig. 3A, **p, ## p < 0.01). Noticeably, the relative cell viability rate in pc-SNHG1 + mimic group was higher than that in mimic group, but lower than that in pc-SNHG1 group (Fig. 3A, ΩΩ p, ^^p < 0.01), moreover, a similar result was also observed in the relative colony formation rate of 22Rv1 cells (Fig. 3C, D, **p, ## p, ΩΩ p, ^^p < 0.01) and in apoptosis rate of LNCaP cells (Fig. 3I, J, θ p < 0.05, ΔΔ p, ## p, ΦΦ p, ΨΨ p < 0.01). Correspondingly, the above results of the relative cell viability, colony formation rate, and apoptosis rate were opposite in 22Rv1 and LNCaP cells (Fig. 3B, E–H, Φ p, Δ p < 0.05, ΔΔ p, θθ p, ΦΦ p, ΨΨ p, **p, ## p, ΩΩ p, ^^p < 0.01). Taken together, the findings showed that miR-377-3p acted as a PCa suppressor, while overexpressed SNHG1 could counteract such a trend to some extent.
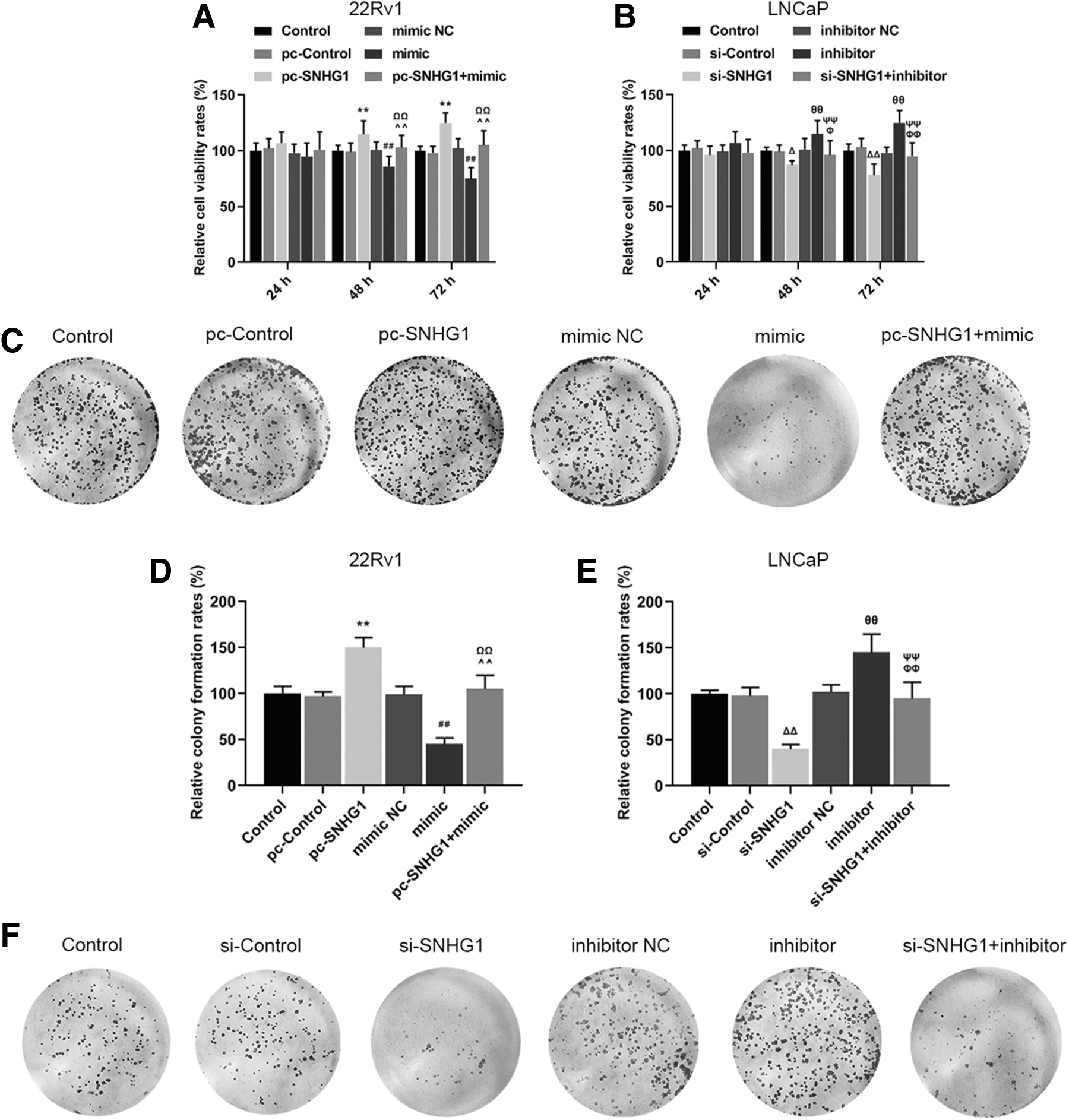
SNHG1 counteracted the favorable effects of miR-377-3p on the inhibition of promotion of prostate cancer cells.
miR-377-3p counteracted the effect of AKT2 on promoting the viability of PCa cells
AKT2 was predicted as a possible target gene, as it shared a part of complementary sequence with miR-377-3p (Fig. 4A). The result of luciferase assay showed that the relative luciferase activities of 22Rv1 and LNCaP cells with wild-type AKT2 decreased compared with the Blank group (Fig. 4B, C, **p < 0.01). Moreover, the relative cell viability of 22Rv1 in AKT2 group was higher than that in NC group, but lower in mimic group (Fig. 4D, *p < 0.05, **p, ## p, ΩΩ p < 0.01), however, the relative cell viability of LNCaP cells was opposite to that of 22Rv1 when it was treated by siAKT2 or miR-377-3p inhibitor (Fig. 4E, Δ p, θ p < 0.05, ΔΔ p, θθ p, ΦΦ p < 0.01). Data suggested that the effect of AKT2 on the proliferation of PCa cells could be counteracted partially by miR-377-3p.
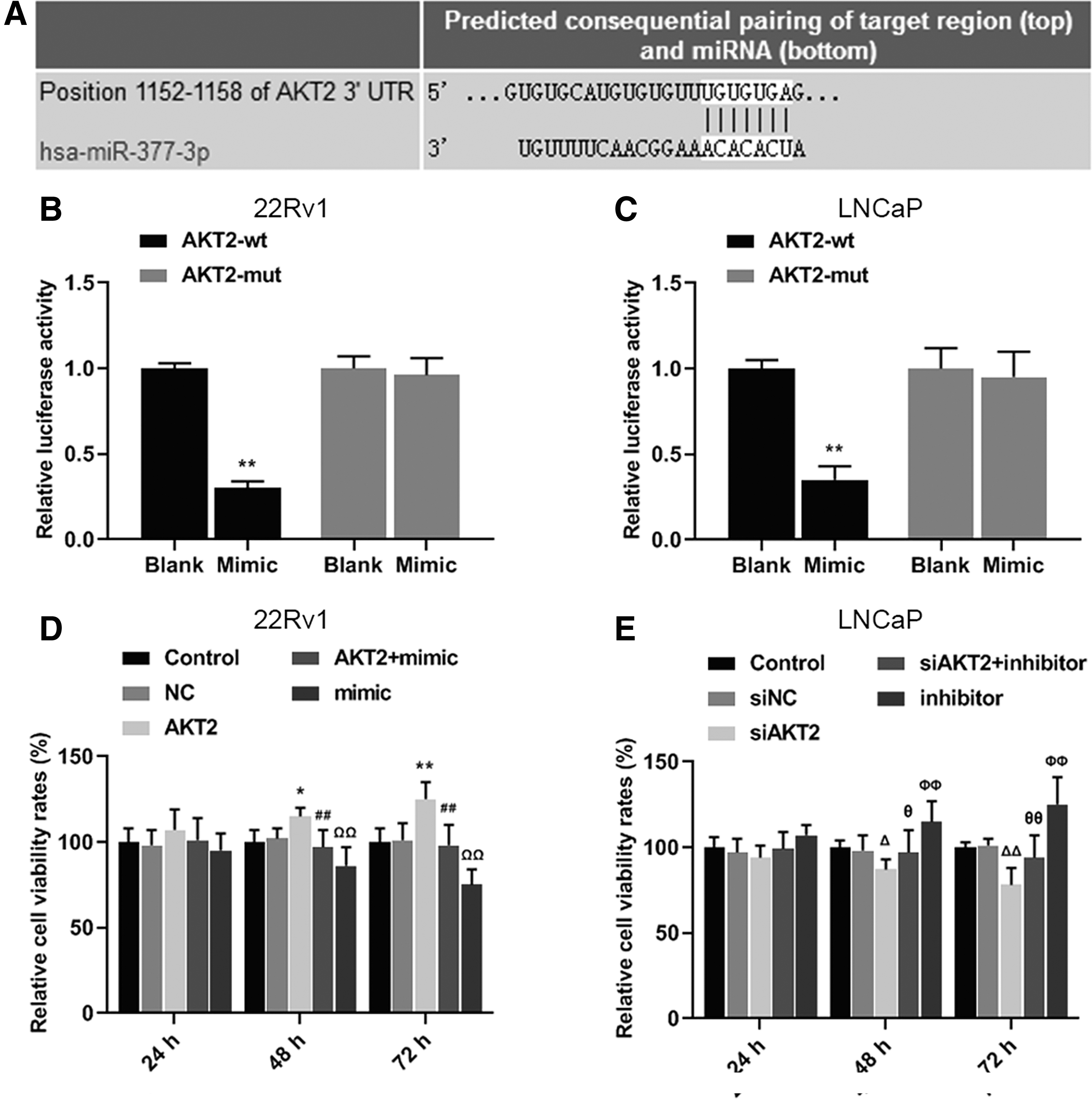
The interactive relationship of miR-377-3p and AKT2 as well as their counteraction on the cell viability of prostate cancer cells.
miR-377-3p counteracted the effect of AKT2 on promoting the growth and suppressing apoptosis of PCa cells
We explored the effects of AKT2 with miR-377-3p on the growth and apoptosis rate of PCa cells. In this study, we found that the relative colony formation rate of 22Rv1 in AKT2 group and apoptosis rate of LNCaP cells in siAKT2 group was elevated compared with NC and siNC group (Fig. 5A, D, **p, ΔΔ p < 0.01). Noticeably, the relative colony formation rate of 22Rv1 in AKT2+mimic group and apoptosis rate of LNCaP cells in siAKT2 + inhibitor group was reduced compared with AKT2 and siAKT2 group (Fig. 5A, D, ## p, ΩΩ p, θθ p, ΦΦ p < 0.01). Reversely, the colony formation rate of LNCaP cells in siAKT2 group and apoptosis rate of 22Rv1 cells in AKT2 group was reduced compared with siNC and NC group, and the colony formation rate of LNCaP cells in siAKT2 + inhibitor group and apoptosis rate of 22Rv1 cells in AKT2 + mimic group was increased compared with siAKT2 and AKT2 group (Fig. 5B, C, **p, ΔΔ p, ## p, ΩΩ p, θθ p, ΦΦ p < 0.01). Thus, the effect of AKT2 on the growth and apoptosis rate of PCa cells was eliminated by miR-377-3p to some extent.
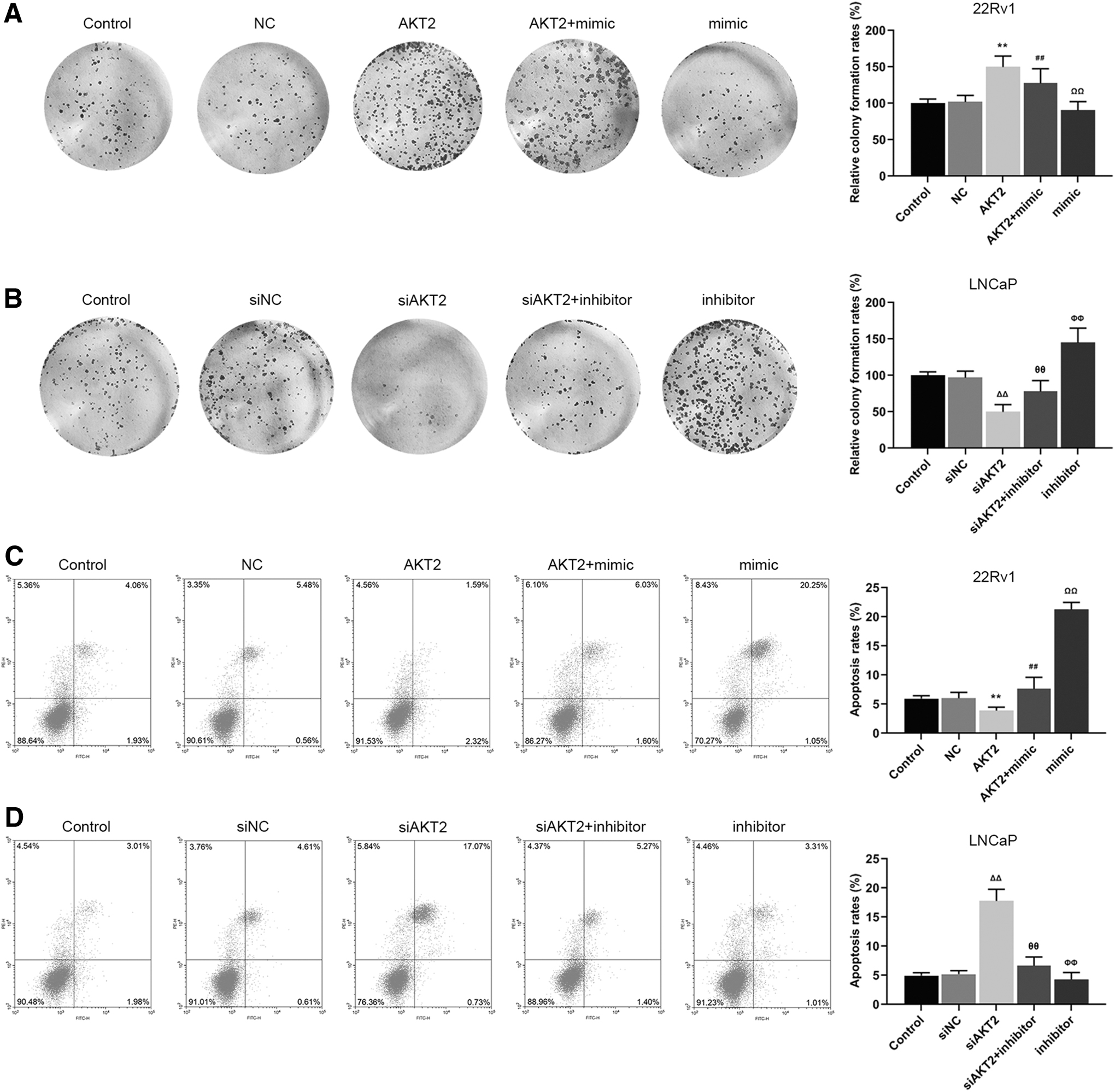
miR-377-3p counteracted the positive effect of AKT2 on the cell growth and negative effect on apoptosis of prostate cancer cells.
AKT2 expression could be regulated by miR-377-3p
To confirm whether AKT2 expression was regulated by miR-377-3p, AKT2 expression was upregulated in 22Rv1 and LNCaP cells with regulated expressions of miR-377-3p and AKT2. Our results showed that the relative protein and mRNA levels of AKT2 in 22Rv1 cells were increased compared with NC group and mimic group, decreased in AKT2+mimic group compared withAKT2 group (Fig. 6A–C, **p, ## p, ΩΩ p < 0.01). However, the relative protein and mRNA levels of AKT2 in LNCaP cells transfected with siAKT2 or miR-377-3p inhibitor demonstrated opposite results to those described above (Fig. 6D–F, Φ p < 0.05, ΔΔ p, θθ p, ΦΦ p < 0.01). Thus, miR-377-3p could target and inhibit the expression of AKT2.
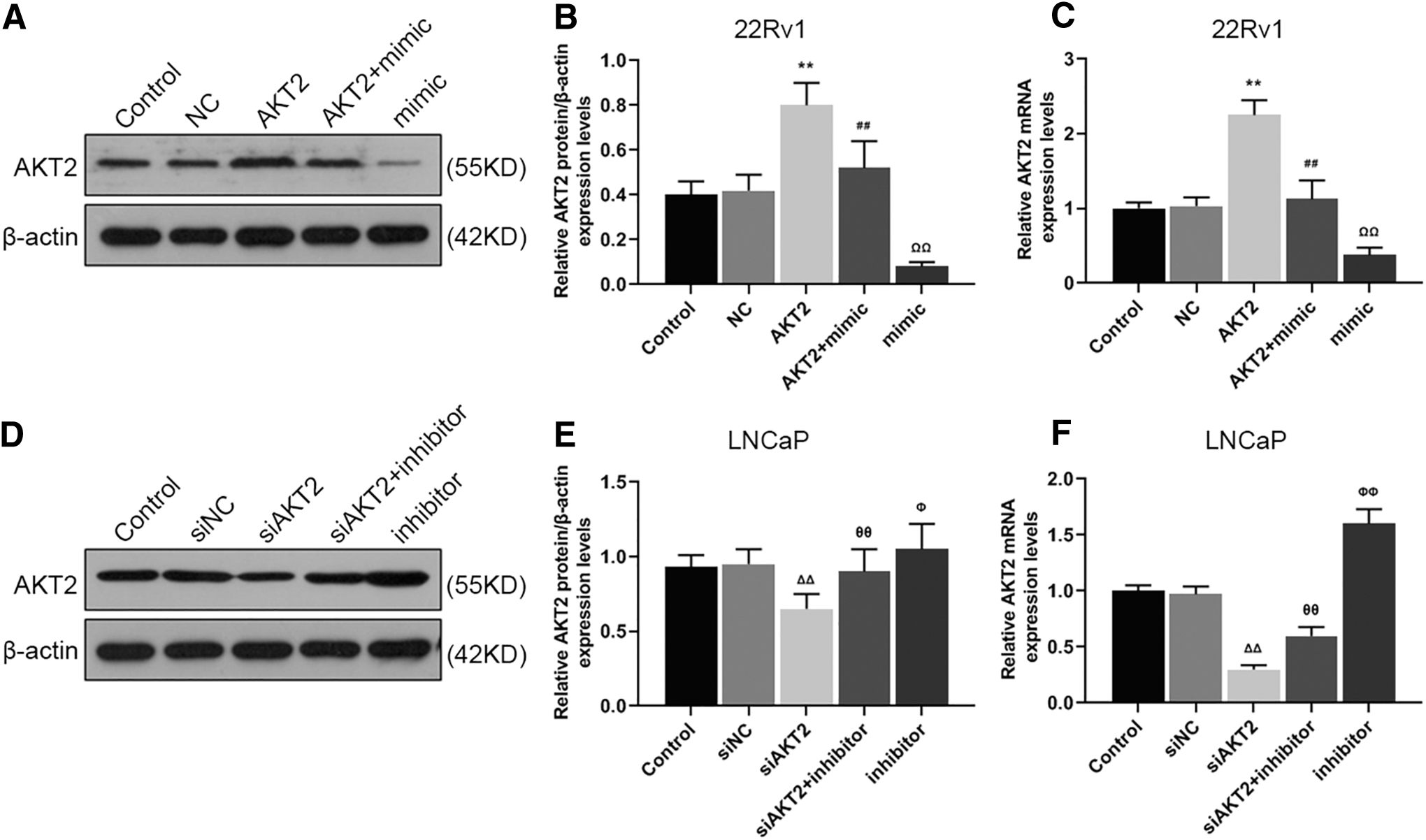
The expression of AKT2 could be regulated by miR-377-3p.
Discussion
In the present study, SNHG1 expression was found increased in PCa tissues, leading to lower overall survival rate of PCa patients. miR-377-3p was confirmed to be the target gene of SNHG1 and was proved to be negatively related to the proliferation, but positively related to the apoptosis of PCa cells. Moreover, AKT2 was proved to be the target of miR-377-3p, and was positively correlated with the proliferation, but negatively correlated with the apoptosis rate of PCa cells. Also, its function could be modulated by miR-377-3p. The SNHG1/miR-377-3p/AKT2 axis in PCa cells could be further explored to prevent and treat PCa.
In the current study, SNHG1 was overexpressed in the cancer tissues, leading to an unfavorable prognosis of PCa patients. Moreover, we detected a positive relationship between the expression of SNHG1 and PCa cell proliferation, and a negative relationship between the expression of SNHG1 and PCa cell apoptosis. Li et al. demonstrated that in PCa, SNHG1 could promote the proliferation of cells. 23 Huang et al. found that the abnormal expression of SNHG1 served as a promoter in many cancers. 24 Correspondingly, Wan, Xuechao et al. found a higher expression of SNHG1 in invasive extraprostatic tumors at pT3a, pT3b, and T4 stages than in intraprostatic tumors at pT2a, pT2b, and pT2c stages. 25 Taken together, it could be confirmed that SNHG1 overexpression could accelerate PCa progression.
Conversely, miR-377-3p was found reduced in PCa tissues, showing a negative correlation with SNHG1 expression. Furthermore, we found that miR-377-3p could be targeted by SNHG1, and the two noncoding RNAs could counteract each other's effects on the proliferation and apoptosis of PCa cells. Formosa et al. pointed that miR-377 performed its antitumorigenic effect in PCa cells through targeting FZD4, which was thought to be a mediator of epithelial/mesenchymal transition. 26 Al-Kafaji et al. detected that expression of miR-377 decreased in PCa patients as compared with healthy subjects. 27 Moreover, Shi et al. showed that miR-377-3p overexpression decreased the capacity of proliferation of gastric cancer cells, and LINC00339 overexpression could reverse such an inhibitory effect of miR-377-3p. 28 In addition, Li et al. demonstrated that miR-377-3p overexpression decreased the differentiation and the expression of adipogenic markers in adipocyte. 29 Thus, miR-377-3p might play a suppressive role in the process of PCa, and such an effect was regulated by SNHG1.
AKT activation plays a critical role in the growth and survival of cells. 30 Researches proved that Akt expression reduced cell apoptosis, which attracted research attention on the signaling elements related to Akt in cancer studies. 31 –34 As shown in the results, AKT2 overexpression increased the proliferation and reduced the apoptosis rate of PCa cells (the downregulation of AKT2 exhibited the opposite results), which was in line with the findings shown in previous studies, for instance, Le Page et al. observed that at the transcription and translational levels, AKT2 was overexpressed in PCa cell lines 35 ; Bertacchini et al. mentioned that maintaining high phosphorylation status of AKT2 would promote the migratory and invasive phenotypes of prostate epithelial cells 36 ; similarly, Chin et al. reported that loss of AKT2 in PTEN-deficient PCa would trigger significant regression of tumors. 37 Thus, AKT2 overexpression contributes to the development of PCa, however, AKT2 could be targeted and interact with miR-377-3p.
Previous studies showed that upregulation of SNHG1 was associated with metastasis and poor prognosis in cancers. 38 –41 In addition, Tian et al. pointed out that upregulation of SNHG1 was correlated with poor prognosis of acute myeloid leukemia patients. 42 These data suggested that downregulating SNHG1 level might be a novel therapeutic approach to the treatment of cancers. However, these assumptions should be comprehensively determined in the future studies. The current study found that the SNHG1 contributes to the promotion of PCa cells through regulating miR-377-3p/AKT2 axis by performing in vitro experiment, however, as lncRNA-miRNA function is not a single-strand contact, but rather a largely unknown network mode, in vivo study is required to further confirm the results. Thus, these results primarily elucidated the underlying mechanism of SNHG1 in PCa, and provided a new molecular target for the treatment of the cancer.
Conclusions
The SNHG1/miR-377-3p/AKT2 regulatory axis in PCa cells was detected. The upregulated AKT2 might be a result of the dysregulation of the interaction balance between the expressions of miR-377-3p and SNHG1. Based on such a discovery, the intervention of SNHG1/miR-377-3p/AKT2 axis could be further explored in the treatment of PCa.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
This work was supported by the Medical Health Technology Development Plan of Shandong Provincial (2016WSA06053); the Science and Technology Plan of Yantai City (2017YD004); and the Yantai Science and Technology Plan Project Returns (2017YT06000060).