
Abstract
Background:
Breast cancer is the second most common cancer in women, which is usually treated by radiation therapy. However, resistance of cancer cells to radiation therapy has made treatment difficult. Therefore, finding effective ways to reduce the radiation resistance of cancer cells is an urgent problem to be solved.
Materials and Methods:
MCF-7 and MDA-MB-231 cells (on accepting radiation) were established to model radiation resistance, namely MCF-7/R and MDA-MB-231/R. The authors then examined the expression of miR-634 through quantitative reverse transcription–polymerase chain reaction. MCF-7/R and MDA-MB-231/R cells were transfected with overexpressed miR-634 mimics. In addition, TargetScan predicted which binding site was targeted by miR-634, and luciferase assay detected the signal transducer and activator of transcription 3 (STAT3) 3′UTR luciferase activity after transfection of mimics expressing miR-634 into HEK-293 cells. 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT), flow cytometry, and western blot assays were used for examination of different levels of biological function.
Results:
miRNA-634 expression was significantly decreased in radiated MCF-7 and MDA-MB-231 cells. When miR-634 mimic was transfected into radiation-resistant MCF-7/R and MDA-MB-231/R cells, the survival rate of radiation-tolerant cells was significantly reduced. Moreover, STAT3 was found to directly interact with miR-634, and further studies demonstrated that miR-634 negatively regulated STAT3.
Conclusion:
miR-634 was able to regulate STAT3 and enhance the sensitivity of breast cancer cells to radiation; these results might shed new light on radiation therapy for breast cancer.
Introduction
Breast cancer is one of the most common malignant tumors in women and the second most common cancer condition leading to death. To date, there was no effective means to treat cancer. Traditionally, surgery resection, chemotherapy, and radiation therapy are the primary methods of breast cancer treatment. 1 Radiation therapy is an effective means of treating about 50% of cancer patients. 2 This treatment relies on accumulation of energy doses in tumor cells. After irradiating tumor cells with rays accompanied by high energy, such as gamma rays, X-rays, and high-energy ion beams, the cancer cells or their vasculature will be destroyed, thereby achieving the purpose of inducing tumor death. 3
In previous randomized trials, radiation therapy was found to significantly reduce breast cancer recurrence rates and moderately reduce breast cancer mortality. 4,5 For patients with long-term radiation therapy, the ability to reduce or even eliminate cancer cells is remarkably strengthened. 5 –7 Radiation therapy is effective for many early-stage breast cancers. However, controlling local cancer through radiation therapy remains an urgent problem to be solved. In patients receiving radiation therapy, 8%–15% of breast cancer patients still have local control failure because of the range of sensitivity of breast tumor cells to radiation. 8 Another reason for poor radiation therapy is that radiation therapy is not ideal due to the lack of predictive biomarkers for radiotherapy that can be used for routine clinical use to indicate optimal radiation doses. 9
miRNAs are small RNA molecules that act as translational inhibitors of messenger RNA through their binding to the 3′-untranslated region 10,11 and their expression has undergone profound changes in cancer and/or is strongly regulated during carcinogenesis. To date, some miRNAs have been shown to play an essential role in development, progression, metastasis, or invasion of breast cancer, such as let-7, 12 miR-30, 13 and miR-21. 14 In addition, some miRNAs such as miR-139-5p, 15 miR-22, 16 and miR-668 17 have been reported to promote antiradiation or increase sensitivity in the radiation of breast cancer. Studies have shown that miR-634 regulates the Ras-MAPK pathway in ovarian cancer, thereby increasing drug sensitivity 18 ; miR-634 increases the sensitivity of nasopharyngeal carcinoma cells to paclitaxel and inhibits cell growth in vitro and in vivo 19 ; miR-634 can regulate the mTOR pathway, reduce cervical cancer cell proliferation, and induce apoptosis 20 ; and the mitochondrial apoptosis pathway can be induced by miR-634, which enhances chemotherapy-induced cytotoxicity. 21
In this study, the authors revealed that miR-634 expression was downregulated in antiradiation breast cancer cells. The sensitivity of breast cancer cells to radiation and apoptosis of antiradiation cells were enhanced by overexpression of miR-634. At the same time, miR-634 can target and inhibit signal transducer and activator of transcription 3 (STAT3). Further studies indicated that miR-634 inhibited STAT3 expression, which was able to increase the sensitivity of breast cancer cells to radiation. These results suggest that miR-634 may be a new marker of radiotherapy resistance in breast cancer.
Materials and Methods
Antiradiation cell lines and culture
Breast cancer cells, MCF-7 and MDA-MB-231 cells, (2 × 106 cells/well) were cultured in DMEM added with 10% fetal bovine serum. Ten minutes before irradiation, the medium was replaced with 120 mL of fresh complete medium to form appropriate backscatter. The cells were irradiated with 6-MV X-rays generated by a 6-MV linear accelerator at a dose rate of 100 cGy per minute for a total dose of 1.8 Gy per dose. After the end of irradiation, the medium was changed to fresh intact medium. Subculture was carried out when the cell fusion reached about 90%. The second irradiation was performed when cell fusion reached about 50%. Establishing a radiation-tolerant cell population requires at least 24 exposures with a total dose of 43.2 Gy. A population of radiation-tolerant cells, called MCF-7/R and MDA-MB-231/R cells, was selected. Unirradiated parental cells served as control cells. Cells were irradiated with different doses of X-rays. Then, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) and quantitative reverse transcription–polymerase chain reaction (qRT-PCR) assays were used to, respectively, detect the cell viability and expression of miR-634.
Cell transfection
miR-634 mimic, inhibitor, and associative control were constructed by Hanbio Co. (Shanghai, China). MCF-7/R and MDA-MB-231/R cells were transfected with overexpressing miR-634 mimics, mimic control, miR-634 inhibitor, and inhibitor control. Transfection reagent Lipofectamine 2000 (Invitrogen) was used for cell transfection, according to the manufacturer's instructions. Then, cells were irradiated with X-rays and harvested 24 h after transfection.
Dual-luciferase reporter assay
The 3′UTR of STAT3, including the binding site of miR-634 or the mutating miR-634 seed region, was synthesized by the Beijing Genomics Institute (BGI, Beijing, China) and then subcloned into the pmirGLO vector (Promega, Madison, WI). Radiation-resistant MCF-7, HEK293, and MDA-MB-231 cells were transfected with STAT3 WT or STAT3 MUT; after this, miR-634 mimic or negative control (NC) was transfected into cells by Lipofectamine 2000 (Invitrogen). The dual-luciferase reporter assay system (Promega) was performed according to the manufacturer's protocol 48 h after transfection.
Quantitative reverse transcription–polymerase chain reaction
Total RNAs from cell lines were extracted by using TRIzol reagent (Invitrogen) according to the manufacturer's protocol. First-strand cDNA was performed using the reverse transcriptase kit (Takara, Otsu, Japan). The Bio-Rad real-time PCR machine was used to test the target gene expression using the Probe qPCR Mix (RR391S; Takara) instrument (Bio-Rad, Hercules, CA). miR-634 was normalized to that of U6, then each expression level was calculated with the 2−ΔΔCt method. The sequences of primers for STAT3, GAPDH, miR-634, cleaved caspase-3, and cleaved PARP are listed in Table 1. The experiment was performed in triplicate.
List of Primer Sequences were Used in the Study
MTT assay
During MTT assay, cells were seeded at a density of 1 × 103 cells/mL per well in 96-well plates and grew to 80% confluence. Fifteen microliters of MTT solution (5 mg/mL; Sigma, St. Louis, MO) was added and incubated for 4 h at 37°C. Next, 150 μL of DMSO was added to dissolve formazan crystals. The authors then measured the optical density at 490 nm using a microplate reader (BioTek, Winooski, VT).
Flow cytometer assay
In the apoptotic assay, the authors used a 7AAD-Annexin apoptosis kit to quantify the population of apoptotic cells (BD, Franklin Lakes). In brief, EDTA-free trypsin was used to detach targeted cells, and DMEM with serum was used to neutralize trypsin. Then, cells were washed after supplementation of flow buffer. After washing, 5 μL of Annexin V/FITC and 5 μL of propidium iodide (PI) were, respectively, added into the tube containing target cells and incubated for 15 min at room temperature in the dark. Finally, the FACSCalibur flow cytometer (BD Biosciences, CA) was used for analysis of cell apoptosis. All experiments were run in triplicate.
Western blotting
After cell collection, the authors added RIPA lysis buffer (Sigma) into the culture plate for total protein extraction, and a protein quantification kit (BCA; Beyotime Biotechnology, China) was used to extract and qualify total protein according to the manufacturer's instructions. After protein extraction, 10% SDS-PAGE and electronic transferring machine were used to separate and reload the protein onto the polyvinylidene difluoride membrane. Then, the membrane was incubated with 5% blocking buffer (10% defatted milk and 1% BSA in TBST) for 1–2 h at room temperature. Primary antibodies such as anti-STAT3 (1:5000; Abcam), anti-cleaved caspase-3 (1:1000; Abcam), and anti-cleaved PARP (1:1000; Abcam) were used to identify target protein expression.
Statistical analysis
Mean ± standard deviation was used for result interpretation. One-way analysis of variance with Bonferroni's correction was used to analyze the data for comparison of multiple groups. In addition, an independent Student's t-test was used to analyze data for comparison of two groups. GraphPad Prism 6.0 software was used for data analysis; p < 0.05 was considered as statistically significant.
Result
miR-634 expression is decreased in radiation-resistant breast cancer cells
MCF-7 and MDA-MB-231 cells were analyzed for their radiation sensitivity. After irradiation, the cellular proliferation activity was determined by MTT (WST1) assay (Fig. 1A). In this study, the authors observed the expected reduced survival capacity of MCF-7, MDA-MB-231, MCF-7/R, and MDA-MB-231/R cells. However, MCF-7 and MDA-MB-231 cells were found to be significantly decreased in comparison with MCF-7/R and MDA-MB-231/R (p < 0.01, Fig. 1A). In addition, the expression level of miR-634 was significantly decreased in MCF-7/R and MDA-MB-231/R in comparison with MCF-7 and MDA-MB-231 (p < 0.01, Fig. 1B).
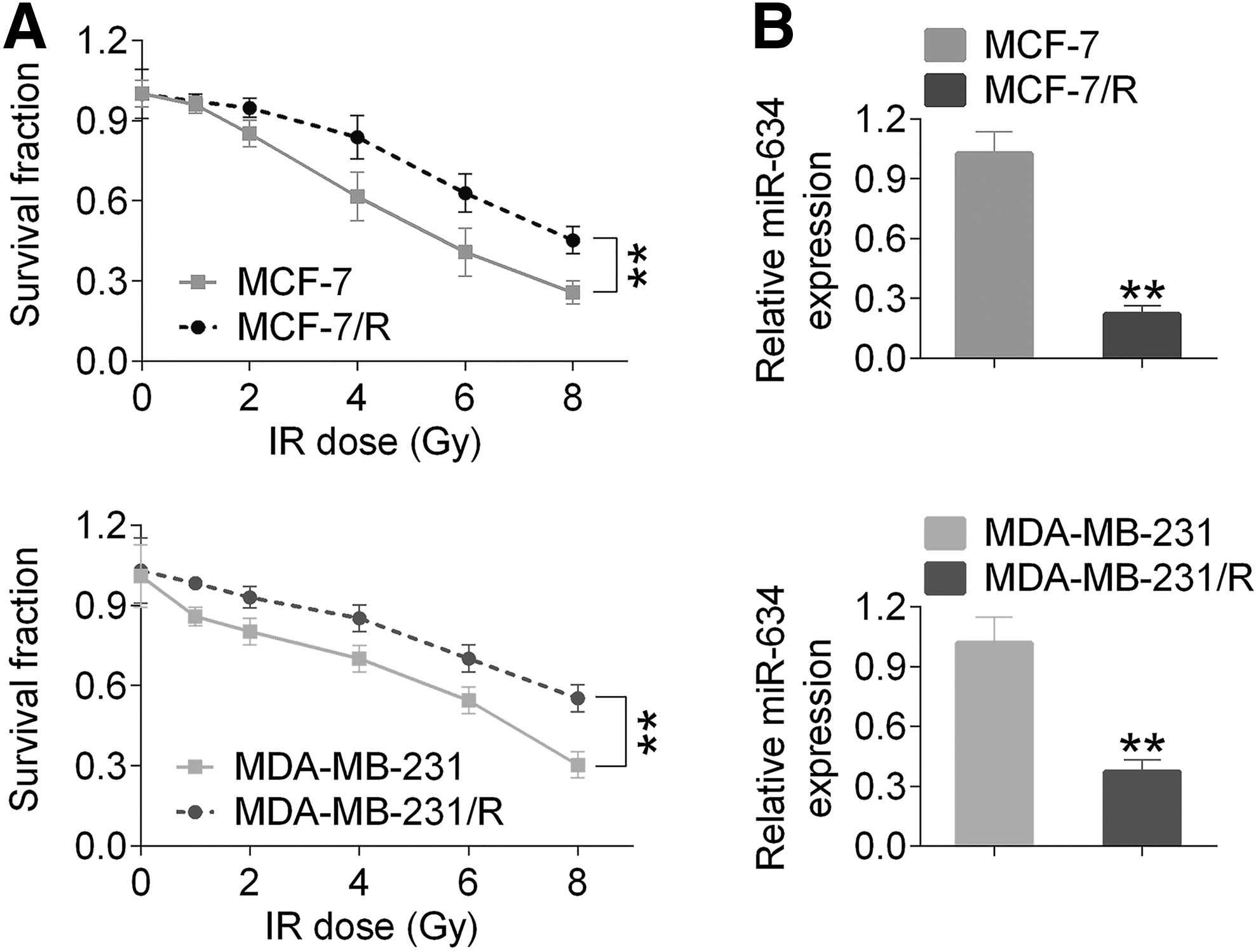
Breast cancer cell survival and expression of miR-634 in MCF-7, MDA-MB-231, MCF-7/R, and MDA-MB-231/R cells after irradiation.
Overexpression of miR-634 increases radiosensitivity of radiation-resistant cells
MCF-7/R and MDA-MB-231/R cells were transfected with miR-634 mimics and then the survival capability of cells after irradiation was detected. qRT-PCR and MTT assays were performed to examine the effect of miR-634 on postirradiation cell survival. The authors observed that miR-634 was overexpressed in MCF-7/R and MDA-MB-231/R cells in comparison with the control mimic group (p < 0.01, Fig. 2A), but the expression of miR-634 was significantly reduced in the miR-634 inhibitor group compared with the NC inhibitor group (p < 0.01, Fig. 2A). In addition, the survival curves for cells were detected after exposure to irradiation over a dose range of 0 to 8 Gy. The results showed that miR-634 significantly decreased survival of radiation-tolerant cells compared with the control mimic group in both cell lines (p < 0.01, Fig. 2B).
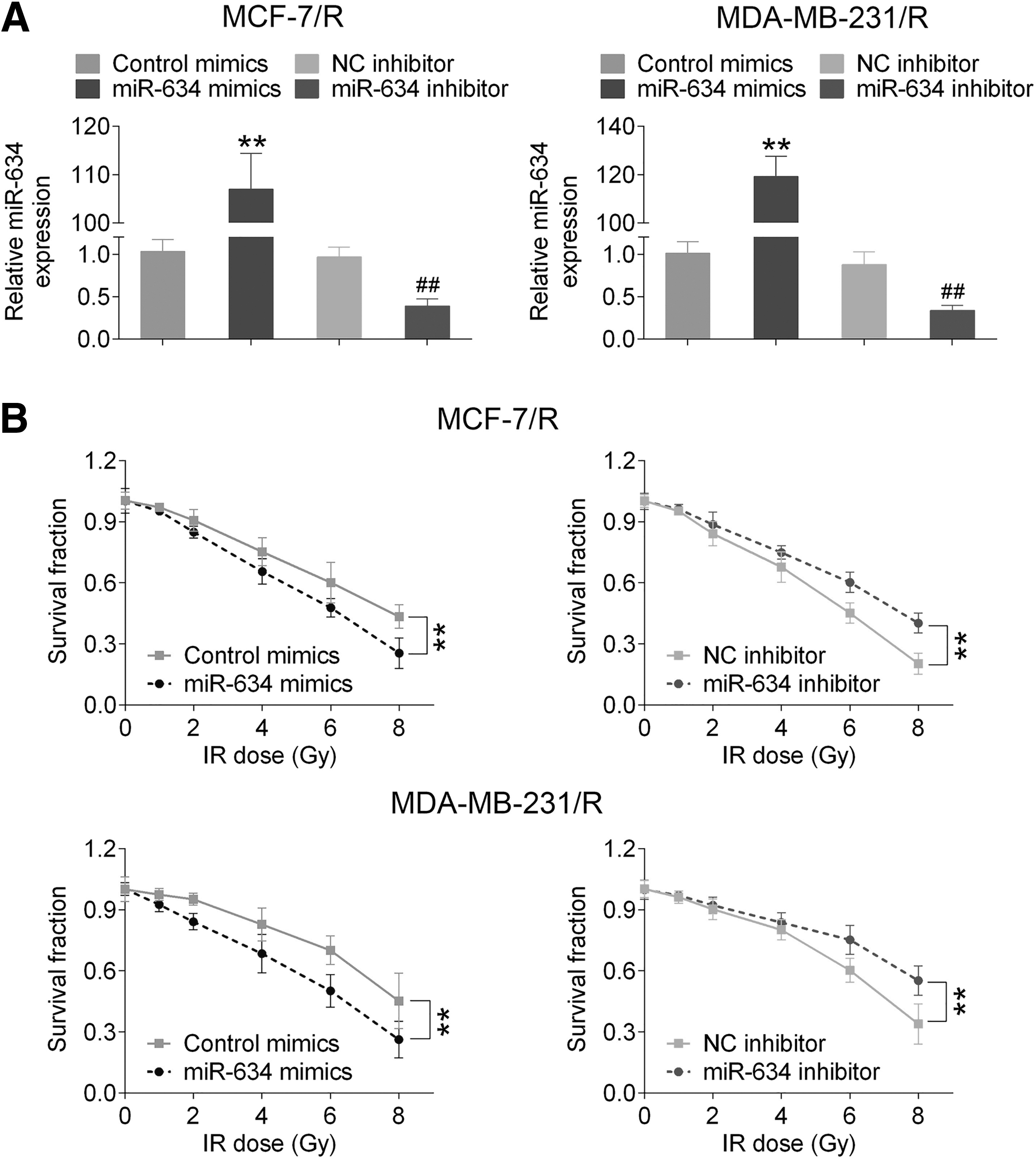
Overexpression of miR-634 enhanced radiosensitization in vitro.
Overexpressed miR-634 induces apoptosis of radiation-resistant cells
To investigate whether miR-634 could induce apoptosis of cells by enhancing radiation sensitivity of radiation-resistant cells, MCF-7/R and MDA-MB-231/R cells were transfected with miR-634 in control mimic, NC mimic, miR-634 mimic, and miR-634 inhibitor groups. After incubation for 24 h, these cells were exposed to irradiation (4 Gy) and subjected to Annexin V/PI double staining and flow cytometry analysis 24 h later. As shown in Figure 3A, the percentage of apoptotic cells in the miR-634 mimic group was significantly higher than that of control groups (p < 0.01), and the apoptotic rate was lower in the miR-634 inhibitor group compared with the NC inhibitor group (p < 0.05, p < 0.01). In addition, Western blot was used to detect the expression of cleaved caspase-3 and cleaved PARP in the miR-634 mimic group, which was obviously higher than the control group (p < 0.01, Fig. 3B), but the expression levels of these two proteins were significantly lower in the miR-634 inhibitor group compared with the control group (p < 0.01, Fig. 3B).
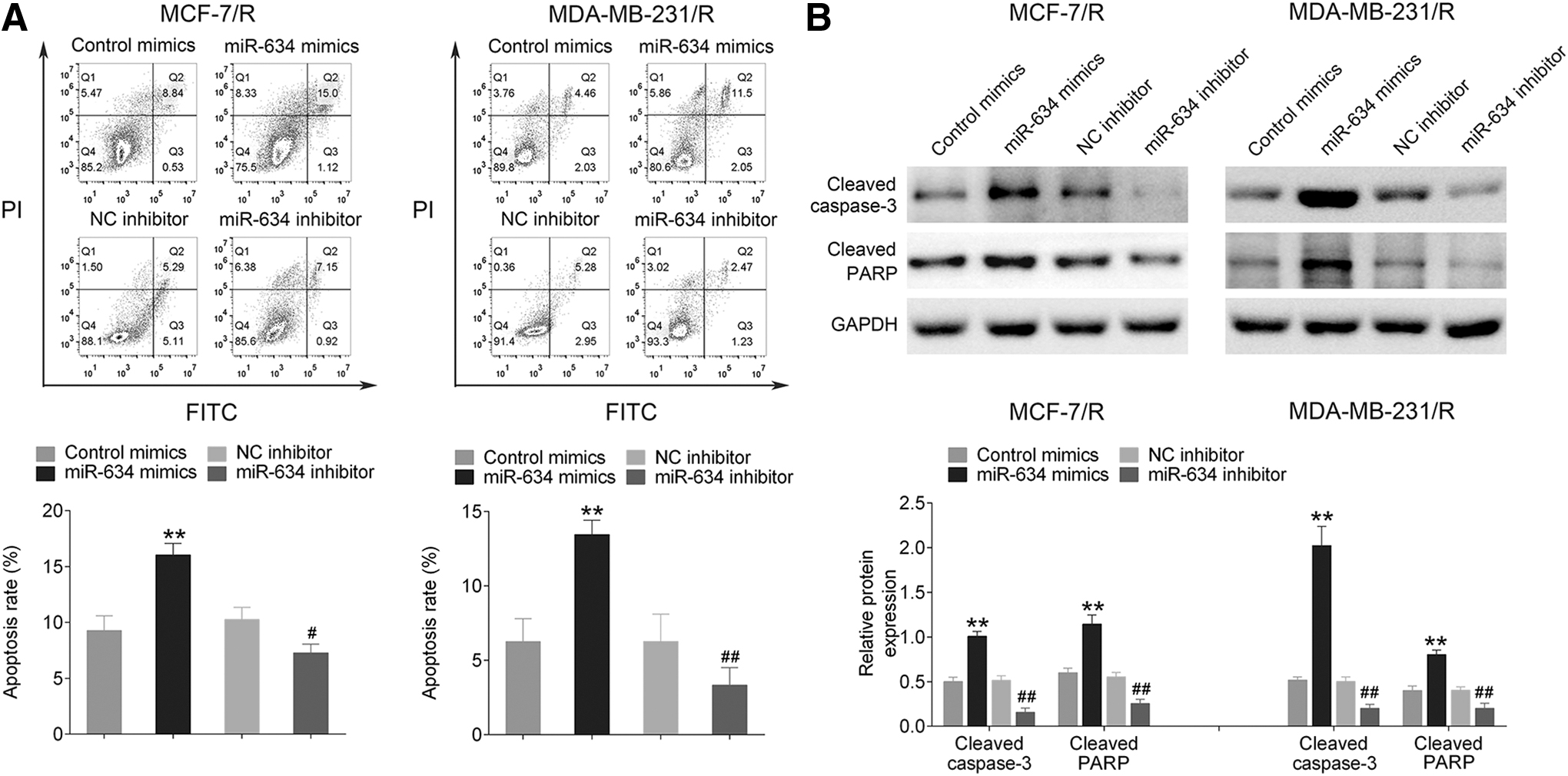
The influence of miR-634 upon MCF-7/R and MDA-MB-231/R cells.
miR-634 can target STAT3 and regulate its expression
miRNA is considered as a critical modulator that participates in action of lncRNA. To investigate the mechanism of miR-634 in MCF-7/R and MDA-MB-231/R cells, TargetScan software was used to predict the binding site of the target gene. Luciferase reporter assay was used to confirm miR-634 as a potential target miRNA of STAT3, with highest conservation (Fig. 4A). Luciferase plasmids of STAT3 wild type (STAT3 WT) contained the potential miR-634 binding sites as well as a mutated version of each site (STAT3 MUT) (Fig. 4A). The luciferase activity of cells transfected with the STAT3 WT plasmid was significantly decreased by miR-634 mimics (p < 0.01), while there was no difference in cells transfected with the STAT3 MUT plasmid (as shown Fig. 4B). These indicated that miR-634 was able to bind the STAT3 transcript. Meanwhile, as shown in Figure 4C, STAT3 was efficiently pulled down in the miR-634 mimic group (p < 0.01), and instead, expression of STAT3 was increased in the miR-634 inhibitor group (p < 0.01).
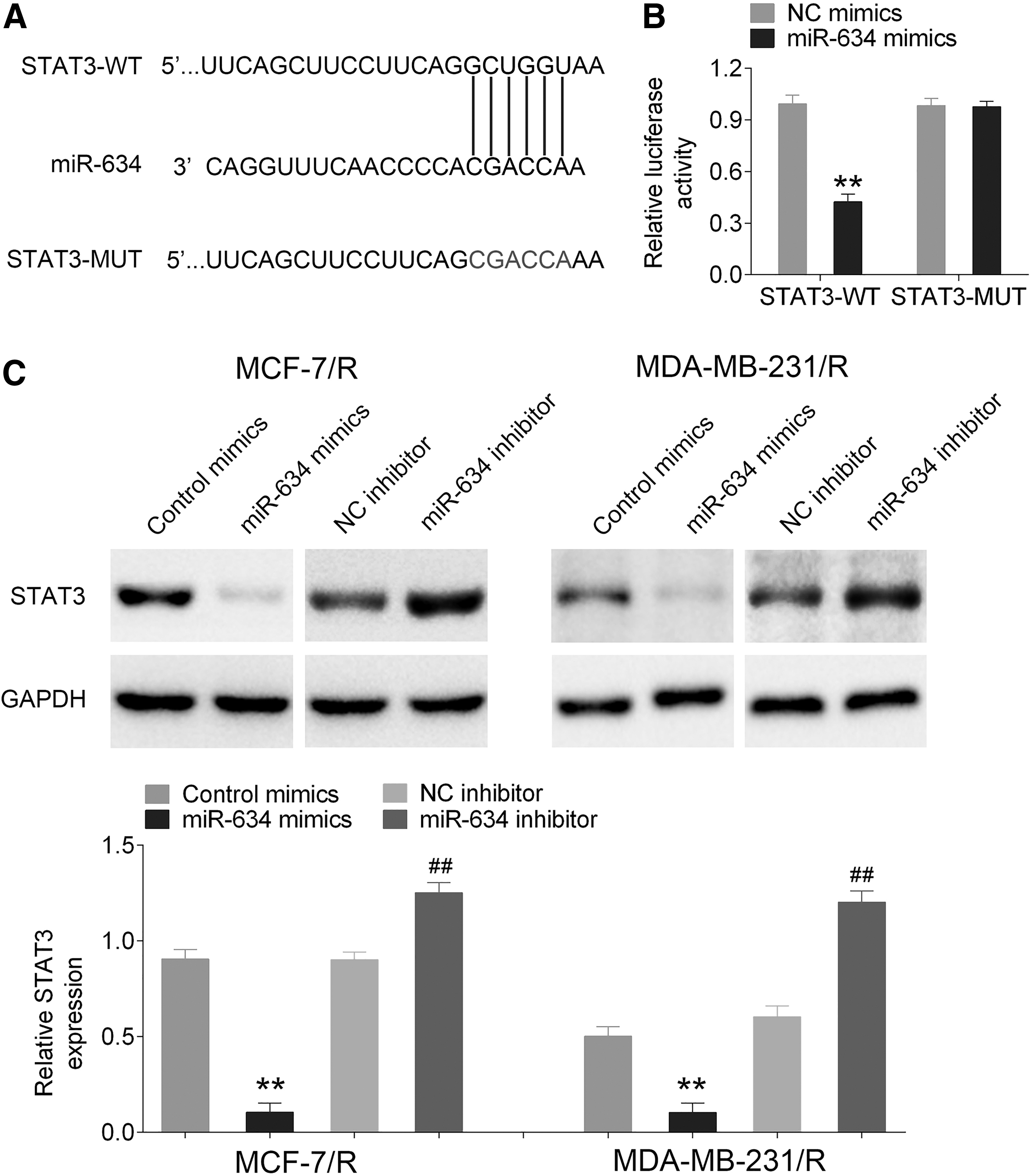
miR-634 interacted with STAT3.
STAT3 was negatively regulated by miR-634
To explore the effect of miR-634 on breast cancer cells during the regulation of STAT3, miR-634 mimics and plasmids containing overexpressing STAT3 were cotransfected into H/R-MCF-7/R cells. The experiment of qRT-PCR has been used to confirm that mRNA of STAT3 was overexpressed in H/R-MCF-7/R cells (as shown in Fig. 5A). MTT and flow cytometer assays have been used to detect survival and apoptotic rate of cells. From Figure 5B and C, the survival rate in the miR-634 mimics+vector group was significantly decreased in comparison with control mimics+vector and miR-634 mimics+STAT3 groups (p < 0.01). The apoptotic rate of H/R-MCF-7/R cells was clearly decreased in the miR-634 mimics+vector group compared with other two miR-634 mimics+STAT3 and miR-634 mimics+vector groups (p < 0.01). Western blot was utilized to investigate the expression level of STAT3, cleaved caspase-3, and cleaved PARP. As shown in Figure 5D, the expression of STAT3 was significantly upregulated in the miR-634 mimics+STAT3 group (p < 0.01), but it was downregulated in the miR-634 mimics+vector group (p < 0.01). In addition, the expression levels of cleaved caspase-3 and cleaved PARP were found to be significantly increased in the miR-634 mimics+vector group (p < 0.01), but decreased obviously in the miR-634 mimics+STAT3 group (p < 0.01).
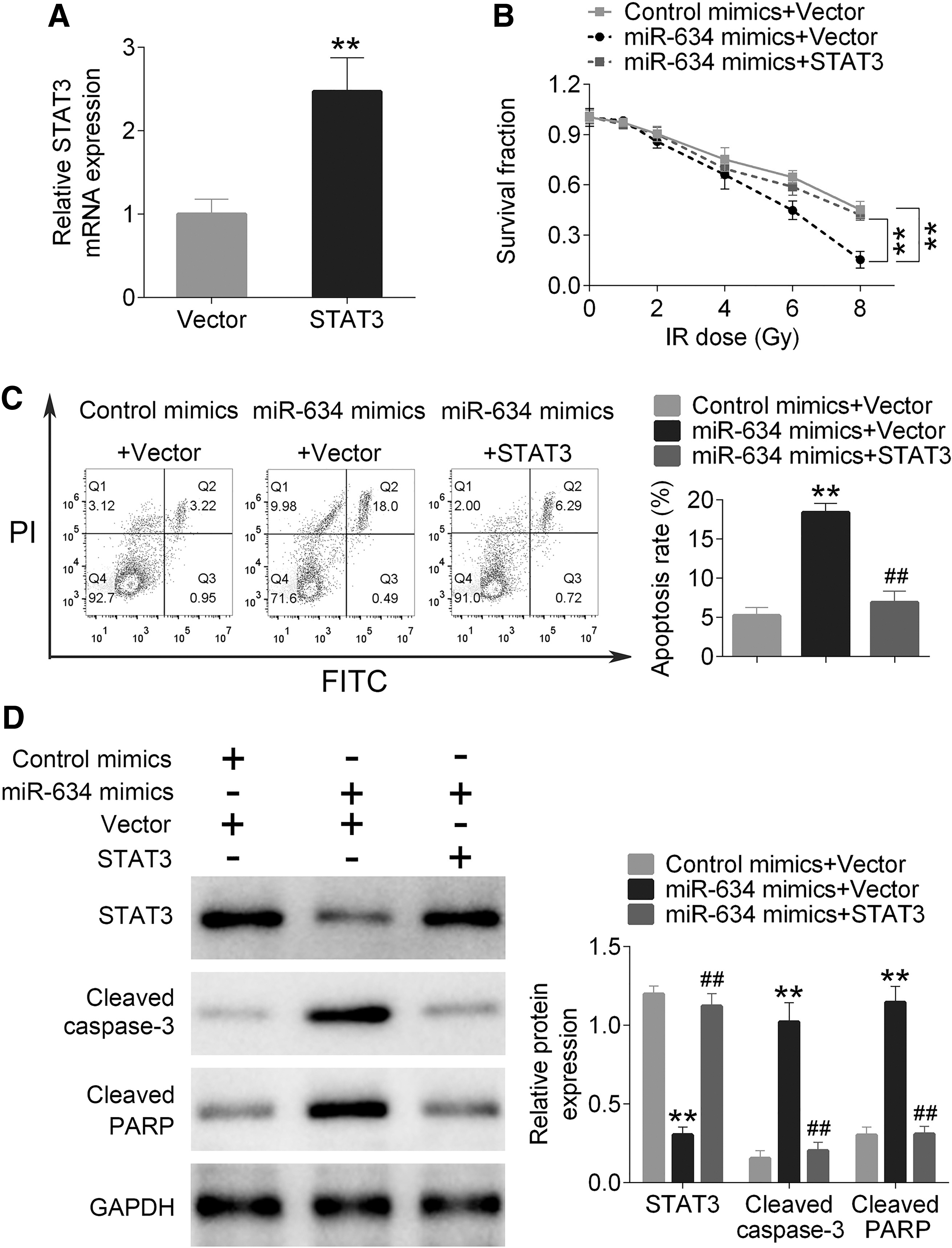
miR-634 regulating STAT3 to influence survival and the apoptosis rate.
Discussion
Although there is growing evidence that miRNAs can play a key role in cancer, the role of miRNAs in cancer radiotherapy responses remains largely unexplored. In the present study, results indicate that miR-634 is involved in the function of reducing radiation resistance in breast cancer cells by regulating STAT3. The study elaborated on the mechanism of miR-634 regulating STAT3 and it might be a new way for cancer cell radiotherapy.
In this research, the authors found that miR-634 was highly inhibited in radiation-tolerant cells, MCF-7/R and MDA-MB-231/R cell lines. To detect the influence of miR-634 on MCF-7/R and MDA-MB-231/R cells, the author's transfected the overexpressed miR-634 into these two cell lines, which was found to drastically decline in cell survivability. In addition, miR-634 was able to induce increased apoptosis of MCF-7/R and MDA-MB-231/R cells. Meanwhile, the expression levels of cleaved caspased-3 and cleaved PARP were found to be upregulated in MCF-7/R and MDA-MB-231/R cells transfected with miR-634 mimics. Further exploration indicated that miR-634 could target STAT3 and negatively regulate its expression. To investigate the effect of STAT3 on MCF-7/R and MDA-MB-231/R cells, overexpressed STAT3 and miR-634 mimics were cotransfected into cells. The survival rate increased, as well as the apoptotic rate decreased, in comparison with the miR-634 mimic group. The expression of STAT3 in the cotransfection experiment was found to be reduced, but expression of cleaved caspased-3 and cleaved PARP was found to be enhanced. From these results, the authors found that miR-634 was able to increase sensitivity of breast cancer cells to radiation. Further investigation revealed that miR-634 was able to target STAT3 and inhibit its expression. However, this behavior weakened the ability of miR-634 to increase sensitivity of breast cancer cells to radiation. This may be due to partial binding of miR-634 and STAT3, resulting in a decrease in the total amount of miR-634, thereby the trend of apoptosis was found to be slowed down.
Previous researches indicated that STATs act as important signal transduction effector proteins for cytokine or hormone-inducing pathways, controlling development, proliferation, or differentiation and homeostasis of many cell types. 22 –24 In STATs, STAT3 is more highly conserved than other members of the STAT family, can be activated by a wider range of factors and stimuli, and may regulate a wider variety of processes. These results lead to constitutive activation or overexpression of STAT3 in tumor-derived cell lines as well as many primary human cancer tissues. 25,26 In this research, the authors verified that STAT3 was the target of miR-634, and miR-634 could inhibit expression of STAT3 in breast cells. Studies have revealed that miR-634 was identified as an essential miRNA in tumor suppression, such as miR-634 was able to attenuate the growth and metastasis of liver cancer cells, 27 act as a tumor suppressor in pancreatic cancer, 28 and enhance chemotherapy-induced cytotoxicity. 21 Consistent with previous conclusions, this study showed that miR-634 was also able to suppress breast cancer cells by targeting STAT3 and increasing the sensitivity of cancer cells for radiation therapy. In addition, inhibition of STAT3 will effectively block the JAK/STAT3 pathway, which will further affect cancer cells.
In conclusion, the present findings suggested that miR-634 is downregulated in radiation-resistant breast cancer cells and serves as a tumor suppressor by directly targeting STAT3. Additionally, the results further clarified the importance of the miR-634/STAT3 molecular network in breast cancer development and may provide a novel therapeutic approach for treating breast cancer.
Footnotes
Ethical Approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Author Contribution Statement
Y.W. and B.L.Y. conceived and designed the experiments; F.K., Z.J.C., and D.S.F. analyzed and interpreted the results of the experiments; and J.L. and J.Z. performed the experiments.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.