
Abstract
Objective:
LncRNA-KAT7 is a recently identified tumor suppressor in colorectal cancer, whereas its roles in other malignancies remain unclear. This study aimed to investigate the roles of KAT7 in nonsmall cell lung cancer (NSCLC).
Results:
The results showed that KAT7 was downregulated in NSCLC and predicted poor survival. KAT7 negatively correlated with miR-10a in NSCLC. In NSCLC cells, overexpression of KAT7 led to downregulated miR-10a, whereas silencing of KAT7 led to upregulated miR-10a. Methylation-specific polymerase chain reaction revealed that KAT7 positively regulated the methylation of miR-10a. Cell proliferation assay showed that overexpression of miR-10a led to increased proliferation rate of NSCLC cells. In addition, overexpression of KAT7 played an opposite role and reduced the effects of the overexpression of miR-10a.
Conclusion:
In conclusion, KAT7 negatively regulates miR-10a through epigenetic mechanisms to participate in NSCLC cell proliferation.
Introduction
Lung cancer is the most common cancer among all malignancies for both prevalence and mortality. 1 In 2018 alone, a total of 1,761,007 patients died of lung cancer, which accounted for 18.4% of all cancer deaths. 2 In the same year, 2,093,876 patients were affected by lung cancer, accounting for 11.6% of all new cancer cases. 2 Nonsmall cell lung cancer (NSCLC) is the main subtype of lung cancer. 3 Smoking is the major risk factor for NSCLC, whereas smoking itself is not sufficient for the occurrence and development of this disease. 4 In addition, NSCLC also affects never smokers, highlighting the importance of other factors, such as genetic factors involved in this disease. 5
Previous investigations of the molecular pathogenesis of lung cancer have revealed a large number of genetic factors with critical functions in the occurrence and progression of NSCLC. 6,7 Understanding the functionality of genetic factors involved in NSCLC would provide new insights into the development of novel targeted therapies. 8,9 More and more studies have shown that, although noncoding RNAs (ncRNAs), such as long ncRNAs and miRNAs, are not involved in protein synthesis, they regulate gene expression at multiple levels to regulate biological processes. 10 KAT7 has been characterized as a tumor suppressor in colorectal cancer. 11 Based on our knowledge, the roles of KAT7 in NSCLC are unknown. Our preliminary microarray analysis revealed the downregulation of KAT7 in NSCLC and its inverse correlation with miR-10a, which is an oncogenic miRNA in NSCLC, indicating the potential interaction between KAT7 and miR-10a. This study was, therefore, performed to investigate the potential interactions between KAT7 and miR-10a in NSCLC.
Materials and Methods
NSCLC patients and tissue collection
NSCLC tissue specimens and paired fresh nontumor tissues were obtained from 60 NSCLC patients (40 men and 20 women, 39 to 68 years old, mean age 55.8 ± 7.5 years old) through biopsies, which were performed using fine needle under the guidance of MRI. These patients were admitted to the Affiliated Heping Hospital of Changzhi Medical College between May 2012 and May 2014. All patients were newly diagnosed cases and no therapies were initiated before admission. Patients complicated with other clinical disorders were excluded. This study was approved by the Ethics Committee of Affiliated Heping Hospital of Changzhi Medical College. All patients signed informed consent.
Treatment and follow-up
Different therapeutic approaches, such as surgical resection, chemotherapies, radiation therapies, and targeted therapies, were used mainly based on patients' clinical stages and health conditions. Based on the AJCC staging system, there were 12, 16, 17, and 15 cases at stages I, II, III, and IV, respectively. All patients were followed up for 5 years after admission. All patients completed follow-up.
Cell lines and cell transfections
Two human NSCLC cell lines H522 and H1703 (ATCC) were used in this study. Cells of both cell lines were cultivated in a medium composed of 10% FBS and 90% RPMI-1640 medium. Cells were cultivated under conditions of 37°C, 95% humidity, and 5% CO2. KAT7 siRNA (5′-GGGGCUGUGUAUGAUUUUGUC-3′) and negative control (NC) siRNA (5′-UGUGUCAGUCAGGCUUAGCGU-3′), as well as miR-10a mimic (5′-UACCCUGUAGAUCCGAAUUUGUG-3′) and NC miRNA (5′-AUGGCAUGUCGAUGCGGUUAGCA-3′), were synthesized by Invitrogen (Shanghai, China). The pcDNA 3.1 vector (Invitrogen) was used as backbone to construct KAT7 expression vector. H522 and H1703 cells were harvested at confluence of 75%–85%, followed by transfection of 45 nM miRNA or siRNA, or 10 nM vector into 106 cells. Untransfected cells were used as control (C) cells. Cells with NC siRNA or miRNA, or empty vector transfection were used as NC cells. Cells used in subsequent experiments were harvested at 24 h post-transfection.
RNA preparations and quantitative real-time polymerase chain reaction analysis
Total RNAs were extracted using TRIzol (Invitrogen). To remove genomic DNAs, RNA samples were digested with DNA eraser (Takara) at 37°C for 2 h. Reverse transcriptions (RTs) were performed using the Precision nanoScript2 Reverse Transcription Kit (Primerdesign), followed by quantitative polymerase chain reaction (qPCR) prepared using the SYBR Premix Ex Taq™ kit (Takara). The expression levels of KAT7 were measured using GAPDH as endogenous control. MiRNAs were extracted using the High Pure miRNA Isolation Kit (Sigma-Aldrich). The expression levels of mature miR-10a were determined using the All-in-One™ miRNA qRT-PCR Detection Kit (Genecopoeia). Primer sequences used were 5′-AGCTCTTGGTTGAGCCCTTC-3′ (forward) and 5′-GGGGCTGTGTGTGATTTTGTC-3′ (reverse) for KAT7; 5′-ACCACAGTCCATGCCATCAC-3′ (forward) and 5′-TCCACCCTGTTGCTGTA-3′ (reverse) for GAPDH. The forward primer for miR-10a was 5′-TACCCTGTAGATCCGAATTTG-3′. Universal miRNA reverse primer and U6 primers were from the kit. The 2−ΔΔcq method was used to analyze the changes of gene expression across samples. All polymerase chain reactions (PCRs) were repeated three times.
Methylation-specific PCR
Genomic DNA Isolation Kit (ab65358; Abcam) was used to extract genomic DNAs from H522 cells. The EZ DNA Methylation-Gold™ Kit (ZYMO RESEARCH) was used to convert DNAs, followed by PCRs prepared using the Taq PCR Master Mix Kit (QIAGEN) to evaluate the methylation of miR-10a gene. Primer sequences were 5′-GATCTGTCTGTCTTCTGTATAT-3′ (forward); 5′-AGAGCGGAGTGTTTATGTCAACT-3′ (reverse). The methylation-specific PCR (MSP) primer sequences were 5′-GATTTGTTTGTTTTTTGTATATATTT-3′ (forward) and 5′-AAAACGAAATATTTATATCAACT-3′ (reverse).
Cell proliferation assay
H522 and H1703 cells were collected at 24 h post-transfection to prepare single cell suspensions with a cell density of 105 cells/mL. Cells were seeded into each well of a 96-well culture plate with 0.1 mL cell suspension per well. Cells were cultivated under the aforementioned methods and CCK-8 solution (DOJINDO, Japan) was added into each well at 4 h before cell collection to reach 10% final concentration. Cells were harvested every 24 h until 96 h, OD values at 450 nM were measured using a microplate reader.
Statistical analysis
The GraphPad Prism 6 software was used to process the data from three biological replicates involved in each experiment. Paired t test was used to compare differences between two types of tissue. Differences among multiple groups were explored by one-way analysis of variance, followed by comparing differences between two groups using Tukey test. Correlations were analyzed by Pearson's correlation coefficient. With the median expression level of KAT7 as cutoff value, patients were divided into high and low level groups (n = 30). Based on the follow-up data, survival curves of both groups were plotted using k-means methods, and log-rank test was used to compare the two curves. p < 0.05 was statistically significant.
Results
KAT7 was downregulated in NSCLC and predicted poor survival
The differential expression of KAT7 in NSCLC was evaluated by measuring the expression levels of KAT7 in both NSCLC and nontumor tissues from the 60 NSCLC patients. Compared with nontumor tissues, the expression levels of KAT7 were significantly lower in NSCLC tissues (Fig. 1A, p < 0.001). Survival curves were plotted and it showed that the survival rate of patients in the high KAT7 level group was significantly higher than that of patients the in low KAT7 level group (Fig. 1B). Therefore, downregulation of KAT7 is involved in NSCLC and predicts poor survival.
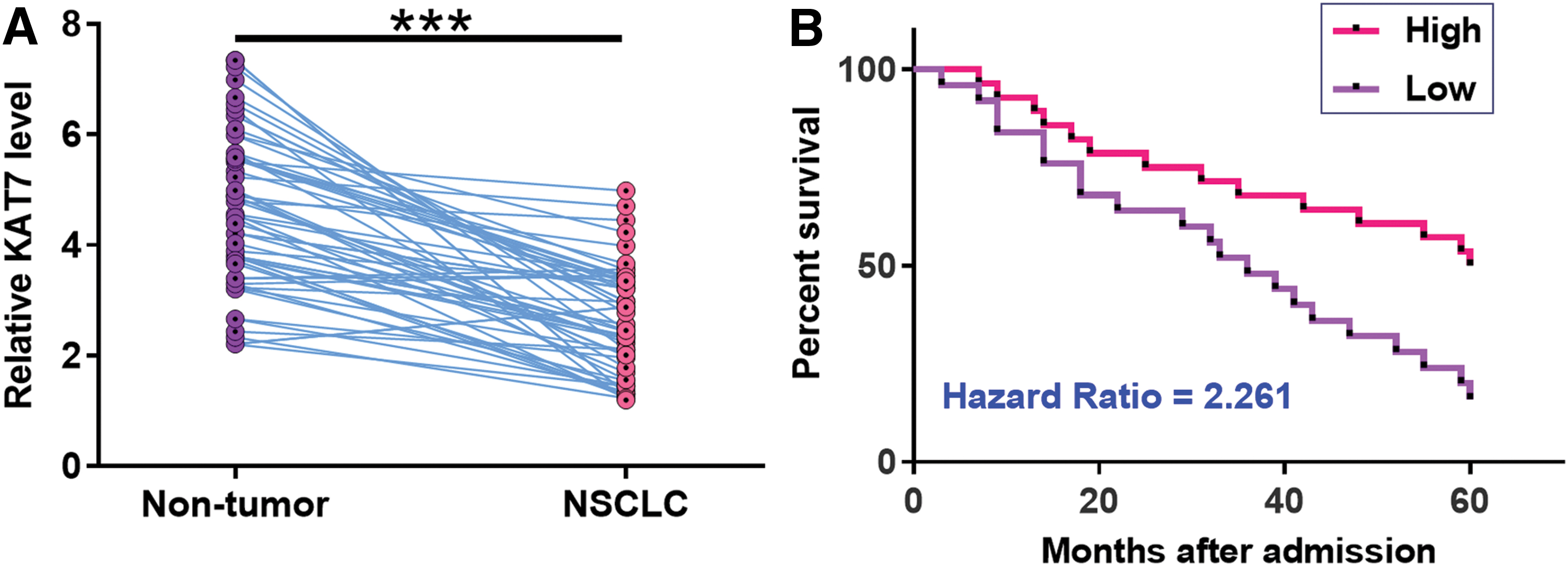
KAT7 was downregulated in NSCLC and predicted poor survival. The differential expression of KAT7 in NSCLC was evaluated by measuring the expression levels of KAT7 in both NSCLC and nontumor tissues from the 60 NSCLC patients. PCRs were repeated three times and paired t test was used to compare two groups. With the median expression level of KAT7 as cutoff value, patients were divided into high and low level groups (n = 30)
KAT7 is negatively correlated with miR-10a in NSCLC
The expression levels of miR-10a in NSCLC tissues were also measured by miR-10a. Pearson's correlation coefficient was used to analyze the correlations between expression levels of miR-10a and KAT7 across NSCLC tissues. As shown in Figure 2, a significant and inverse correlation was observed between them. Therefore, KAT7 and miR-10a may have interactions in NSCLC.

KAT7 is negatively correlated with miR-10a in NSCLC. Pearson's correlation coefficient was used to analyze the correlations between expression levels of miR-10a and KAT7 across NSCLC tissues. Color images are available online.
KAT7 negatively regulated miR-10a in H522 cells
To investigate the interactions between KAT7 and miR-10a, H522 cells were transfected with KAT7 expression vector, KAT7 siRNA or miR-10a mimic. Compared with the C and NC groups, the expression levels of KAT7 and miR-10a were significantly altered (Fig. 3A, p < 0.05). Moreover, overexpression of KAT7 led to downregulation of miR-10a, whereas silencing of KAT7 led to upregulation of miR-10a (Fig. 3B, p < 0.05). In contrast, overexpression of miR-10a did not affect the expression of KAT7 (Fig. 3C).
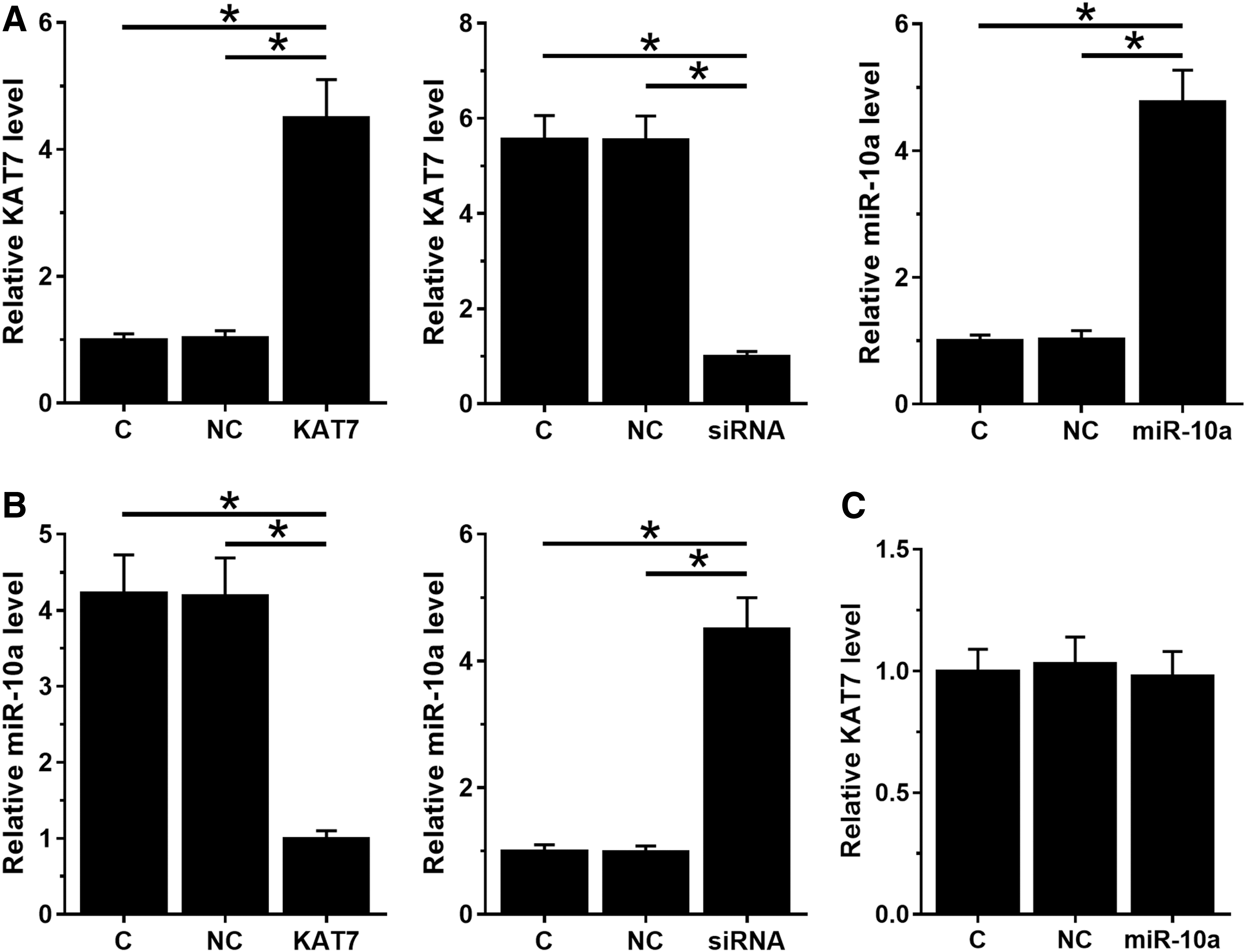
KAT7 negatively regulates miR-10a in H522 cells. To investigate the interactions between KAT7 and miR-10a, H522 cells were transfected with KAT7 expression vector, KAT7 siRNA, or miR-10a mimic. Transfections were analyzed at 24 h post-transfection
KAT7 positively regulated the methylation of miR-10a
To explore the mechanisms of the regulation of miR-10a by KAT7, MSP was performed to analyze the effects of overexpression and silencing of KAT7 on the methylation of miR-10a gene. It showed that overexpression of KAT7 led to increased methylation rate of miR-10a (Fig. 4A). In contrast, the silencing of KAT7 led to decreased methylation rate of miR-10a (Fig. 4B). These results indicated that KAT7 may downregulate miR-10a through methylation.
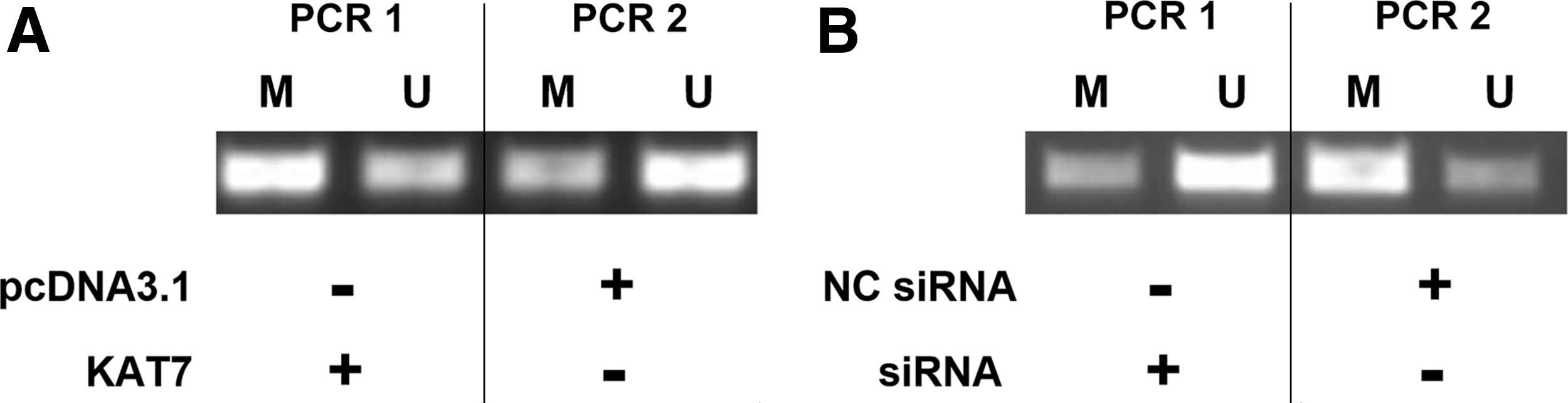
KAT7 positively regulated the methylation of miR-10a gene. MSP was performed to investigate the effects of overexpression
KAT7 suppressed the proliferation of H522 and H1703 cells through miR-10a
The effects of transfections on the proliferation of H522 and H1703 cells were assessed by CCK-8 assay. Compared with the C group, overexpression of miR-10a and silencing of KAT7 led to increased proliferation rate of NSCLC cells. Overexpression of KAT7 played an opposite role and reduced the effects of overexpressing miR-10a (Fig. 5, p < 0.05).
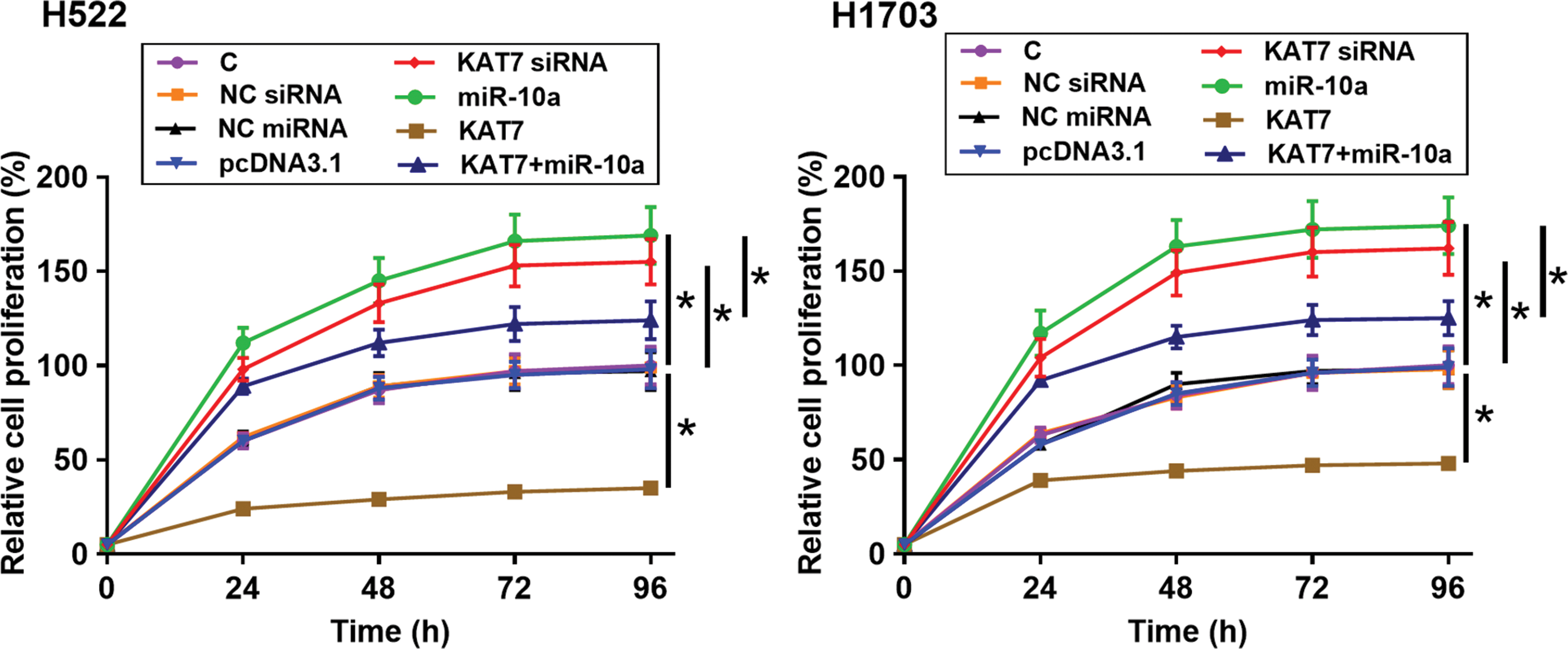
KAT7 suppressed the proliferation of H522 and H1703 cells through miR-10a. The effects of transfections on the proliferation of H522 and H1703 cells were assessed by CCK-8 assay. All experiments were repeated three times and mean values were used for comparisons. C, control cells without transfections; NC, negative control cells with empty vector; NC miRNA or NC siRNA transfection; siRNA, cells transfected with KAT7 siRNA; KAT7, cells transfected with KAT7 expression vector; miR-10a, cells transfected with miR-10a mimic; KAT7+ miR-10a, cells cotransfected with KAT7 expression vector and miR-10a mimic; *p < 0.05. Color images are available online.
Discussion
This study mainly investigated the involvement of KAT7 in NSCLC. Our findings revealed that KAT7 was downregulated in NSCLC and predicted poor survival. In addition, KAT7 may regulate the expression of miR-10a to participate in the proliferation of NSCLC cells.
The functionality of KAT7 has only been investigated in colorectal cancer. 11 It showed that KAT7 was downregulated in colorectal cancer and its overexpression led to suppressed proliferation and migration of cancer cells. 12 To the best of our knowledge, this study is the first to report the downregulation of KAT7 in NSCLC. In addition, our data showed that KAT7 negatively regulated the proliferation of NSCLC cells. Therefore, KAT7 may play tumor suppressive roles in NSCLC.
Despite the development of novel anticancer therapies, the overall survival of NSCLC patients is still poor due to the high prevalence of tumor metastasis by the time of diagnosis. 13,14 In addition, significant improvement in the early diagnosis of NSCLC is not practical owing to the lack of early diagnostic marker. 15 Therefore, development of accurate prognostic markers would be an alternative approach to improve the survival of NSCLC patients by guiding the selection of therapeutic approaches and developing individualized postoperative care system. This study showed that low expression levels of KAT7 were closely correlated with the poor survival of NSCLC. However, the reliability of KAT7 as a prognostic marker for NSCLC remains to be further evaluated by future studies with bigger sample size.
MiR-10a plays different roles in different types of cancer. 12,16 MiR-10a is downregulated in prostate cancer and targets KDM4A to suppress prostate. 16 In contrast, miR-10a is upregulated in NSCLC and targets gene of phosphate and tension homology deleted on chromsome 10 to promote cancer progression. 12 Consistently, our study also showed the enhancing effects of miR-10a on the proliferation of NSCLC, which confirmed the oncogenic functions of miR-10a in NSCLC. Interestingly, our study showed that KAT7 can positively regulate the methylation of miR-10a gene to negatively regulate the expression of miR-10a. However, the epigenetic factors involved in this process remain to be further explored.
In conclusion, KAT7 is downregulated in NSCLC and KAT7 may negatively regulate miR-10a through methylation pathways to suppress cancer cell proliferation.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this articlestudy.