
Abstract
Background:
Hepatocellular carcinoma (HCC) poses a growing threat to humans due to poor prognosis. Extract of stellera chamaejasme L. (ESC) is reported to inhibit metastasis of HCC. However, the underlying mechanism of ESC in regulating the progression of HCC needs to be further investigated.
Methods:
3-(4, 5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was used to measure cell proliferation. Flow cytometry was employed to check cell apoptosis. Transwell assay was conducted to assess the abilities of cell migration and invasion. The protein levels of proliferating cell nuclear antigen, cleaved caspase 3 (c-caspase 3), E-cadherin, janus kinase 1 (JAK1), signal transducer and activator of transcription 3 (STAT3), and phosphorylated STAT3 were detected by Western blot. The interaction between miR-134-5p and JAK1 was predicted by starBase, which was verified by the dual-luciferase reporter assay and RNA pull-down assay. The messenger RNA levels of miR-134-5p and JAK1 were determined by quantitative real-time polymerase chain reaction.
Results:
The results showed that the higher concentration or the longer time treatment of ESC led to the lower survival rate of HCC cells. Besides, ESC induced apoptosis and impeded migration and invasion of HCC cells. Moreover, downregulation of miR-134-5p inverted the effects of ESC-mediated repression on HCC progression. Further studies indicated that miR-134-5p targeted the 3′-untranslated region (3′UTR) of JAK1 and reversed JAK1-mediated impacts on HCC progression. Simultaneously, ESC inactivated JAK1/STAT3 pathway by regulating the expression of miR-134-5p.
Conclusion:
ESC suppressed HCC progression by upregulating the expression of miR-134-5p and blocking JAK1/STAT3 pathway.
Introduction
Hepatocellular carcinoma (HCC) is a typical primary liver malignancy, accounting for about 80% liver cancer worldwide, 1 and it is estimated to be the fourth most common cause of cancer mortality. 2 The morbidity and mortality of liver cancer in China exceeds that of the rest of the world. 3 Hence, it is urgent to look for new therapeutic approaches and molecular targets for HCC treatment.
Stellera chamaejasme L. is a perennial herb, universally used in traditional Chinese medicine. Asada Y et al. reported that the extract of stellera chamaejasme L. (ESC) functioned in anti-HIV. 4 Kim M et al. found that ESC induced cutaneous wound healing and anti-inflammatory activities. 5 Recent studies showed that ESC could impede the progression of various human cancers. Liu et al. confirmed that ESC induced lung cancer cell apoptosis. 6 Kan et al. reported that ESC inhibited the proliferation of HCC cells. 7 Another article showed that ESC repressed metastasis of HCC cells. 8 However, the mechanism of ESC in modulating HCC progression is still unclear and needs to be further studied.
MicroRNAs (miRNAs) are short noncoding RNAs (about 22 nucleotides) and regulate gene expression by targeting the 3′-untranslated region (3′UTR) of messenger RNA (mRNA) at the post-transcriptional level. 9 Emerging reports have deeply improved the authors' comprehension of the importance of miRNAs in human cancers. 10,11 MiR-134-5p was reported to be associated with gastric cancer, 12 cervical cancer, 13 and nasopharyngeal carcinoma. 14 Yin et al. demonstrated that miR-134-5p exerted a dramatically suppressive effect on HCC progression. 15 A new study indicated that miR-134-5p was upregulated under ESC treatment in HCC cells, 8 yet the underlying mechanism had not been completely addressed.
Janus kinase 1 (JAK1), a human tyrosine kinase protein, plays a key role in cancer therapy. 16,17 JAK1 could phosphorylate signal transducer and activator of transcription 3 (p-STAT3) to respond to cytokines and growth factors. The JAK-STAT signaling pathway was reported in lots of cancers, 18,19 including HCC. 20,21 Therefore, JAK1 may be an attractive cancer drug target and novel regulators that modulate JAK1 expression require to be defined.
In this research, the authors analyzed the impacts of ESC on proliferation, apoptosis, migration, and invasion of HCC cells. Besides, the potential mechanism of ESC in inhibiting HCC progression was further explored by subsequent experiments.
Materials and Methods
Preparation of ESC
The experiment was conducted according to a previous report. 8 In brief, the stellera chamaejasme L. was extracted using ethanol and the condensed liquid was washed on a polyamide column with 60% ethanol. After the liquid was dried under vacuum, this final compound was defined as ESC.
Cell culture
Human HCC cell lines (Hep3B and Huh-7) were purchased from the cell bank of the Chinese Academy of Sciences (Shanghai, China). Dulbecco's modified Eagle's medium (Sigma, St. Louis, MO), containing 5% CO2 and 10% fetal bovine serum (FBS) (Sigma), was utilized to culture cells.
Proliferation experiment
Cell proliferation was evaluated by 3-(4, 5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Cells were treated with various concentrations of ESC for 48 h or 20 μg/mL ESC for different time. Twenty microliters of MTT solution (5 mg/mL) (Sigma) was added to the well to incubate for 4 h. Afterward, 200 μL dimethyl sulfoxide (Sigma) was added to the well after discarding medium. Optical density values were examined at 490 nm wavelength under the microplate reader (Bio-Rad, Richmond, CA).
Flow cytometry assay
Annexin Apoptosis Detection Kit (Sigma) was hired to assess cell apoptosis following the provided protocols. Briefly, cells were resuspended in binding buffer and then 5 μL Annexin V-fluorescein isothiocyanate (Annexin V-FITC) and 5 μL propidium iodide were added to the buffer for incubating for 10 min in the dark. The stained cells were analyzed by flow cytometry (Thermo Fisher Scientific, Rockford, IL).
Transwell assay
Transwell chamber precoated with Matrigel (Corning Life Sciences, Corning, NY) or not was introduced to assess the abilities of cell invasion or migration, respectively. Transfected cells were added to the upper chamber, while the medium containing FBS was added to the lower chamber. The migrated or invaded cells were analyzed at an inverted microscope (MultiskanEX; Lab Systems, Helsinki, Finland) after being incubated for 24 h and stained with crystal violet.
Western blot analysis
Proteins from samples were isolated using radio immunoprecipitation assay (RIPA) buffer (Vazyme, Nanjing, China). Pierce™ Rapid Gold BCA Protein Assay Kit (Thermo Fisher Scientific) was used to check the concentration. Proteins were segregated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred onto polyvinylidene difluoride (PVDF) membranes (Merck Millipore, Billerica, MA). The membranes were blocked with 5% skimmed milk for 3 h. After being washed with phosphate buffered saline, the membranes were incubated with the primary antibodies against proliferating cell nuclear antigen (PCNA) (1:3000, ab18197; Abcam, Cambridge, UK), c-caspase 3 (1:1500, ab2302; Abcam), E-cadherin (1:3000, ab40772; Abcam), MMP-2 (1:2000, ab97779; Abcam), MMP-9 (1:1000, ab38898; Abcam), JAK1 (1:2000, ab47435; Abcam), STAT3 (1:3000, ab31370; Abcam), p-STAT3 (1:3000, ab76315; Abcam), or glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (1:2500, ab9485; Abcam) overnight. The secondary antibody Goat Anti-Rabbit IgG H&L (HRP) (1:3000, ab205718; Abcam) was added for incubating for 3 h after being rewashed again. The protein bands were visualized using the ChemiDoc™ MP Imaging System (Bio-Rad) after being treated with ECL kit (Vazyme).
RNA isolation and quantitative real-time polymerase chain reaction
RNAiso Plus kit (Takara, Dalian, China) was introduced to extract RNA from cells according to the provided steps and then RNA was reversed to cDNA by PrimeScript™ RT Master Mix kit (Takara). The quantitative real-time polymerase chain reaction (qRT-PCR) was conducted by Fast SYBR™ Green Master Mix kit (Vazyme). The data were analyzed by using 2−ΔΔCt method. Beta-actin (β-actin) and U6 were introduced as the endogenous controls. Primers in this study were as follows:
miR-134-5p (forward 5′-ACAATGCGCTGACCTAGCTGTA-3′, reverse 5′-CAGTCCACCAAGCAGTCACAG-3′); JAK1 (forward 5′-CGCTCTGGGAAATCTGCT-3′, reverse, 5′-TGATGGCTCGGAAGAAAGG-3′); β-actin (forward 5′-GCACCACACCTTCTACAATG-3′, reverse, 5′-TGCTTGCTGATCCACATCTG-3′); and U6 (forward, 5′-TCCGGGTGATGCTTTTCCTAG-3′, reverse, 5′-CGCTTCACGAATTTGCGTGTCAT-3′).
Cell transfection
The miR-134-5p mimic (miR-134-5p) and miR-134-5p nhibitor (anti-miR-134-5p), as well as corresponding controls (miR-NC and anti-miR-NC), were obtained from GenePharma (Shanghai, China). JAK1 expression plasmid (JAK1) and its control (Vector) were acquired from RiboBio (Guangzhou, China). Transfection experiments were performed by utilizing Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA) following the given procedures.
Dual-luciferase reporter assay
The potential target sites of miR-134-5p and JAK1 were predicted by starBase. 22 The sequences of wild-type JAK1 (JAK1 3′UTR-WT) and its mutant (JAK1 3′UTR-MUT) harboring putative target sites of miR-134-5p were cloned and inserted into pGL3 vectors (Promega, Madison, WI). Then the vectors with miR-134-5p or miR-NC were cotransfected into Huh-7 and Hep3B cells using Lipofectamine 2000 (Invitrogen). The luciferase activity was measured by the Dual-Glo® Luciferase Assay System (Promega).
RNA pull-down assay
Biotin-labeled probe against miR-134-5p (Bio-miR-134-5p) and its negative control (Bio-NC) were obtained from Sangon (Shanghai, China). After being incubated with cell lysates, the probes were captured by streptavidin-coupled beads (Sangon). Proteinase K was used to digest proteins and the enrichment of JAK1 was checked by qRT-PCR.
Statistical analysis
Experimental data were presented by mean ± standard deviation. Two independent groups were compared by Student's t-test. For more than two groups, the one-way analysis of variance (ANOVA) followed by Turkey's test was used to assess the difference. Each experiment was conducted at least three times independently. p < 0.05 represented statistical significance.
Results
ESC inhibited proliferation, migration, and invasion, and induced apoptosis of HCC cells
To explore the function of ESC in HCC progression, the authors first checked the cytotoxicity of ESC on HCC cells. The results showed that the cell survival rate gradually decreased with the increasing ESC concentrations and a notable downtrend was observed at 20 μg/mL ESC (Fig. 1A, B). These data indicated that 20 μg/mL ESC exerted inhibition effect on cell viability and the cell survival rate was ∼80%, and 20 μg/mL ESC exhibited a low cytotoxicity in HCC cells. Thus, they chose 20 μg/mL ESC for the subsequent experiments. They then investigated the effect of ESC on HCC cells under different treatment time, and the result indicated that cell survival rate was significantly declined under the treatment of 24, 48, and 72 h compared with control (Fig. 1C, p < 0.05). Apoptosis assay showed that ESC conspicuously promoted apoptosis of HCC cells (Fig. 1D, p < 0.05). In addition, transwell assay elucidated that ESC repressed migration and invasion of HCC cells (Fig. 1E, F, p < 0.05). Next, the protein levels of PCNA (related to proliferation), c-caspase 3 (related to apoptosis), and E-cadherin (related to metastasis) were measured and the data illustrated that ESC markedly downregulated the expression of PCNA and upregulated the levels of c-caspase 3 and E-cadherin (Fig. 1G–I, p < 0.05). These data revealed that 20 μg/mL ESC significantly inhibited the expression of MMP-2 and MMP-9 in both Hep3B and Huh-7 cells, suggesting that ESC inhibited cells migration and invasion by downregulating MMP-2 and MMP-9 expression (Supplementary Fig. S1A, B, p < 0.05). Together, these results suggested that ESC could impede the progression of HCC cells in vitro.
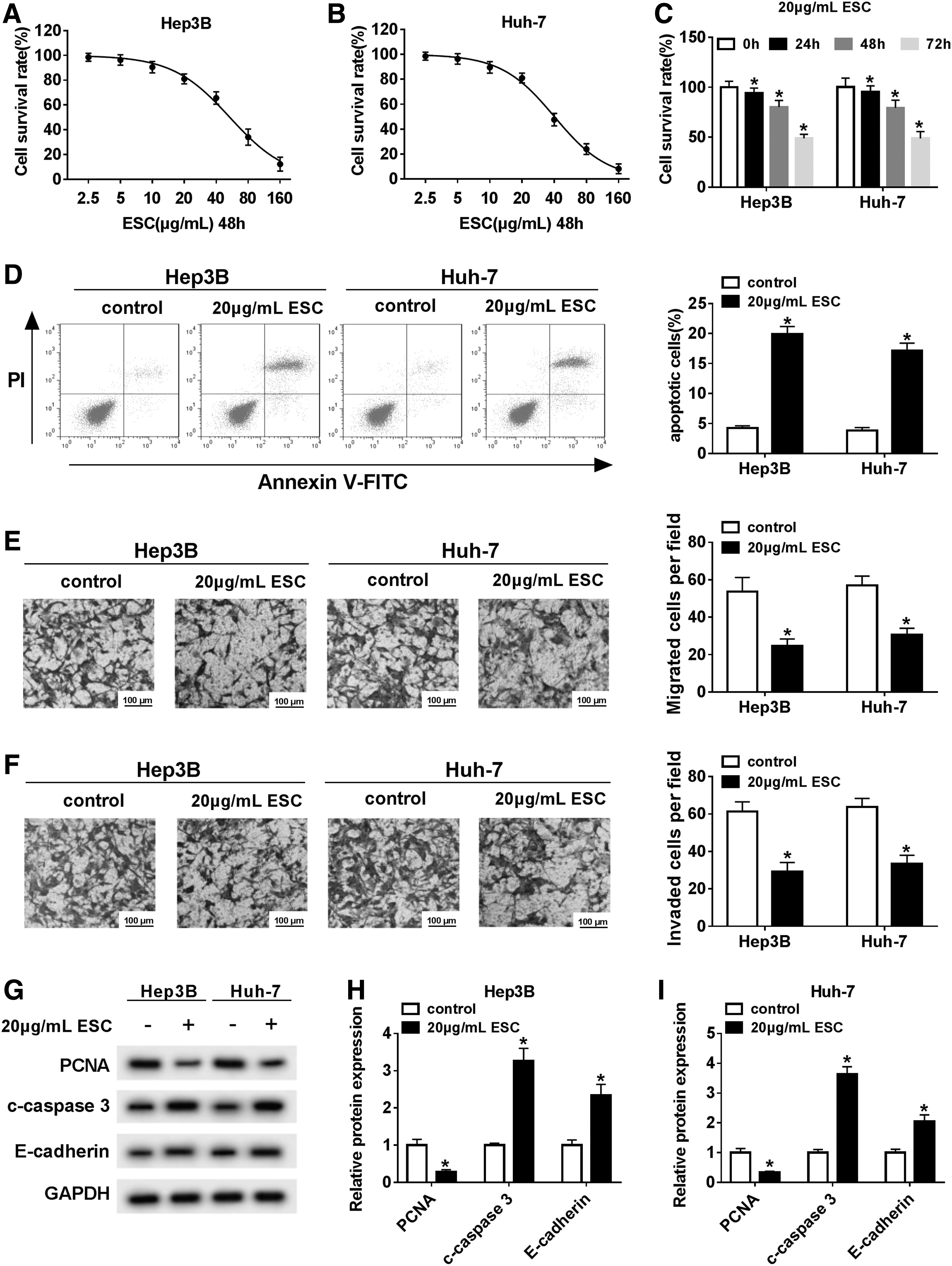
ESC repressed proliferation, migration, and invasion, and promoted apoptosis.
ESC elevated the expression of miR-134-5p in HCC cells
A previous report indicated that miR-134-5p was highly expressed under ESC treatment. 8 To investigate the relationship between ESC and miR-134-5p, the authors checked the expression of miR-134-5p in HCC cells treated with ESC. The results showed that the level of miR-134-5p continued to increase with increasing ESC concentrations (Fig. 2A, B, p < 0.05). Similarly, miR-134-5p was upregulated under treatment of ESC with different time (24, 48, and 72 h) compared with control (Fig. 2C, D, p < 0.05). All in all, these results manifested that ESC enhanced the expression of miR-134-5p in HCC cells in vitro.
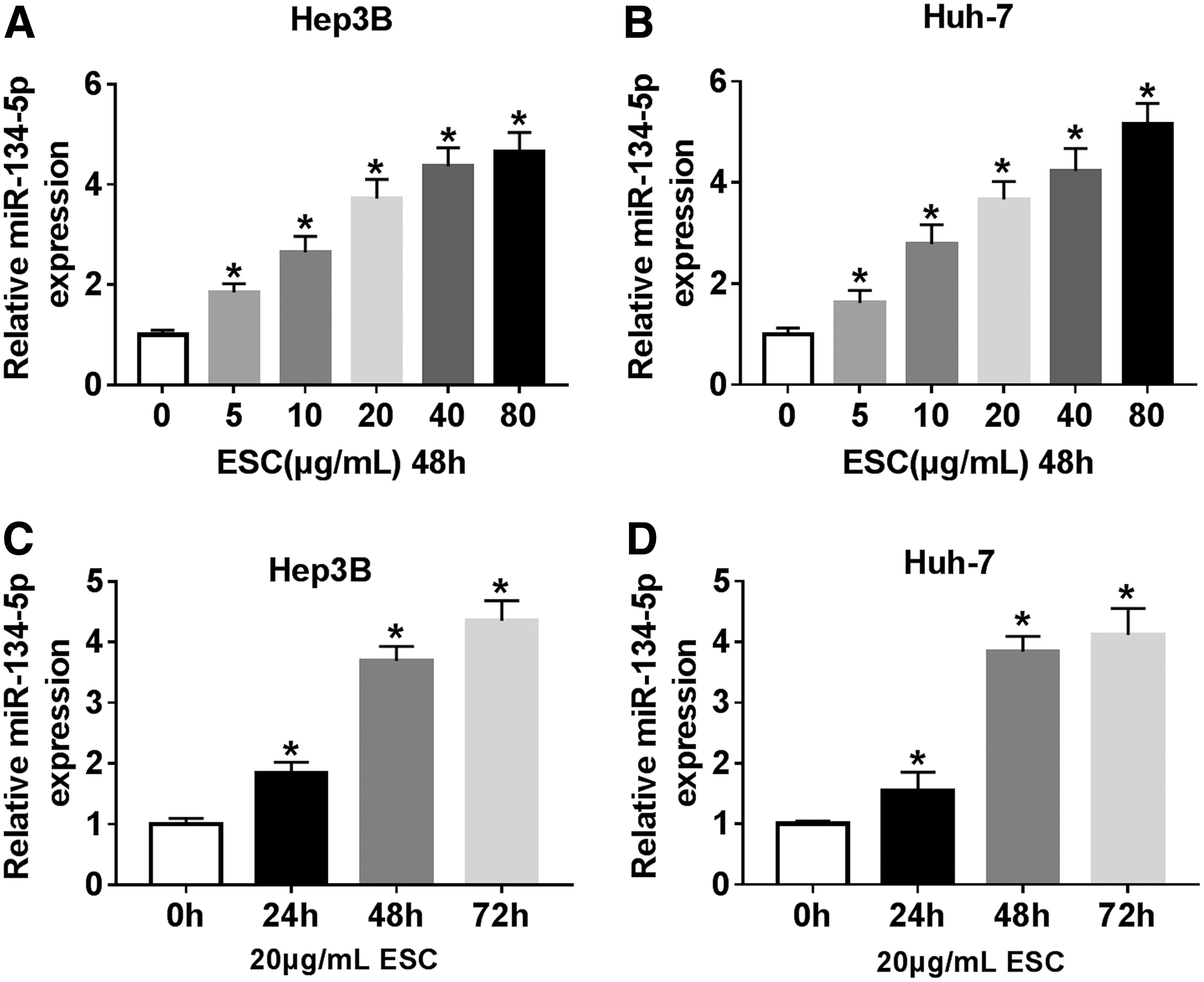
MiR-134-5p was upregulated under ESC treatment.
ESC repressed HCC progression by regulating the expression of miR-134-5p
To further study the relationship of the two in HCC, the authors evaluated the level of miR-134-5p in HCC cells treated with ESC, anti-miR-134-5p, or ESC+anti-miR-134-5p, as well as the corresponding controls. The data showed that miR-134-5p was remarkedly upregulated in ESC group, whereas the expression was significantly reduced in anti-miR-134-5p and ESC+anti-miR-134-5p group (Fig. 3A, p < 0.05). MTT assay indicated that downregulation of miR-134-5p transposed the effect of ESC-mediated inhibition on proliferation of HCC cells (Fig. 3B, C, p < 0.05). Besides, the effect of ESC-mediated promotion on apoptosis was reversed by downregulating miR-134-5p (Fig. 3D, p < 0.05). Furthermore, miR-134-5p inhibitor rescued the inhibitory effect of ESC on migration and invasion of HCC cells (Fig. 3E, F, p < 0.05). Meanwhile, the expression levels of three proteins (PCNA, c-caspase 3, and E-cadherin) in the ESC group were reversed following the transfection with anti-miR-134-5p (Fig. 3G–J, p < 0.05). From these results, it could be concluded that ESC suppressed proliferation, migration, and invasion, and induced apoptosis of HCC cells by modulating the expression of miR-134-5p in vitro.

MiR-134-5p inhibitor reversed ESC-mediated effects on HCC progression.
MiR-134-5p targeted the 3′UTR of JAK1 and negatively regulated the expression of JAK1
To figure out the potential mechanism of miR-134-5p in HCC, starBase was hired to find the possible target genes of miR-134-5p. The result showed that miR-134-5p could bind to the 3′UTR of JAK1 (Fig. 4A). To verify the prediction, corresponding reporter plasmids (JAK1 3′UTR-WT and JAK1 3′UTR-MUT) were constructed and data showed that miR-134-5p apparently diminished the luciferase activity of JAK1 3′UTR-WT in HCC cells, rather than JAK1 3′UTR-MUT (Fig. 4B, C, p < 0.05). Simultaneously, RNA pull-down assay confirmed that miR-134-5p was able to precipitate JAK1 in Hep3B and Huh-7 cells (Fig. 4D, E, p < 0.05). Further research indicated that upregulation of miR-134-5p significantly reduced the mRNA and protein levels of JAK1 in HCC cells, whereas downregulation of miR-134-5p obviously increased the expression of JAK1 (Fig. 4F–I, p < 0.05). Taken together, these results illustrated that miR-134-5p could target the 3′UTR of JAK1 and negatively modulated the mRNA and protein levels of JAK1 in vitro.
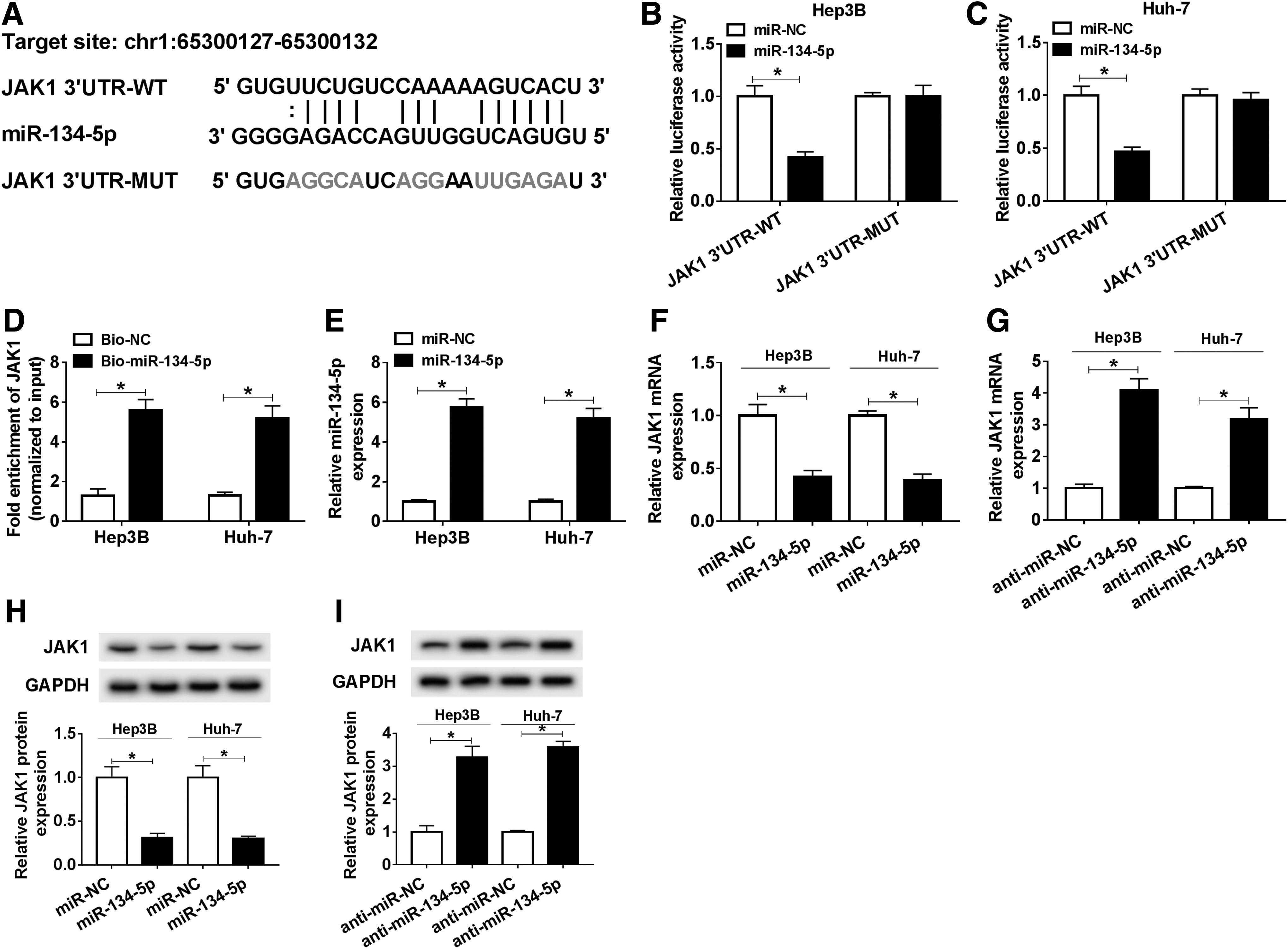
miR-134-5p targeted the 3′UTR of JAK1 in HCC cells.
Upregulation of miR-134-5p reversed the impact of JAK1-mediated promotion on HCC progression
To understand the role of the interaction between miR-134-5p and JAK1 in HCC progression, the authors checked the expression of JAK1 in HCC cells transfected with JAK1 or JAK1+miR-134-5p, as well as the matched controls. The data showed that JAK1 was evidently upregulated in JAK1 group, while the expression level of JAK1 was clearly overturned after the transfection with miR-134-5p (Fig. 5A–C, p < 0.05). In addition, elevated expression of miR-134-5p rescued the effect of JAK1-mediated promotion on proliferation of HCC cells (Fig. 5D, E, p < 0.05). Apoptosis assay elucidated that miR-134-5p mimic revoked the inhibitory effect of JAK1 on apoptosis of HCC cells (Fig. 5F, p < 0.05). Moreover, upregulating the expression of miR-134-5p abolished the effects of JAK1-mediated enhancement on migration and invasion of HCC cells (Fig. 5G, H, p < 0.05). Afterward, the protein levels of PCNA, c-caspase 3, and E-cadherin were checked in transfected HCC cells and the results disclosed that the expression of these proteins in JAK1 group was inverted after the transfection with miR-134-5p (Fig. 5I–L, p < 0.05). To sum up, these results demonstrated that miR-134-5p inhibited cell proliferation, migration, invasion and promoted cell apoptosis by regulating JAK1 in HCC cells.

Upregulation of miR-134-5p inverted JAK1-mediated effects on proliferation, apoptosis, migration, and invasion of HCC cells.
ESC repressed HCC progression by regulating the expression of JAK1
To further investigate the role of JAK1 signaling in ESC-treated HCC cells, Hep3B and Huh-7 cells were treated with ESC and transfected with JAK1 or Vector. As shown in Supplementary Figure S2A and B, ESC significantly inhibited the expression of JAK1, which was restored by JAK1 overexpression (p < 0.05). MTT assay suggested that JAK1 overexpression reversed the inhibitory effect of ESC on cell proliferation in both Hep3B and Huh-7 cells (Supplementary Fig. S2D, E, p < 0.05). Moreover, the promotion effect of ESC on cell apoptosis was blocked by upregulating JAK1 (Supplementary Fig. S2F, p < 0.05). ESC significantly suppressed cell migration and invasion, which were hindered by JAK1 upregulation (Supplementary Fig. 2G, H, p < 0.05). Besides, the inhibition on PCNA expression and promotion on c-capase-3 and E-cadherin expression of ESC were reversed by overexpression of JAK1 in both Hep3B and Huh-7 cells (Supplementary Fig. S2I, J, p < 0.05). These findings indicated that ESC inhibited HCC progression by downregulating JAK1 expression.
ESC modulated JAK1/STAT3 pathway through regulating miR-134-5p
JAK1 is a core member of the JAK1-STAT3 pathway. To explore whether ESC could affect JAK1/STAT3 pathway, the relative expression of JAK1 and p-STAT3/STAT3 in treated HCC cells was measured. The results showed that the expression levels of JAK1 and p-STAT3/STAT3 conspicuously declined under the treatment with ESC, whereas the effect was reversed by downregulating miR-134-5p (Fig. 6A, B, p < 0.05). Collectively, these results explicated a fact that ESC could inactivate JAK1/STAT3 pathway by upregulating miR-134-5p.
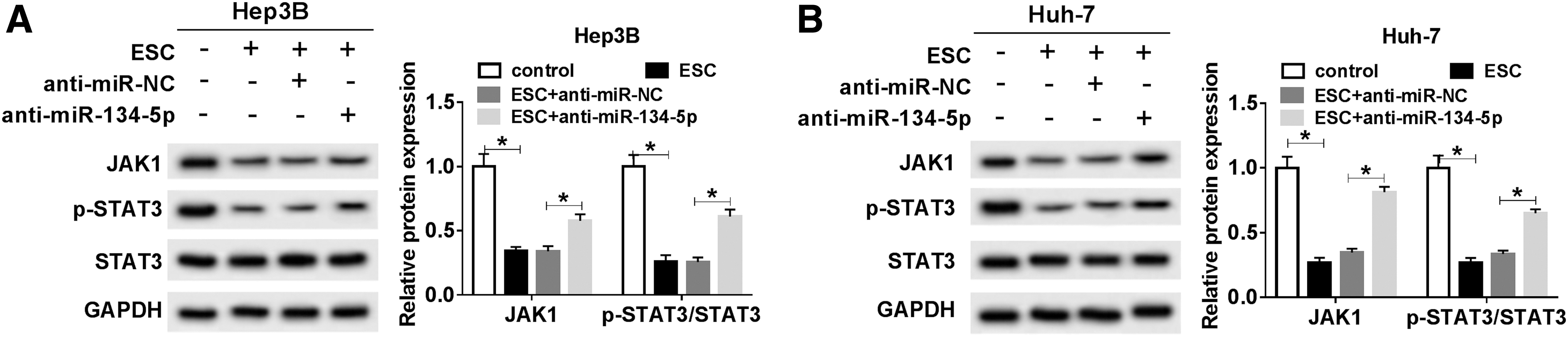
ESC reduced the expression of proteins associated with JAK1/STAT3 pathway by miR-134-5p.
Discussion
HCC is a growing threat to humans and is responsible for cancer-associated deaths worldwide. 23 Therefore, it is essential to find more anti-HCC drugs and investigate the molecular mechanisms behind them, which will contribute to the development of more effective treatments for HCC.
ESC is widely utilized in traditional Chinese medicine and confirmed to possess antibacterial, 24 antivirus, 25 and antitumors activities. 6,7 To explore the effects of ESC on HCC progression, the authors first detected its cytotoxicity to HCC cells and found that ESC inhibited cell proliferation in a dose- and time-dependent manner, and cell survival rate had a sharp downtrend at 20 μg/mL ESC. They then chose this concentration for the next experiments. Apoptosis assay and transwell assay indicated that ESC significantly promoted apoptosis and repressed migration and invasion of HCC cells, respectively. Meanwhile, ESC notably decreased the protein level of PCNA and increased the levels of c-caspase 3 and E-cadherin. These results suggested that ESC indeed hindered the development of HCC cells in vitro.
Abnormal expression of miRNA was often observed in many cancers. 26,27 Recently, an article reported that the expression of many miRNAs was changed under ESC treatment in HCC. 8 Hence, the level of miR-134-5p in HCC cells treated with ESC was determined and the data illustrated that the expression of miR-134-5p was positively correlated with ESC concentration or treatment time. That miR-134-5p was upregulated under ESC treatment was in line with Liu's report. 8 Next, the function of miR-134-5p in HCC progression was studied and the results manifested that downregulation of miR-134-5p promoted proliferation, migration, and invasion, and inhibited apoptosis of HCC cells. Similarly, the protein levels of PCNA, c-caspase 3, and E-cadherin in HCC cells treated with ESC were also reversed after the transfection with anti-miR-134-5p. Together, these results elucidated that ESC damaged HCC progression by upregulating miR-134-5p in vitro.
To further explore the underlying mechanism of miR-134-5p in HCC progression, bioinformatics analysis was performed to find its possible target genes. The data showed JAK1 was a target of miR-134-5p and the interaction was proved by dual-luciferase reporter assay and RNA pull-down assay. Also, miR-134-5p negatively modulated the expression of JAK1 in HCC cells. In addition, overexpression of JAK1 boosted proliferation, migration, and invasion, and hampered apoptosis of HCC cells, whereas elevating the expression of miR-134-5p inverted these impacts. Growing evidence has emphasized the fact that JAK1/STAT3 pathway plays a pivotal role in human cancers. Chen et al. found that HOXA10 deteriorated gastric cancer through activating JAK1/STAT3 pathway. 28 Zhou et al. reported that CtBPs promoted the malignancy of osteosarcoma cells through JAK1/STAT3 pathway. 29 From the results above, the authors knew that ESC regulated the expression of miR-134-5p and miR-134-5p could bind to the 3′UTR of JAK1, which inspired the curiosity to investigate whether ESC and miR-134-5p were involved in JAK1-STAT3 pathway. The protein levels of JAK1, STAT3, and p-STAT3 in treated HCC cells were measured and the results indicated that ESC strikingly weakened the levels of JAK1 and p-STAT3/STAT3, while downregulation of miR-134-5p reversed the effects. In summary, ESC upregulated miR-134-5p that interacted with and downregulated JAK1, thereby reducing the level of p-STAT3/STAT3, ultimately leading to the inactivation of JAK1-STAT3 pathway.
Conclusions
In conclusion, this research demonstrated that ESC could inhibit the progression of HCC by upregulating miR-134-5p and inactivating miR-134-5p-mediated JAK1-STAT3 pathway. This new mechanism may facilitate the development of novel therapeutic approaches for HCC.
Footnotes
Authors' Contributions
X.Z. and X.H. conceptualized the study. Y.C. and X.H. collected the data. F.L. and H.Y. analyzed the data. All the authors revised and approved the final version.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was approved by Special fund for medical and health development on inheriting and developing of Traditional Chinese Medicine in 2018 (Guangdong Traditional Chinese Medicine Office [2018] No.5).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.