
Abstract
The article entitled, “lncRNA IGF2-AS Promotes Cell Proliferation, Migration, and Invasion of Gastric Cancer by Modulating miR-937/EZH2 Axis,” by Zizi Li, Zhanyu Li, Zhijuan Zhong, Jianhui Zhou, Shenhao Huang, Wenying Zhou, and Jianfeng Xu (Cancer Biother Radiopharm epub 25 May 2020; Doi: 10.1089/cbr.2019.3275) is being officially retracted from the literature.
The Editor-in-Chief of Cancer Biotherapy and Radiopharmaceuticals (CBR) received an email from the corresponding author of the article, Wenying Zhou, on September 7, 2020, indicating:
“…this manuscript should no longer be published [because of] information from our superior and also after discussion with my research team, we want to repeat our experiment and then revise the [manuscript]. Our team has a lot of controversy about these results. In order to ensure the quality of the data and not to mislead readers, we decided to [retract] the manuscript. We will be more rigorous in our further study.”
The editor requested further information about precisely which data could not be reproduced, to which Dr. Wenying Zhou responded:
“…we found that the results of Figure 4B and 4C, Figure 5A and 5B could not be repeated. Our repeated results of Figure 4B and 4C were as follow[s]: compared with miR-NC group, there was no significant decrease of the luciferase activity in miR-937 group when cells co-transfected with EZH2 3′-UTR-WT.
Our repeated results of Figure 5A and 5B were as follow[s]: miR-937 inhibited the proliferation of MNK-28 and SGC-7901 cells, but the cell proliferation had no significant change in miR-937+EZH2 group compared with miR-937+pcDNA group
.
These findings resulted in the uncertainty of the targeted relationship between miR-937 and EZH2. We also repeated these experiments in different environments, but they all contradict the results in the original data. So, the conclusion is very controversial. In a responsible attitude to the readers, we decide to [retract] the manuscript
.”
The authors have extended their apologies to the Editor and to the readers of CBR.
Though in the author's original email, the request was made to “withdraw” the paper, CBR has decided to fully retract it due to irreproducible data, as the journal is committed to preserving the scientific literature and the community it serves.
Introduction
Gastric cancer (GC) is one of the most common malignancies in the world. In China, GC is highly prevalent and there are around 380,000 new cases diagnosed annually, accounting for more than 40% of the cancer incidence worldwide. 1 –3 In spite of the large developments in the treatment and diagnosis, the outcome of advanced stage patients remains rather poor. 4,5 About 50%–80% of GCs have the high risk of metastasis and recurrence after creative resection. 6,7 Thus, it is fundamental to investigate the molecular mechanisms underlying GC progression to discover prognosis biomarkers and therapeutic targets to predict patient survival.
Long noncoding RNAs (lncRNAs) are a class of noncoding RNA molecules that are longer than 200 nucleotides in length. Recently, emerging studies have reported that lncRNAs are dysfunctional in diverse tumors and abnormal lncRNAs expression is implicated in the modulation of diverse biological processes in cancers, such as tumorigenesis, proliferation, invasion, metastasis, and so on. 8,9
In GC, some lncRNAs have been revealed to function as tumor suppressors or oncogenes, thereby affecting cancer progression and prognosis. 10 For examples, lncRNA ZEB1-AS1 was associated with unfavorable prognosis in GC 11 ; lncRNA SNHG5 regulated KLF4 expression by sponging microRNA (miR)-32 to inhibit cell proliferation and migration in GC. 12 lncRNA insulin growth factor 2 antisense (IGF2-AS) was first identified in Wilms' tumor. 13 Soon afterward, an increasing amount of research has shown the important regulatory functions of IGF2-AS in cell proliferation, invasion, metastasis, metabolism, apoptosis, prognosis, and outcome in various diseases, 14 –16 indicating the potential regulatory roles in cancer development.
In this study, we took the IGF2-AS as a research target to highlight the potential role of IGF2-AS in GC. We found that IGF2-AS was elevated in GC tissues and cell lines and highly expressed IGF2-AS predicted worse prognosis. In addition, IGF2-AS knockdown inhibited cell proliferation, invasion, and migration in GC. Besides that, we first identified that upregulated IGF2-AS promoted cell progression in GC through miR-937/enhancer of zeste homolog 2 (EZH2) axis, providing a novel idea for the prognosis and treatment of GC.
Materials and Methods
Patients and specimens
The study was approved by the Ethics Committee of The Fifth Affiliated Hospital, Sun Yat-sen University and informed consent was obtained from all patients. Tumor specimens (7 cases of grade I, 6 cases of grade II, 9 cases of grade III, and 9 cases of grade IV) and adjacent normal tissues from 31 surgical patients with GC were collected from November 2012 to January 2014 at The Fifth Affiliated Hospital, Sun Yat-sen University.
The clinical features, including age, gender, TNM stages, lymph node metastasis, differentiation, and tumor size, were collected from the recruited cases. Follow-up was conducted regularly every 3 months in the first 2 years after surgery, and it was reduced to once every 6 months from the third year onward. The last follow-up was carried out in January 2019. All specimens were diagnosed by 2 independent pathologists and did not accept any treatment before surgery. Samples were immediately stored at −80°C until they were used.
Cell culture and transfection
Normal gastric epithelium cell line (GES-1) and human GC cell lines (MKN-28 and SGC-7901) were obtained from Shanghai Academy of life Sciences (Shanghai, China) and maintained in RPMI-1640 (Gibco, Carlsbad, CA) harbored with 10% fetal bovine serum (FBS) (Gibco) with 5% CO2 at 37°C.
Small interfering RNA (siRNA) targeting IGF2-AS (si-IGF2-AS), siRNA negative control (si-NC), short hairpin RNA (sh-RNA) against IGF2-AS (sh-IGF2-AS) or sh-RNA negative control (sh-NC), pcDNA or pcDNA-IGF2-AS overexpression vector (IGF2-AS), or pcDNA-EZH2 overexpression vector (EZH2) were synthesized by Genepharma (Shanghai, China). The miRNA mimic or inhibitor targeting miR-937 (miR-937 or in-miR-937) and their corresponding negative control (miR-NC or in-miR-NC) were obtained from RIBOBIO (Guangzhou, China). All oligonucleotides or vectors were transfected into MKN-28 and SGC-7901 cells by Lipofectamine TM2000 reagent (Invitrogen, Thermo Scientific, Waltham), respectively. Subsequently, cells were harvested after 48 h and transfected for further experiments.
Quantitative real-time polymerase chain reaction
TRIzol reagent (Invitrogen) was utilized to isolate RNA from tissues and cells by following the manufacturer's protocol. For IGF2-AS and EZH2 detection, complementary DNA (cDNA) was generated by PrimeScript RT Reagent (TaKaRa, Dalian, China). As to the miR-937 levels, RNA was reverse transcribed to cDNA by using miScript Reverse Transcription kit (TaKaRa). Subsequently, SYBR Premix Ex Taq (TaKaRa) was used to conduct a quantitative polymerase chain reaction (PCR). The relative expression was calculated by the 2−ΔΔCt method and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
The primer sequences were listed as follows: IGF2-AS F, 5′-CCTTGGACTTTGAGTCAAATT-3′; R, 5′-GGTCGTGCCAATTACATTTCA-3′; GAPDH F, 5′-TGCACCACCAACTGCTTAGC-3′, R, 5′-GGCATGCACTGTGGTCATGAG-3′; miR-937 F, 5′-CGGTGAGTAGGTGGGTGGG-3′, R, 5′-GTGCAGGGTCCGAGGT-3′; EZH2 F, 5′-TGCAGTTGCTTCAGTACCCATAAT-3′, R, 5′-ATCCCCGTGTACTTTCCCATCATAAT-3′.
Chromogenic in situ hybridization
Detection of IGF2-AS was carried out by chromogenic in situ hybridization (CISH) using the RNAscope® Singleplex Target Probe and the RNAscope 2.0 HD Detection kit (Advanced Cell Diagnostics, Newark, CA) according to the manufacturer's protocol. The staining intensity and the proportion of IGF2-AS-positive cells were measured in 31 GC tumor tissues and matched normal tissues. The sample images were visualized by a light microscope at 100 × magnification. CISH staining was scored by 2 independent observers using a three-tiered scoring system as follows: 0, no staining; 1, a few dots observed in a few tumor cells; and 2, >10 dots homogeneously observed in tumor cells. For each GC case, the mean score of the 10 tissue cores was evaluated and the case was classified as “IGF2-AS-negative or with low expression” when the mean score was ≤1, and as “IGF2-AS-positive or overexpressed” when the mean score was >1, respectively.
Cell counting kit-8 assay
Transfection cells were seeded into 96-well plates at a concentration of 3000 per well and cultured overnight at 37°C. After that, each well was incubated with 10 μL cell counting kit-8 (CCK-8) solution (Beyotime, Shanghai, China) at 37°C for another 4 h. Finally, the absorbance value was measured by using a microplate reader at 450 nm.
Transwell assay
For migration assay, cells suspended with serum-free RPMI-1640 medium were seeded on the upper chamber. As for invasion assay, suspended cells were placed on the upper chamber that was precoated with Matrigel (BD Biosciences, San Jose, CA). Then, the lower chamber was filled with 500 μL RPMI-1640 mixed with 10% FBS. After incubation for 24 h at 37°C, migrated or invaded cells were fixed and stained. Finally, cells in five randomly selected fields were counted with a microscope.
Dual-luciferase reporter assay
The WT or MUT IGF2-AS/EZH2 containing the putative target sequences of miR-937 were cloned into the pmiR-RB-Report (Promega, Madison, WI) to construct IGF2-AS-WT or -MUT and EZH2-WT or -MUT luciferase reporter vector. Then, MKN-28 and SGC-7901 cells were co-transfected with constructed luciferase reporters and miR-937 mimics or mimic control by using Lipofectamine 2000 for 48 h. Finally, a dual-luciferase assay kit (Promega) was used to analyze luciferase activities.
RNA immunoprecipitation
miR-937 mimics or miR-NC was transfected into MKN-28 and SGC-7901 cells. After transfection, cells were lysed by RNA immunoprecipitation (RIP) buffer (Millipore, Billerica, MA) and then the lysis was interacted with magnetic beads conjugated with anti-Ago2 (Millipore) or IgG antibody (Abcam, Cambridge, MA). Finally, the enrichment of IGF2-AS was measured by using quantitative real-time PCR (qRT-PCR).
Western blot
Western blot assay was carried out as described earlier. 17 Immunoblot assays were performed by using primary antibodies against EZH2 (1:500, ab191080; Abcam) and β-actin (1:1000, ab8226; Abcam) at 4°C overnight, followed by interaction with horseradish peroxidase-conjugated secondary antibodies (1:1000; ab9482; Abcam) at 37°C for 1 h. The protein signals were observed by an enhanced chemiluminescence method.
Murine xenograft assay
BALB/c nude mice (male, 4–6 weeks old, n = 6) were used to establish the xenograft models. SGC-7901 transfected with Lenti-sh-RNA-IGF2-AS or Lenti-sh-NC were subcutaneously injected into BALB/c nude mice. Subsequently, tumor volume was detected every 7 d. After 35 d, all mice were sacrificed and tumor masses were weighted and harvested for further study. The study has obtained the permission of the Animal Research Committee of The Fifth Affiliated Hospital, Sun Yat-sen University.
Statistical analysis
Data from three independent experiments were expressed as the mean ± standard deviation. Survival curves were estimated by using the Kaplan–Meier method and assessed by using the log-rank test. Spearman's correlation analysis was used to detect the correlation analysis. Statistically significant differences were calculated by using student's t test or one-way analysis of variance (ANOVA) as appropriate. Statistical analyses were performed by GraphPad Prism 7.0 (GraphPad, Inc., San Diego, CA). p < 0.05 indicated statistical significance.
Results
IGF2-AS is elevated in GC tissues and cell lines and high IGF2-AS expression is correlated with poor prognosis
The expression of IGF2-AS was evaluated; results showed that IGF2-AS was greatly elevated in all GC tissues compared with that of the adjacent normal tissues (Fig. 1A), and the expression of IGF2-AS in the high-grade GC tissues (grade III–IV) was significantly higher than that in the low-grade GC tissues (grade I–II) (Fig. 1B). Similarly, CISH analysis revealed that the expression of IGF2-AS was also notably increased in the GC tissue group compared with the normal group (Fig. 1C). In addition, the correlation between IGF2-AS expression and clinicopathological features in 31 GC patients was analyzed (Table 1). The results showed that IGF2-AS expression was closely correlated to TNM stages, lymph node metastasis, and tumor size whereas it was not associated with age, gender, and differentiation in GC patients.
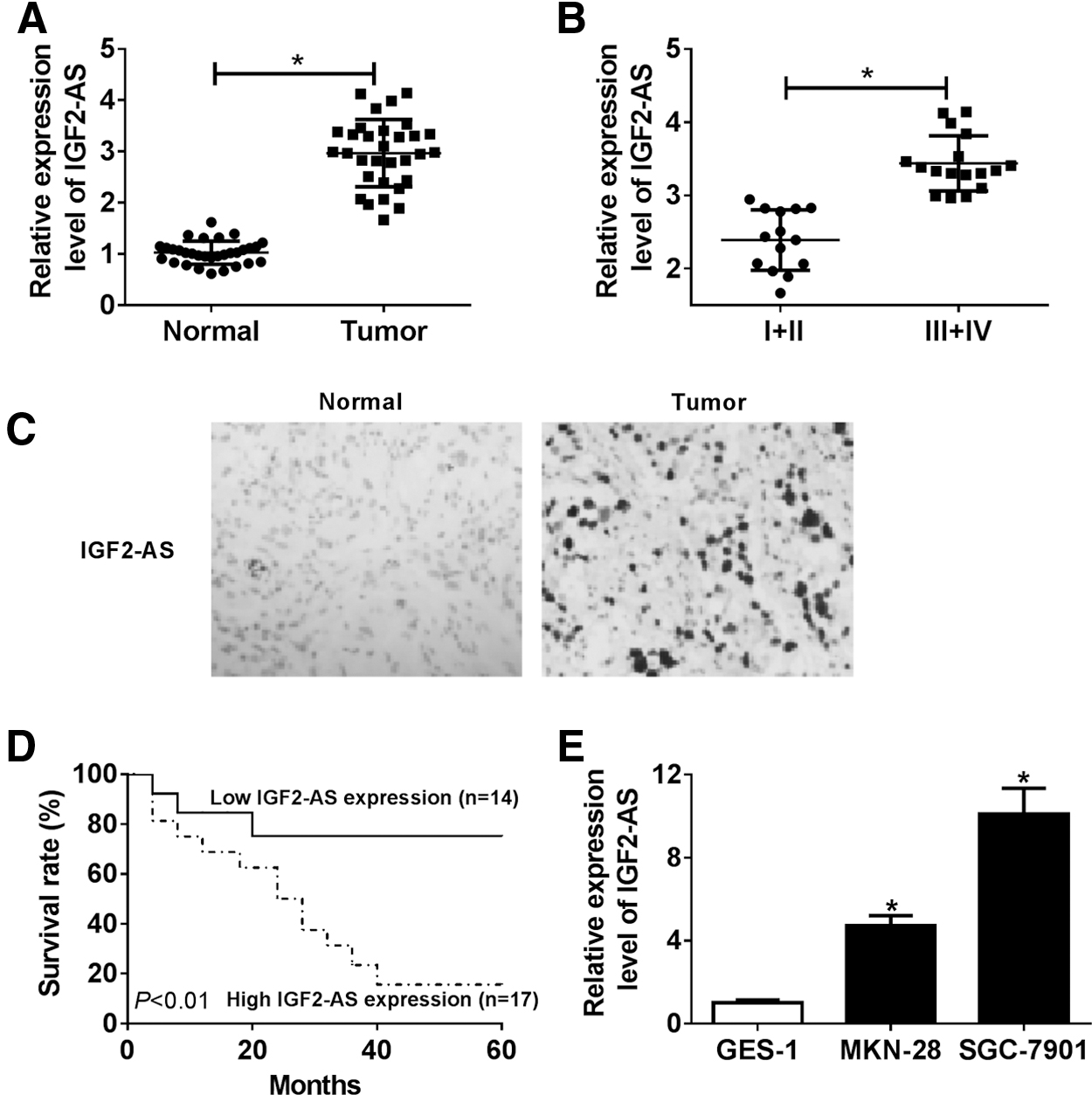
The IGF2-AS expression in GC tissues and cells and the role of IGF2-AS in GC prognosis. The expression of IGF2-AS in GC tissues and normal tissues
Correlation of Relative IGF2-AS Expression with the Clinicopathological Features of 31 Patients with Gastric Cancer
IGF2-AS, insulin growth factor 2 antisense.
Further, the correlation between the IGF2-AS expression and overall survival was analyzed via Kaplan–Meier survival analysis (log-rank test) in 31 GC patients and we found that highly expressed IGF2-AS predicted worse prognosis in GC patients compared with those with low expression (Fig. 1D). Subsequently, the expression of IGF2-AS in human GC cell lines, including MKN-28 and SGC-7901, and a normal GES-1 was measured and the data indicated, compared with GES-1 cells, that the expression of IGF2-AS was also higher in MKN-28 and SGC-7901 cell lines (Fig. 1E).
IGF2-AS directly binds to miR-937 and suppresses miR-937 expression
Based on the prediction of starBase v2.0 database, IGF2-AS might be a sponge of miR-937 with a putative binding site (Fig. 2A). To ascertain this hypothesis, the luciferase reporter assay was performed and a significant decrease of luciferase activity in MKN-28 and SGC-7901 cells co-transfected with IGF2-AS-WT and miR-937 was detected compared with the NC control, whereas the luciferase activity was not significantly affected after co-transfection with IGF2-AS-MUT and miR-937 (Fig. 2B, C). Then, we also conducted the RIP assay and the results showed that overexpression of miR-937 engendered the evident enrichment of IGF2-AS in MKN-28 and SGC-7901 cells after RIP-Ago2, whereas there was no response to RIP-IgG (Fig. 2D).
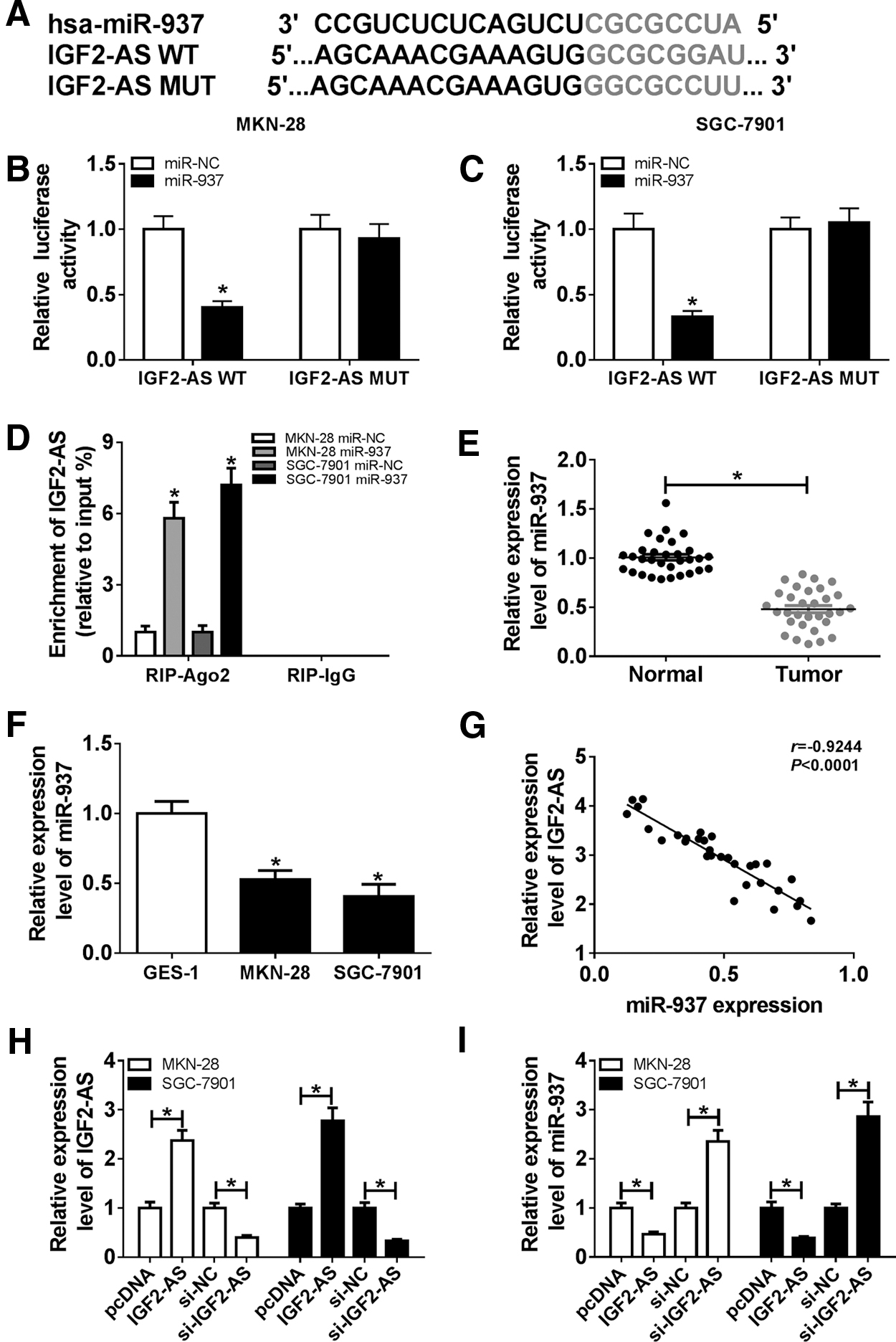
IGF2-AS was a sponge of miR-937 in GC cells.
Subsequently, the expression of miR-937 was investigated and we found a significant decrease of miR-937 expression in GC tissues and cell lines compared with the controls (Fig. 2E, F). In addition, a perfectly negative correlation between miR-937 and IGF2-AS was validated (r = −0.9244, p < 0.0001) (Fig. 2G). Therefore, to determine the relationship between miR-937 and IGF2-AS, MKN-28 and SGC-7901 cells were transfected with pcDNA or IGF2-AS or si-NC or si-IGF2-AS and the transfection efficiency was measured by qRT-PCR (Fig. 2H). In the meanwhile, we discovered that IGF2-AS transfection inhibited miR-937 expression, whereas silencing of IGF2-AS promoted miR-937 expression in MKN-28 and SGC-7901 cells (Fig. 2I). All these results revealed that IGF2-AS targeted as well as inhibited miR-937 expression.
IGF2-AS promotes cells proliferation, migration, and invasion abilities in GC via regulating miR-937
To explore the interaction of IGF2-AS and miR-937 in GC cells' progression, MKN-28 and SGC-7901 cells were transfected with si-NC, si-IGF2-AS, si-IGF2-AS+in-miR-NC, or si-IGF2-AS+in-miR-937. Then, the proliferation viability was detected by CCK-8 assay and the data showed that proliferation viability was notably suppressed though silencing IGF2-AS but enhanced by knockdown of miR-937 in MKN-28 and SGC-7901 cells (Fig. 3A, B). Simultaneously, decreased expression of IGF2-AS induced an obvious inhibition on cell migration and invasion abilities, whereas miR-937 inhibition could counteract these effects in MKN-28 and SGC-7901 cells (Fig. 3C, D). These data indicated that knockdown of IGF2-AS could inhibit the proliferation, migration, and invasion abilities in GC cells by targeting miR-937.
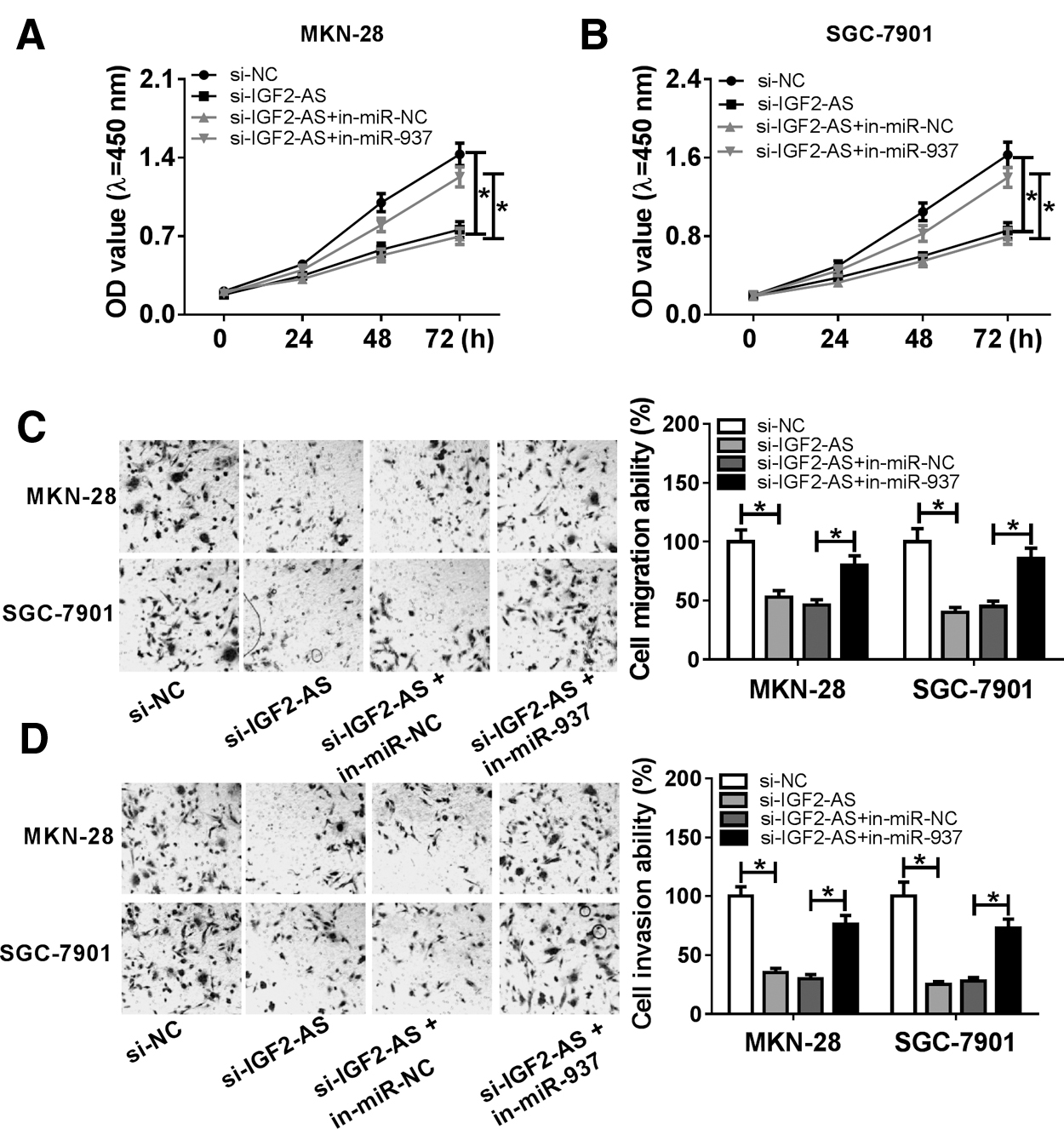
IGF2-AS promoted cell proliferation, migration, and invasion by regulating miR-937. Cells were transfected with si-RNA, si-IGF2-AS, si-IGF2-AS+miR-NC, or si-IGF2-AS+miR-937.
EZH2 is a target of miR-937
Since we found the functional role of miR-937 in regulating GC cancer, the associated signaling pathways or downstream target genes were speculated and EZH2 might be a target of miR-937 with a putative binding site according to the prediction of starBase v2.0 database (Fig. 4A). Immediately, luciferase reporter assay was used to confirm this hypothesis and the data indicated that the miR-937 mimic reduced the luciferase activities of the EZH2-WT reporter vector but not the EZH2-MUT reporter vector in MKN-28 and SGC-7901 cells (Fig. 4B, C).

EZH2 was a target of miR-937 in GC cells.
Subsequently, the level of EZH2 was measured by using qRT-PCR and high expression of EZH2 was detected in GC tissues and cell lines (MKN-28 and SGC-7901) compared with the normal tissues and GES-1 cells (Fig. 4D, E). In addition, a highly negative correlation between EZH2 and miR-937 expression was demonstrated (r = −0.9426, p < 0.0001) (Fig. 4F). Besides, western blot results showed that EZH2 was downregulated in GC cell lines when miR-937 was upregulated (Fig. 4G). All the data suggested that miR-937 targeted as well as inhibited EZH2 expression.
miR-937 inhibits cells proliferation, migration, and invasion abilities in GC via targeting EZH2
MKN-28 and SGC-7901 cells were transfected with miR-NC, miR-937, miR-937+pcDNA, or miR-937+EZH2 to investigate the effects of IGF2-AS and miR-937 on GC cells' progression. Then, we found that the abilities of cell proliferation, migration, and invasion were significantly inhibited by re-expressed miR-937 but enhanced by overexpressed EZH2 in MKN-28 and SGC-7901 cells (Fig. 5A–D). Therefore, we confirmed that miR-937 could inhibit cell proliferation, migration, and invasion abilities in GC via targeting EZH2 suppression.
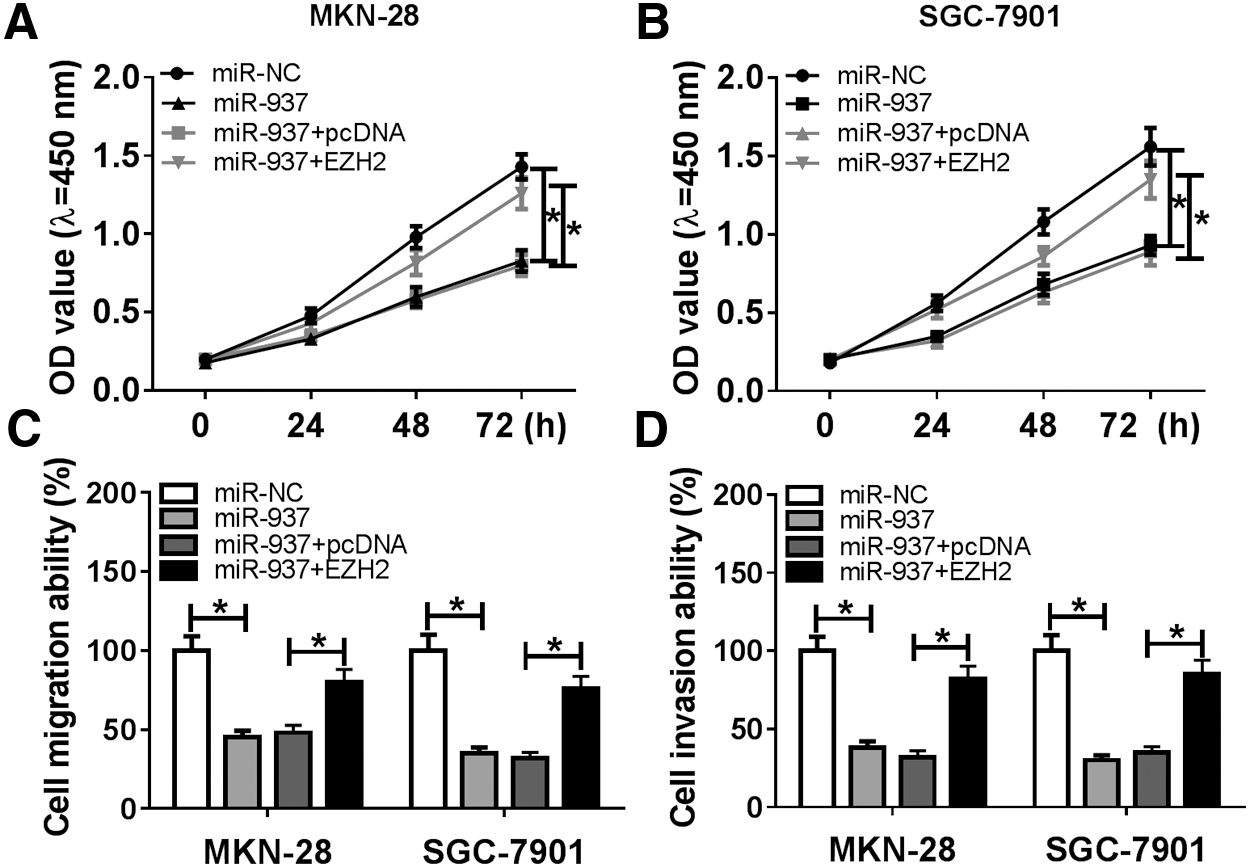
miR-937 inhibited cell proliferation, migration, and invasion by regulating EZH2. Cells were transfected with miR-NC, miR-937, miR-937+pcDNA, or miR-937+EZH2.
The IGF2-AS regulates EZH2 expression via directly interacting with miR-937
Based on miR-937/EZH2, we further determined whether IGF2-AS silencing could regulate EZH2 expression in GC cells by sponging miR-937. Results showed that IGF2-AS deletion inhibited EZH2 expression, whereas this inhibition could be rescued by following miR-937 inhibition in MKN-28 and SGC-7901 cells (Fig. 6A, B). Besides that, we also observed that IGF2-AS overexpression upregulated EZH2 expression, and miR-937 inhibition enhanced IGF2-AS-induced elevation on EZH2 level in MKN-28 and SGC-7901 cells (Supplementary Fig. S1A, B). Also, a highly positive correlation was detected between IGF2-AS and EZH2 expression (r = 0.9101, p < 0.001) (Fig. 6C). These data illustrated that IGF2-AS indirectly regulated EZH2 expression via sponging miR-937.
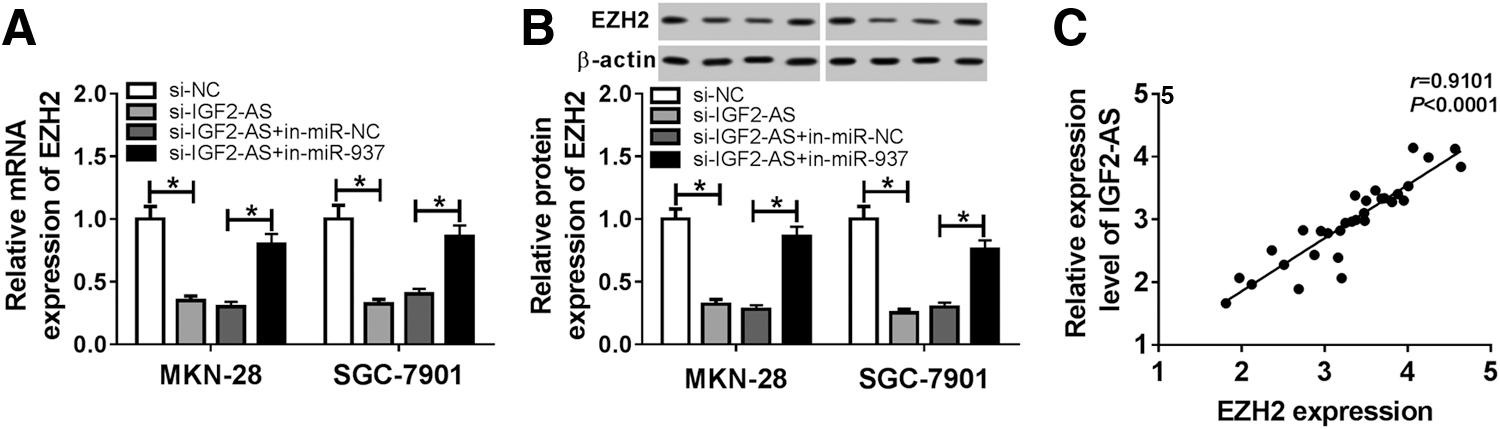
IGF2-AS promotes EZH2 expression via regulating miR-937. The mRNA
Knockdown of IGF2-AS inhibited the growth of GC by regulating miR-937 and EZH2 expression in vivo
The xenograft model was established to confirm the carcinogenic activity of IGF2-AS in vivo. Tumor volume and weight were measured and great inhibition was discovered in the sh-IGF2-AS group (Fig. 7A, B). Moreover, the expression of IGF2-AS, miR-937, and EZH2 was detected via qRT-PCR or western blot, respectively. We found significant inhibition of IGF2-AS (Fig. 7C) and EZH2 (Fig. 7E, F) expression but a promotion of miR-937 expression (Fig. 7D). Taken together, knockdown of IGF2-AS inhibited GC tumor growth in vivo.

Knockdown of IGF2-AS inhibited tumor growth by regulating miR-937 and EZH2 expression in vivo. Xenograft model was established in nude mice injected with transfected Lenti-sh-NC or Lenti-shRNA-IGF2-AS.
Discussion
IGF2-AS is an RNA gene, and it is affiliated with the noncoding RNA class. IGF2-AS is well known to be associated with Diabetes Mellitus, Insulin-Dependence, and Wilms Tumor 1. 13,18 However, the regulatory roles of IGF2-AS as it served as a cancer-related lncRNA in diverse tumors have been investigated. For example, Zhang et al. found that overexpressed IGF2-AS functioned as an antitumor factor that inhibited cell proliferation, migration, and drug resistance in non-small cell lung cancer (NSCLC) and was a prognostic biomarker for NSCLC. 16 Cao et al. revealed that IGF2-AS was upregulated in prostate cancer and acted as an oncogene to promote cancer cell proliferation, thus affecting tumor survival. 19 Liang et al. discovered that IGF2-AS was elevated in colorectal cancer and associated with clinical prognosis. 20
Besides that, a recent study also indicated that IGF2-AS functioned as an oncogene to induce promotion, proliferation, and motility in gastric adenocarcinoma by regulating the miR-503/SHOX2 axis. 21 Thus, IGF2-AS may be a potential regulator for cancer progression, including GC. In this study, we focused on the roles of IGF2-AS in GC. First, we found that IGF2-AS was greatly elevated in GC tissues, especially in the high-grade GC tissues (grade III–IV), and we found that highly expressed IGF2-AS predicted worse prognosis in GC patients compared with those with low expression. After that, functional experiments indicated that IGF2-AS silence restrained cell proliferation, migration, and invasion in GC in vitro as well as suppressed tumor growth in vivo.
In spite of the biological effects of IGF2-AS in gastric adenocarcinoma having been identified, the detailed molecular mechanisms by which IGF2-AS affects tumorigenesis in GC still need to be elaborated. With the proposed competing endogenous RNA hypothesis, lncRNAs were indicated to serve as sponges of miRNAs to induce the inhibition of regulatory effects of miRNAs on targeted messenger RNAs (mRNAs) 22,23 and the miRNA-lncRNA-mRNA axis has been reported to play important roles in the occurrence and progression of GC. 24 Here, we further explored the miRNA targets of IGF2-AS, and miR-937 was identified to be a target of IGF2-AS. miR-937 is a novel recognized cancer-related miRNA.
Recent evidence suggests that miR-937 is downregulated and performs antitumor functions in breast cancer 25,26 and GC, 27 which may be a potential therapeutic target for cancers. In this study, we found that the levels of miR-937 in GC tissues and cell lines were decreased, which was consistent with previous studies. 27 Subsequently, co-expression analysis showed that IGF2-AS negatively regulated miR-937 expression both in vitro and in vivo. In addition, functional experiments indicated that restoration of miR-937 expression inhibited cell carcinogenesis in GC, and IGF2-AS exerted antitumor effects by interacting with miR-937.
EZH2 is a histone-lysine N-methyltransferase enzyme that has been revealed to be involved in the modulation of cell tumorigenesis, proliferation, invasion, and migration. 28 EZH2 was found to act as an oncogene and to be involved in poor prognosis or increased cancer risk in GC. Upregulation of EZH2 is associated with p53 and Ki-67 expressions and it is notably correlated with GC cells' invasion and metastasis. 29 –31 Therefore, in this work, we detected the correlation between EZH2 and miR-937 in GC and results indicated that EZH2 was a target of miR-937. Moreover, rescue assay was performed and overexpressed EZH2 could reverse re-expression of miR-937-mediated inhibition on cell carcinogenesis in GC. In addition, we also observed a highly positive correlation between IGF2-AS and EZH2. Meanwhile, IGF2-AS could indirectly regulate EZH2 expression via sponging miR-937 both in vivo and in vitro.
Conclusions
Our results demonstrated that IGF2-AS was elevated in GC tissues and cell lines. Overexpressed IGF2-AS was closely associated with worse prognosis as well as cancer progression in GC patients. In addition, we first proposed the IGF2-AS/miR-937/EZH2 axis in GC progression, which provides novel prognostic and therapeutic targets for GC.
Footnotes
Authors' Contributions
Conception and design: Z.L., Z.L., W.Z., and J.X.; development of methodology: Z.Z. and J.Z.; acquisition of data: Z.Z. and J.Z.; analysis and interpretation of data: Z.L. and Z.L.; writing, review, and revision of article: Z.L., Z.L., W.Z., and J.X. All coauthors have reviewed and approved of the article before submission.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.