
Abstract
Cancer Biotherapy and Radiopharmaceuticals officially retracts the paper entitled, “Targeted Regulation of FoxO3a by miR-372 to Mediate Gastric Carcinoma Cell Apoptosis and DDP Drug Resistance” by Changting Wang, Chunxiao Wang, Chengzhi Qiu, and Zhiming Su (Cancer Biother Radiopharm 2020;35(10):753–759; doi: 10.1089/cbr.2019.3299).
The published paper was flagged as a potential submission from a paper mill. In an effort to determine the paper's validity, the journal contacted the corresponding author, Dr. Changting Wang, and requested the raw data files as they pertained to the work. Dr. Wang was not able to provide the data and indicated via email, “Our experiment is still going on. At the end of the experiment, we will add the corresponding content as soon as possible.”
Dr. Wang's response is impermissible. As per customary practices of study protocols, raw data image files should be archived and readily available upon request. Conducting new experiments after the work has been published is not acceptable as it indicates that the results of the work may not be reproducible and renders the conclusions questionable.
The journal officially retracts the paper from the literature based on Dr. Wang's indication of ongoing experiments and inability to produce the raw data. This is a violation of standard protocols and is considered an infraction against the rigorous standards of scientific publishing.
The Editor and Publisher of Cancer Biotherapy and Radiopharmaceuticals are committed to preserving the scientific literature and the community it serves.
Introduction
Gastric cancer (GC) is a malignant tumor in the digestive tract with high incidence and mortality rate among cancers worldwide. 1,2 Chemotherapy is frequently used in GC treatment, but its efficiency is compromised by drug resistance, which is one important factor affecting patient survival and prognosis. 3,4
Forkhead transcription factor O (FoxO) subfamily is a group of transcriptional factors that are highly conserved during evolution and widely involved in the regulation of multiple biological processes, including embryonic development, cell proliferation, cell cycle, and apoptosis. 5 –7 FoxO3a is a tumor suppressor gene in FoxO transcriptional factor family. Accumulating evidences show the correlation between expressional or functional deficits in occurrence, progression, and drug resistance of multiple tumors, including breast carcinoma, 8 ovarian cancer, 6 and prostate cancer. 9 FoxO3a downregulation has also been suggested to be closely related with GC occurrence and progression. 10,11
MicroRNA (miR) are a group of small-molecule single-stranded endogenous noncoding RNA of 22–25 nucleic acids in length. They can regulate target gene expression by complementary binding 3′-untranslated region (3′-UTR) of target gene mRNA to degrade mRNA or to suppress translation. More than 30% of human genes can thus be modulated by miR, which occupies only 1% of total genome. 12 The role of miR expression or functional abnormality in tumorigenesis draws increasing research interests. 13,14 Previous studies showed abnormally elevated miR-372 expression in GC tumor tissues, 15 –17 indicating the possible oncogenic role of miR-372 in GC pathogenesis. Bioinformatics analysis revealed the existence of complementary binding sites between miR-372 and 3′-UTR of Foxo3a mRNA. This study investigated if miR-372 plays a role in mediating Foxo3a expression and affecting proliferation, apoptosis, and cisplatin (DDP) resistance of GC cells.
Materials and Methods
Major reagents and materials
Normal human gastric mucosal cell line gastric epithelial cell line-1 (GES-1), GC cell lines MGC803 and MKN28 were purchased from Jiniou Biotech (China). HEK293T cell was purchased from Cell Bank of Chinese Academy of Science. RPMI 1640 culture medium, serum-free Opti-MEM medium, and fetal bovine serum (FBS) were purchased from Gibco. TRIzol and Lipofectamine 2000 transfection reagent were purchased from Thermo. The QuantiTect SYBR Green RT-PCR Kit was purchased from Qiagen (Germany). miR-372 mimic, miR-372 inhibitor, and miR-negative control (NC) were purchased from RioBio (China). Rabbit anti-human Foxo3a and anti-β-actin polyclonal antibody were purchased from CST. Goat anti-rabbit horseradish peroxidase (HRP)-conjugated secondary antibody was purchased from Jackson ImmunoResearch. BeyoECL Plus developing reagent, and EdU Cell Proliferation Test Kit were purchased from Beyotime (China). The Annexin V/PI Cell Apoptotic Assay Kit was purchased from BioLegend. Dual luciferase reporter system Dual-Glo Luciferase Assay System and pMIR plasmid were purchased from Promega. DDP was purchased from Sigma. The Cell Counting Kit-8 (CCK-8) was purchased from Dojindo Molecular Technologies (Japan).
Cell culture
GES-1, MGC803, and MKN28 cells were incubated in RPMI 1640 medium containing 10% FBS and were kept in 37°C incubator with 5% CO2 for passage at 1:4 ratio. Cells at log-growth phase were used for further assays.
Establishment of DDP drug-resistant cell model
Generation of DDP drug-resistant cell model: MGC803 and MKN28 cells at log-growth phase were treated with 0.1 μg/mL DDP, which was added into culture medium. After stable growth for a 2-week period, the DDP concentration was elevated to 0.2 μg/mL for 2 more weeks of incubation. With similar approaches, DDP concentration was gradually increased to 0.4, 0.8, and 1.6 μg/mL until reaching stable growth and repeated passage of MGC803 and MKN28 cells at 1.6 μg/mL DDP. Drug-resistant GC cell lines MGC803/DDP and MKN28/DDP were thus generated.
MGC803, MKN28, MGC803/DDP, and MKN28/DDP cells were inoculated into a 96-well plate at 10,000 cells per well density. After 24 h of attached growth, cells were treated with 0, 0.1, 1, 10, 100, and 1000 μg/mL DDP, with six parallel wells for each concentration. After 48 h incubation, 10 μL CCK-8 solution was added into each well. Absorbance values at 450 nm wavelength (A450) were measured after 4 h reaction. Inhibition rate = (1 − A450 of drug treatment group)/A450 of control group × 100%. SPSS software was used to calculate the drug concentration to inhibit 50% cell growth (IC50). Resistance index (RI) = IC50 of drug-resistant cells/IC50 of parental cells.
Flow cytometry measuring cell proliferation
The EdU Flow Cytometry Kit was used to measure cell proliferation. In brief, cells were resuspended into RPMI 1640 medium containing 10% FBS. After incubation in 10 μM 5-Ethynyl-2′-deoxyuridine (EdU) for 2 h at 37°C, cells were inoculated into a culture plate for 48 h continuous incubation. Cells were digested by trypsin and were collected, followed by phosphate-buffered saline (PBS) rinsing once in centrifugation. Cells were fixed in paraformaldehyde, washed in PBS centrifugation, and were permeabilized by adding 100 μL buffer. Five hundred microliters of reaction buffer was added for 30-min dark incubation at room temperature. Three milliliters of permeabilization buffer was added for centrifugation, and cells were resuspended in 500 μL wash buffer. Cell proliferation was measured on FC500MCL flow cytometry (Beckman, Coulter).
Dual luciferase activity assay
Full-length fragment or mutant form of 3′-UTR of Foxo3a gene was amplified by polymerase chain reaction (PCR), digested by dual restriction enzymes, and was ligated into pMIR plasmid. After transforming bacteria, plasmids with correct sequence insertion was screened and were named as pMIR-Foxo3a-WT and pMIR-Foxo3a-MUT. Lip2000 was used to cotransfect pMIR-Foxo3a-WT (or pMIR-Foxo3a-MUT) and miR-72 mimic (or miR-NC, miR-375 mimic) into HEK293T cells, which were kept in a 5% CO2 incubator at 37°C. With 48 h continuous incubation, dual luciferase activity was measured by the Dual-Glo Luciferase Assay System Kit.
Cell transfection and grouping
Cultured MGC803/DDP and MKN28/DDP cells were divided into two groups: miR-NC transfection group and miR-375 inhibitor transfection group. In brief, 10 μL Lip2000, 50 nmoL miR-NC, or 50 nmoL miR-375 inhibitor were diluted in 100 μL serum-free Opti-MEM. After 5 min of incubation at room temperature, Lip2000 was gently mixed with miR-NC and miR-375 inhibitor for 20 min of incubation at room temperature. Transfection mixture was added into Opti-MEM for 6 h of incubation. Opti-MEM was then removed and switched to RPMI 1640 complete medium with 10% FBS for 72 h of incubation.
Cells from all four transfection groups were inoculated into a 6-well plate. When cells reached 50% confluence, 1.6 μg/mL DDP was added for treatment. After 48 h continuous culture, cell apoptosis was measured by flow cytometry as described below.
All four groups of cells were digested by trypsin and were collected. After incubation in 10 μM EdU for 2 h, cells were continuously incubated for 48 h as described above, and were treated with 1.6 μg/mL DDP. Cell proliferation was measured by the EdU Assay Kit.
quantitative real-time polymerase chain reaction for gene expression assay
TRIzol was used to extract cellular RNA. One-step quantitative real-time polymerase chain reaction (qRT-PCR) was used to measure target gene expression from RNA using the QuantiTech SYBR Green RT-PCR Kit. In a 20 μL qRT-PCR reaction system, one added 10.0 μL 2XQuantiTect SYBR Green RT-PCR Master Mix, 0.5 μm/L forward and reverse primer (0.5 μm/L), 2 μg template RNA, 0.5 μL QuantiTech RT Mix, and ddH2O upto 20.0 μL. qRT-PCR conditions were: 45°C for 5 min, and 94°C for 30 s, followed by 40 cycles each consisting of 95°C 5 s and 60°C 30 s. Gene expression was measured on Bio-Rad CFX96 real-time fluorescent quantitative PCR.
Western blot
One hundred microliters of radioimmunoprecipitation assay lysis buffer was added into each 106 cells. Protein concentration was measured by bicinchoninic acid (BCA) method. Forty microgram protein samples were loaded and were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (45 V, 150 min). Proteins were then transferred to polyvinylidene difluoride (PVDF) membrane (300 mA, 100 min), which was blocked using 5% defat milk powder. Primary antibody (Foxo3a at 1:800, β-actin at 1:5000) was added for 4°C overnight incubation. On the next day, the membrane was washed in Phosphate Buffered Saline with Tween™ 20 (PBST) for three times. HRP-conjugated secondary antibody (1:10,000 dilution) was added for 60 min of incubation at room temperature. After rinsing the membrane three times, BeyoECL Plus development reagent was added for 2–3 min of development in the dark, followed by exposure and development. The X-ray film was scanned for data storage.
Cell apoptosis assay
Cells were digested by trypsin and were collected, followed by PBS rinsing. One hundred microliters of Annexin V Binding Buffer was added to resuspend cells. Ten microliters Annexin V-FITC and 5 μL PI dye were added for 15 min at room temperature. Four hundred microliters of Annexin V Binding Buffer was added and cell apoptosis was measured by FC500 MCL flow cytometry (Beckman, Coulter).
Statistical analysis
SPSS 18.0 software was used for data statistics and comparison. Measurement data were presented as mean ± standard deviation. Comparison of measurement data between two groups was performed by t-test. The comparison of measurement data among multiple groups was performed by one-way analysis of variance (ANOVA) first, followed by Bonferroni post-hoc test between two groups. A statistical significance was defined when p < 0.05.
Results
Targeted regulation between miR-372 and Foxo3a mRNA
Bioinformatics analysis showed the existence of complementary binding sites between miR-372 and 3′-UTR of Foxo3a mRNA (Fig. 1A). Dual luciferase gene reporter assay showed that transfection of miR-372 mimic significantly depressed relative luciferase activity in HEK293T cells transfected with pMIR-Foxo3a-WT, while the relative luciferase activity in HEK293T cells transfected with pMIR-Foxo3a-MUT was not changed (Fig. 1B). These results showed that miR-372 could target 3′-UTR of Foxo3a mRNA to suppress its gene expression.
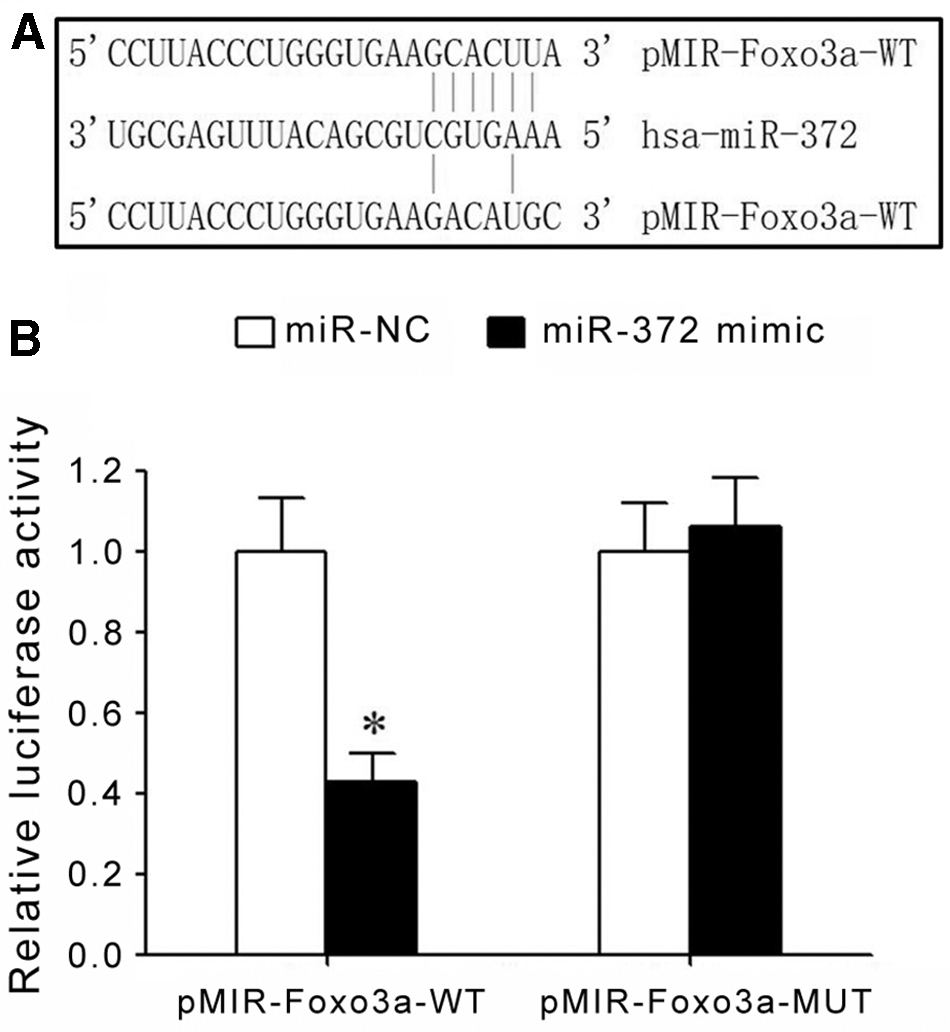
Targeted regulation between miR-372 and Foxo3a mRNA.
Strong proliferation activity and drug resistance in resistant GC cells
CCK-8 assay showed that, under the treatment of same DDP dosage, parental GC cells MGC803 (Fig. 2A) and MKN28 (Fig. 2B) presented significantly lower proliferation activity than MGC803/DDP and MKN28/DDP cells. MGC803 cells had IC50 of 2.15 ± 0.26 μg/mL, while drug-resistant cells MGC803/DDP had IC50 of 31.23 ± 2.13 μg/mL, thus having a RI of 14.53 relative to parental MGC803 cells. MKN28 cells had an IC50 value of 3.53 ± 0.29 μg/mL, and MKC28/DDP cells showed IC50 values at 52.96 ± 3.77 μg/mL, thus having an RI value at 15.29 relative to parental MKN28 cells.
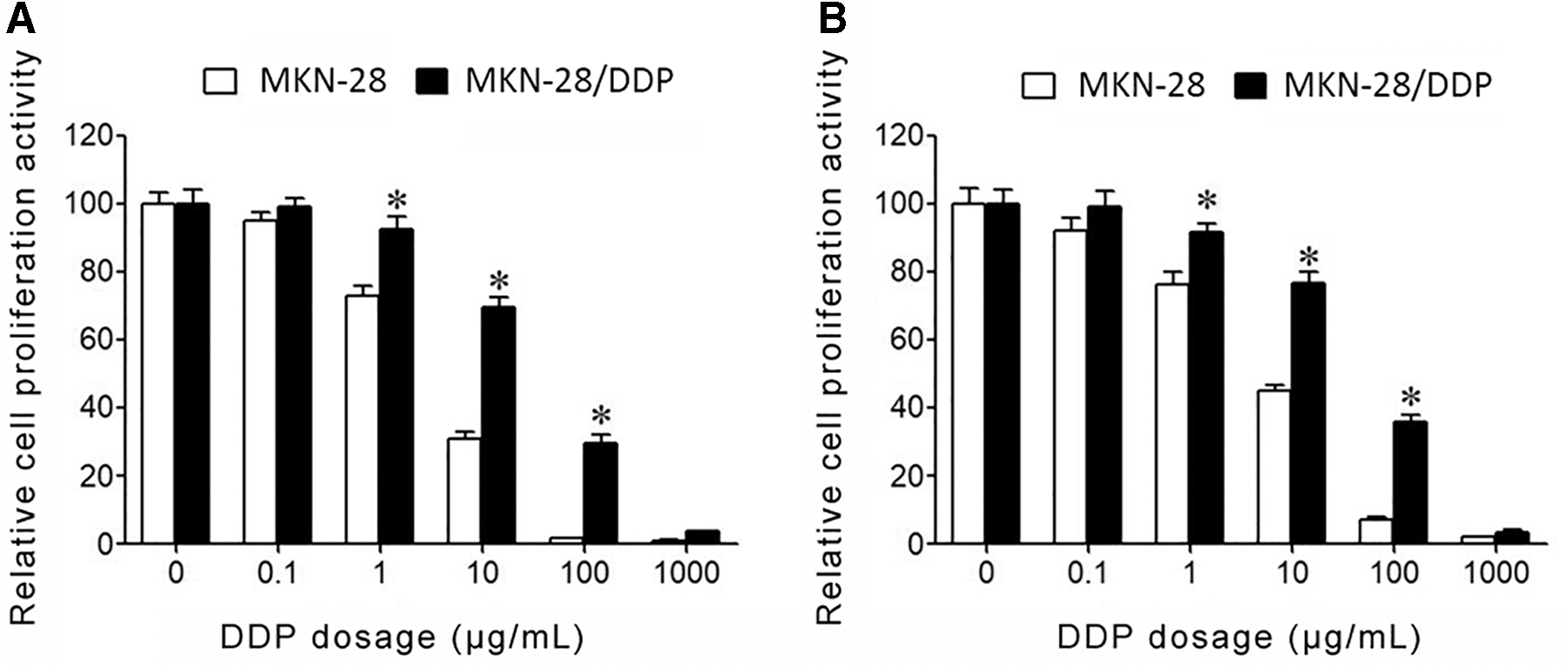
Strong proliferation activity and drug resistance of GC cells.
Upregulation of miR-372 and downregulation of Foxo3a
qRT-PCR results showed that, compared with normal human gastric mucosal cell GES-1, GC cells presented significantly elevated miR-372 expression, which was further enhanced in drug-resistant cells compared with parental GC cells (Fig. 3A). qRT-PCR results also suggested that compared with GES-1 cells, GC cells showed significantly decreased Foxo3a mRNA expression, which was further suppressed in drug-resistant cells (Fig. 3B). Western blot results showed that compared with GES-1 cells, GC cells had lower Foxo3a protein expression, which was decreased in drug-resistant cells (Fig. 3C).

Upregulation of miR-372 and downregulation of Foxo3a.
Downregulation of miR-375 suppressed DDP resistance of GC cells and potentiated apoptosis
qRT-PCR results showed that compared with miR-NC group, miR-372 inhibitor transfection significantly upregulated Foxo3a mRNA expression in MGC803/DDP and MKN28/DDP cells (Fig. 4A). Western blot results showed that transfection of miR-372 inhibitor significantly elevated Foxo3a protein expression in MGC803/DDP and MKN28/DDP cells (Fig. 4B). Flow cytometry revealed that transfection of miR-372 inhibitor significantly elevated apoptosis of MGC803/DDP and MKN28/DDP cells (Fig. 4C), whereas suppressed cell proliferation potency (Fig. 4D). However, transfection of miR-372 mimics did not affect the expression of Foxo3a, cell apoptosis, and proliferation.

miR-372 downregulation suppressed DDP resistance of GC cells and enhanced apoptosis.
Discussion
It is estimated that more than 1.3 million people are newly diagnosed with GC per year. 18 The incidence of GC largely varies across countries and regions due to differences in ethnic groups, life habits, and diet structures. East Asia is one prevalent area for GC and China has the highest incidence, as nearly one half of all GC cases worldwide occur in China. 19 In each year, about 679,000 people were newly diagnosed as GC in China, and around 498,000 people died from it. 20 GC has an insidious disease onset without significant clinical symptoms at an early stage. 21,22 Reaching advanced and terminal stage, however, GC has rapid progression with high malignancy, frequent invasion or distal metastasis, plus lower sensitivity to radiotherapy or chemotherapy, all of which largely compromise its treatment efficiency with worsening survival and prognosis. 23,24 Although major progression has been obtained using combined treatment, including surgery, chemo/radiotherapy, gene therapy and tumor immune therapy, and patient's survival has been elongated to certain extents, the prognosis of GC is still unfavorable and 5-year postoperative survival rate is around 15%–20%. 25
FoxO transcriptional factor family consisted of four members: FoxO1, FoxO3a, FoxO4, and FoxO6, among which FoxO3a is the most widely studied one. 6,9 Belonging to one highly conserved transcriptional factor family, Foxo3a gene locates on human chromosome 6q21 and encodes one protein molecule consisting of 673 amino acid residues. FoxO3a has featured and highly conserved forkhead DNA-binding domain that consists of three α-helices, three β-sheets, and 2 wing-like structures. 26,27 FoxO3a is a widely studied tumor suppressor gene and can regulate the expression of various genes related with cell proliferation, cell cycle, and cell apoptosis, including Bcl-2-interacting mediator of cell death (BIM), p53 upregulated modulator of apoptosis (PUMA), cyclin-dependent kinase inhibitor (CKI), p27Kip1, and cyclin-dependent kinase D1 (cyclin D1), thus regulating various signal pathways and biological processes as the pivotal point. 11,28,29 Accumulating evidence shows the close correlation between FoxO3a downregulation and occurrence and progression of GC. 10,11 miR-372 is involved in the onset, progression, and metastasis of various tumors, including colorectal carcinoma, 30 breast cancer, 31 and testicular germ line carcinoma. 32 Previous studies have revealed significantly upregulated miR-372 in GC tumor tissues, 15 –17 suggesting that miR-372 might be the oncogenic factor in GC pathogenesis. Bioinformatics analysis further showed the existence of complementary binding sites between miR-372 and 3′-UTR of Foxo3a mRNA. This study thus investigated if miR-372 played a role in modulating Foxo3a expression and in affecting proliferation, apoptosis, and DDP resistance of GC cells.
In this study, dual luciferase gene reporter assay showed that transfection of miR-372 mimic remarkably suppressed relative luciferase activity in HEK293T cells transfected with pMIR-Foxo3a-WT, confirming targeted regulation between miR-372 and Foxo3a. CCK-8 assay showed that DDP treatment significantly suppressed proliferation activity of parental GC cells with significantly lower proliferation activity in drug-resistant cells. Further calculation for IC50 of parental and resistant cells showed significantly higher IC50 in drug-resistant GC cells compared with parental tumor cells, indicating successful generation of drug-resistant GC cell mode. Both gene and protein assays showed significantly higher miR-372 expression in GC cells compared with normal gastric mucosal cells, while Foxo3a expression was decreased. Drug-resistant GC cells had significantly higher miR-372 expression than parental tumor cells, whereas Foxo3a expression was significantly lower. Results indicated the role of miR-372 in suppressing Foxo3a expression, which is correlated with GC occurrence in addition to drug resistance.
Currently, few studies have been conducted to assess the correlation between miR-372 and drug resistance of GC cells. This study further investigated if miR-372 could affect DDP resistance of GC cells through modulating Foxo3a expression. Results showed that transfection of miR-375 inhibitor could remarkably enhance Foxo3a expression in drug-resistant GC cells, and increased apoptosis of GC cells that can maintain stable growth in DDP, and weakened cell proliferation potency plus elevated DDP drug sensitivity. In a study for biological effect of miR-372 and GC cells, Cho et al. showed that downregulation of miR-372 in GC cell line human gastric adenocarcinoma cell line (AGS) could suppress GC cell proliferation, induce cell cycle arrest, and facilitate cell apoptosis through targeted inhibition on gene LATS2. 33 Staedel et al. found the oncogenic role of miR-372 in GC biological regulation, as miRNA inhibitor PA-1 could weaken GC cell AGS proliferation activity or clonal formation potency through suppressing miR-372 expression. 15 Vo et al. further showed that the miR-372 upregulation played an oncogenic role in GC, as inhibition of miR-372 expression exerted tumor suppressor role in resisting GC cell proliferation. 16 Zhou et al. showed that transfection of miR-372 inhibitor into GC cells significantly suppressed cell proliferation and induced cell apoptosis, while miR-372 mimic transfection significantly facilitated GC cell proliferation. 17 Moreover, such oncogenic effect of miR-372 is accomplished through targeted inhibition on TNFAIP1 gene. Staedel et al. found the correlation between miR-372 upregulation and malignant biological properties of GC cells, and 8-mer LNAs can inhibit GC cell growth and proliferation, and weakens in vivo growth and tumorigenicity of GC cells through targeted inhibition on miR-372. 15 This study combined miR-372 and targeted regulation of Foxo3a, and revealed the role of miR-372 upregulation in suppressing Foxo3a expression, facilitating GC cell proliferation, decreasing apoptosis, and enhancing DDP resistance, all of which have not been previously reported and are thus a novelty of this study. However, whether miR-372 could regulate Foxo3a expression in the drug resistance of GC patient is still unclear and requires further investigation.
Conclusions
miR-372 upregulation is associated with DDP resistance of GC cells. Downregulation of miR-372 could inhibit proliferation potency of drug-resistant GC cells through upregulating Foxo3a, to facilitate cell apoptosis and suppress DDP resistance.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
This work was supported by the Science and Technology Project of Quanzhou (no. 2018N013S).