
Abstract
Background:
Cervical cancer (CC) is one of the most common cancers among women in the world. Long noncoding RNAs and microRNAs were identified as important regulators in many physiological processes. The objective of this study was to illuminate the mechanism of X-inactive-specific transcript (XIST)/miR-889-3p/Sine oculis homeobox 1 (SIX1) axis in CC.
Methods:
The expression levels of XIST, miR-889-3p, and SIX1 were detected by quantitative real-time polymerase chain reaction. Cell proliferation was assessed by cell counting Kit 8 assay. Cell migration and invasion were evaluated by transwell assay. Cell apoptosis was detected by flow cytometry assay. Murine model was established using transfected Me180 cell. The interaction among XIST, miR-889-3p, and SIX1 was tested by dual-luciferase reporter and RNA immunoprecipitation assays. Protein level of SIX1 was measured by Western blot.
Results:
XIST was highly expressed in CC tissues and cells. Silenced XIST inhibited proliferation, migration, and invasion and induced apoptosis. Moreover, XIST silencing blocked tumor growth in vivo. XIST directly bound to miR-889-3p, and XIST promoted proliferation, migration, and invasion and hindered apoptosis by suppressing miR-889-3p expression. MiR-889-3p targeted SIX1 and negatively regulated SIX1 expression. Furthermore, miR-889-3p had a low expression and SIX1 had a high expression in CC tissues and cells. XIST knockdown reduced SIX1 level by targeting miR-889-3p. In addition, miR-889-3p inhibition abolished the effects of SIX silencing on proliferation, migration, invasion, and apoptosis.
Conclusion:
XIST knockdown restrained cell proliferation, migration, and invasion and promoted apoptosis by regulating miR-889-3p/SIX1 axis.
Introduction
Cervical cancer (CC), which is a common disease of the cervix, has developed into the second leading malignancy among women all over the world. 1 Over 500,000 people are diagnosed each year, and ∼200,000 patients die of CC each year all over the world. 2 Although much improvement has been made in the treatment of CC, the survival rate is still unsatisfactory. 3 Metastasis and easy recurrence were the main reasons leading to the high mortality of CC patients. 4 Of importance, the age of patients first diagnosed with CC tended to be younger in recent years. 5 Current therapeutic methods, including chemotherapy and radiotherapy, could improve some CC patients' condition to some degree, whereas for some late-stage CC patients, these approaches cannot completely cure them. 6 Therefore, an early diagnostic marker is urgently needed for the prevention and treatment of CC.
Long noncoding RNAs (lncRNAs), which were considered as the “garbage RNAs,” have no coding protein potential. 7,8 In general, they have >200 nucleotides in length and function as competing endogenous RNAs (ceRNAs), which bind to microRNAs (miRNAs). 9 Many identified lncRNAs play vital roles in various physiological processes. In addition, lncRNAs were involved in the regulation of cancer development. X-inactive-specific transcript (XIST) is an important human lncRNA and is from the region of chromosome X inactivation center (XIC). 10 XIST was proved to be a predictor of human cancer prognosis because of its aberrant expression in multiple cancers including CC. 11 Tang et al. reported that XIST was highly expressed in lung cancer, and its knockdown exerted inhibitory effects on cell progression and metastasis in vitro and in vivo. 12 Chen et al. discovered the repressive effects of XIST downregulation on cell growth and cell cycle in CC. 13 However, research about the molecular basis of XIST in CC needs to be performed.
MiRNAs, as a part of noncoding RNAs, play a pivotal role in the regulation of target mRNA expression resulting in the loss or inhibition of mRNA function. 14 They often contain 18–25 nucleotides in length. 15 It has been widely reported that miRNAs are important regulators during the pathogenesis of many human diseases. MiR-889 has been demonstrated to promote proliferation in esophageal squamous cell carcinomas (ESCC) and closely related to the clinical pathological characteristics of ESCC patients. 16 In addition, Han et al. determined that miR-889 promoted cell growth and tumorigenesis of nonsmall cell lung cancer by targeting KLF9. 17 Thus, it is necessary to explore the functional role and the potential regulatory mechanism of miR-889-3p in CC.
Sine oculis homeobox 1 (SIX1) is a member of the SIX family that plays a crucial role in the growth of various organs. 18 Emerging studies implicated that SIX1 was highly expressed in many cancers, such as colorectal, ovarian, and hepatocellular carcinoma. 19,20 Of note, the high expression of SIX1 was closely associated with increased mortality of multiple cancers. 21 Accumulating evidence has indicated that SIX1 played a crucial role in tumor metastasis and was involved in the diagnosis and prognosis of various tumors. 22 Although the role of SIX1 in CC has been reported previously, the interaction among XIST, miR-889-3p, and SIX1 remained unknown.
In this research, we determined the expression of XIST and further investigated the underlying mechanism of XIST in CC, which might provide a potential early diagnostic biomarker for better diagnosis and treatment of CC.
Materials and Methods
Patients and cell lines
Thirty five CC patients enrolled in this study were from The first Affiliated Hospital of Hebei North University from January to August in 2018. All patients were divided into four groups according to TNM staging system. CC tissues and the adjacent normal tissues, which were located more than 5 cm far away from the tumor tissues were taken from all participants. All patients have signed written consents before the sample collection. All experiments involved in the present research were permitted by the Ethics Committee of The first Affiliated Hospital of Hebei North University.
Human ectocervical cell line Ect1/E6E7 obtained from American Tissue Culture Collection (Manassas, VA) and human cervical cell lines (Me180 and Hela) purchased from Korean Cell Line Bank (KCLB, Seoul, Korea) were all placed in Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA), which was supplemented with 10% fetal bovine serum (Hyclone, Logan, UT).
Cell transfection
RNA silencing against XIST and negative control (si-XIST#1, 5′-GUAUCCUAUUUGCACGCUATT-3′, si-XIST#2, 5′-GCUGACUACCUGAGAUUUATT-3′ si-XIST#3 5′-GCCCUUCUCUUCGAACUGUTT-3′ and si-NC, 5′-CGTTAATCGCGTATAATACGCGTAT-3′), short hairpin RNA for XIST and control (sh-XIST, 5′-AUAACAGUAAGUCUGAUAGAGGACA-3′ and sh-NC 5′-AUGCUAGAUGCAUACGAAGCA-3′), RNA silencing against SIX1 (si-SIX1, 5′-GCAACACAATTGGCTGCAA-3′) were purchased from Genepharma (Shanghai, China). The mimics of miR-889-3p and NC were bought from RIBOBIO (Guangzhou, China). The overexpression plasmid of XIST and miR-889-3p and empty plasmid (miR-NC and pcDNA) were provided by Hanbio (Shanghai, China). Cell transfection of all the above oligonucleotides and plasmids was performed using the Lipofectamine 2000, which was purchased from Invitrogen (Carlsbad, CA). Then, transfected cells were incubated at 37°C for 48 h.
Quantitative real-time polymerase chain reaction
CC tissues and the adjacent normal tissues were ground, and cells were collected for the extraction of total RNA. Then, the quality of RNA was determined by gel electrophoresis, and the quantity was checked by NanoDrop (Thermo Fisher Scientific, Rockford, IL). For the detection of lncRNA and mRNA expression, cDNA was amplified by the polymerase chain reaction (PCR) machine using M-MLV reverse transcriptase kit from Invitrogen, whereas for miRNA expression, TransScript miRNA First-Strand cDNA Synthesis SuperMix (TransGen Biotech, Beijing, China) was selected to synthesize the first strand of cDNA. Quantitative real-time PCR (qRT-PCR) was carried out by TransStart Top Green qPCR SuperMix (TransGen Biotech). The quantitative results were standardized by the formula (2−ΔΔCt) with the endogenous reference of GAPDH or U6. All primers were purchased from Sangon Biotech (Shanghai, China) and sequences were listed as follows: XIST, GCATAACTCGGCTTAGGGCT (sense) and TCCTCTGCCTGACCTGCTAT (antisense); miR-889-3p, ACACTCCAG CTGGGTTAATATCGGACAAC (sense) and TGGTGTCGTGGAGTCG (antisense); SIX1, TTACGCAGGAGCAAGTGGCG (sense) and CGCTCTCGTTCTTGTGCAGG (antisense); GAPDH, AGGTGAAGGTCGGAGTCAAC (sense) and CGCTCCTGGAAGATGGTGAT (antisense); U6, GTGCTCGCTTCGGCAGCACAT (sense) and TACCTTGCGAAGTGCTTAAAC (antisense).
Cell Counting Kit-8 assay
The determination experiment of cell proliferation was measured by using cell counting kit-8 (CCK-8) method. Me180 and Hela cells with a density of 1 × 104 cells per well were placed into plates and incubated at 37°C for 48 h. Then, Me180 and Hela cell lines were added with CCK-8 reagent for another 2-h incubation. Finally, a microplate reader (Bio-Rad, Hercules, CA) was used to test the absorbance of Me180 and Hela cells at the 450 nm wavelength.
Transwell assay
Both the number of migrated and invaded CC cells were evaluated by transwell migration and invasion assays. In brief, the transfected Hela and Me180 cells were transferred to the upper chamber (Corning, New York, NY) for detection of cell migration, whereas for cell invasion, the upper surface of the chamber was covered by Matrigel (BD Biosciences, Sparks, MD) in advance. All the lower chambers were added with 10% fetal bovine serum medium. After cultivation for 48 h, Me180 and Hela cells located at the upper layers of chambers were wiped with a cotton bud and the cells that passed through the membranes were mixed with crystal violet (Sigma, St. Louis, MO). The countess automatic cell counter (Invitrogen) was used to detect the number of migrated and invaded cells with at least three random field vision.
Flow cytometry assay
Transfected Hela and Me180 cells were seeded into plates and then were put in an incubator for 48 h. All transfected cells were harvested and washed three times using phosphate-buffered saline (PBS). Next, transfected cells were suspended in binding buffer. Annexin-V fluorescein isothiocyanate (FITC) reagent and propidium iodide (PI) solution were added into cell suspension. After 10 min incubation in the dark, the apoptosis of transfected cells was determined by FACScan® flow cytometry (BD Biosciences, San Jose, CA).
Murine xenograft assay
Nude mice were obtained from Tengxin Company (Chongqing, China). To generate the sh-XIST mice model, Me180 cell line was infected with lentivirus harboring shRNA (sh-XIST or sh-NC). Then, all nude mice (n = 5 for each group) were injected with 1 × 106 transfected Me180 cells. Subsequently, the tumor length and tumor width were detected by ruler every 7 d. The tumor volume was calculated by the formula: width 2 × length/2. At 28 d post-inoculation, all mice were killed, and tumors were taken for further experiment. All animal experiments were approved by the Animal Research Committee of The first Affiliated Hospital of Hebei North University and were in line with the guidelines of the National Animal Care and Ethics Institution.
Dual-luciferase reporter assay
The targets of XIST and miR-889-3p were analyzed by LncBase Predicted v2 or micro-T CDS website tool, respectively. Based on the predicted results, the partial sequences of XIST WT and SIX1 WT containing the binding sites were amplified and inserted into the luciferase vector pmirGLO (Promega, Madison, WI). In addition, the mutant types of these were designed and synthesized from Sangon Biotech (Sangon). All luciferase vectors (empty, XIST WT, XIST MUT, SIX1 3′-UTR WT, and SIX1 3′-UTR MUT) were constructed and transfected into Hela and Me180 cells with miR-NC or miR-889-3p. At 48 h post-transfection, a dual-luciferase reporter assay kit (Promega) was used to examine the luciferase activities of transfected cells.
RNA immunoprecipitation assay
The enrichment of XIST and SIX1 was determined by Magna RNA immunoprecipitation (RIP) kit (Millipore, Billerica, MA). After CC cells were transfected with miR-NC or miR-889-3p for 48 h, RIP buffer was added into the transfected cells, and then the cell lysate interacted with magnetic beads covered with antibody Ago2 (ab32381; Abcam, Cambridge, MA) or IgG (ab6721; Abcam). Finally, the enrichment of XIST and SIX1 was determined using qRT-PCR assay.
Western blot assay
Transfected Me180 and Hela cells were collected and transferred into the radioimmunoprecipitation assay buffer (RIPA; Beyotime, Shanghai, China). Then, total proteins were extracted based on the standard protocol. Agarose gel was prepared to test the quality of proteins, and NanoDrop 2000 (Thermo Fisher Scientific) was used to quantify total proteins. Next, equal amount of proteins was loaded into the lanes of freshly prepared sodium dodecyl sulfate–polyacrylamide gel electrophoresis for separating the protein refers to the molecular weight of proteins. After blotting proteins onto polyvinylidene difluoride membranes (Millipore), the membranes were added with 5% nonfat milk for 1-h blocking. Then, the membranes were mixed with antibody I against SIX1 (1:1000, ab86028; Abcam, Cambridge, United Kingdom) or β-actin (1:1000, ab8227; Abcam) for overnight incubation. The next day, the membranes were washed with PBS at least five times and then added with antibody II (ab6721; Abcam) for 2 h. Finally, the visualized proteins by commercial enhanced chemiluminescence chromogenic substrate (Beyotime) were analyzed by Image Lab software (Bio-Rad).
Statistical analysis
All results were presented as mean ± SD from three independent experiments. Student's t-test was used to analyze the comparison between two groups, whereas the comparison among at least three groups was analyzed by one-way analysis of variance (ANOVA). The difference was considered statistically significant at p < 0.05.
Results
XIST was highly expressed in CC tissues and cells
First, the expression of XIST was detected by qRT-PCR assay in CC tissues and cells. The results indicated that XIST expression was largely increased in CC tissues compared with normal tissues (Fig. 1A). In addition, Me180 and Hela cells had the higher expression of XIST than Ect1/E6E7 cell (Fig. 1B). All CC patients were divided into two groups (stage I-II and stage III-IV), and qRT-PCR assay was conducted to measure the expression of XIST. The data showed that XIST expression was enhanced in stage III-IV patients relative to patients at stage I-II (Fig. 1C). Besides, patients with low XIST expression survived longer than that with high XIST expression (Fig. 1D). These data implied that XIST was involved in the development of CC.
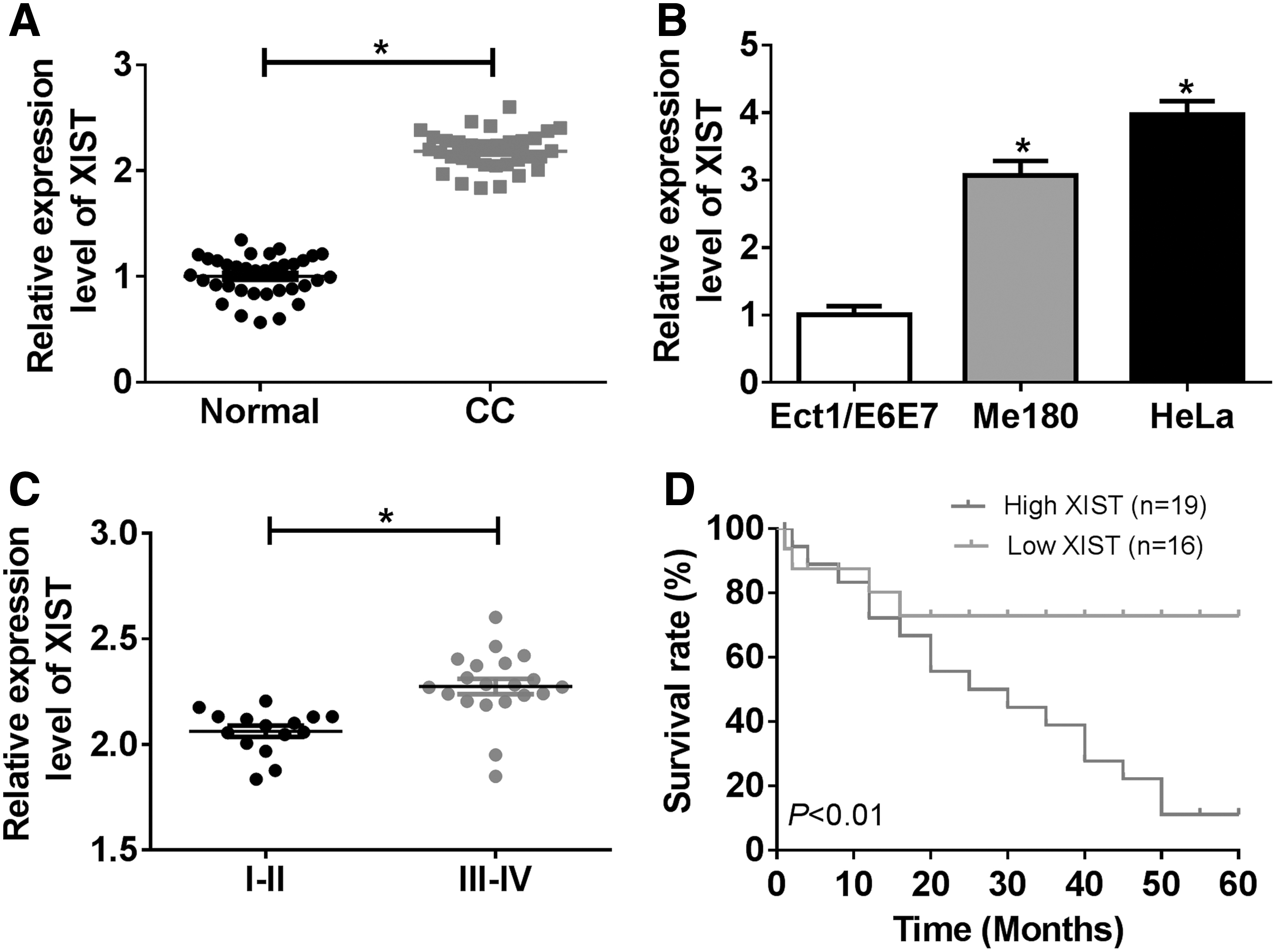
The expression of XIST was increased in CC patients and cells.
Silenced XIST inhibited proliferation, migration, and invasion and induced apoptosis of CC cells
To explore the function of XIST in CC, Me180 and Hela cells were transfected with si-XIST or si-NC. QRT-PCR assay showed that the expression of XIST was remarkably decreased by XIST knockdown in Me180 and Hela cells and the lowest expression of XIST induced by si-XIST#3 was found among these three interference RNAs (Fig. 2A). CCK-8 assay indicated that the proliferation of Me180 and Hela cells were blocked by the knockdown of XIST (Fig. 2B, C). Cell migration and invasion were evaluated by transwell assay. The results determined that both migrated and invaded cells were significantly reduced by XIST silencing in Me180 and Hela cells (Fig. 2D, E). Moreover, flow cytometry assay manifested that the apoptosis of Me180 and Hela cells was promoted by transfection with si-XIST (Fig. 2F). All the above-mentioned data meant that silenced XIST hindered proliferation, migration, and invasion and triggered apoptosis of CC cells.
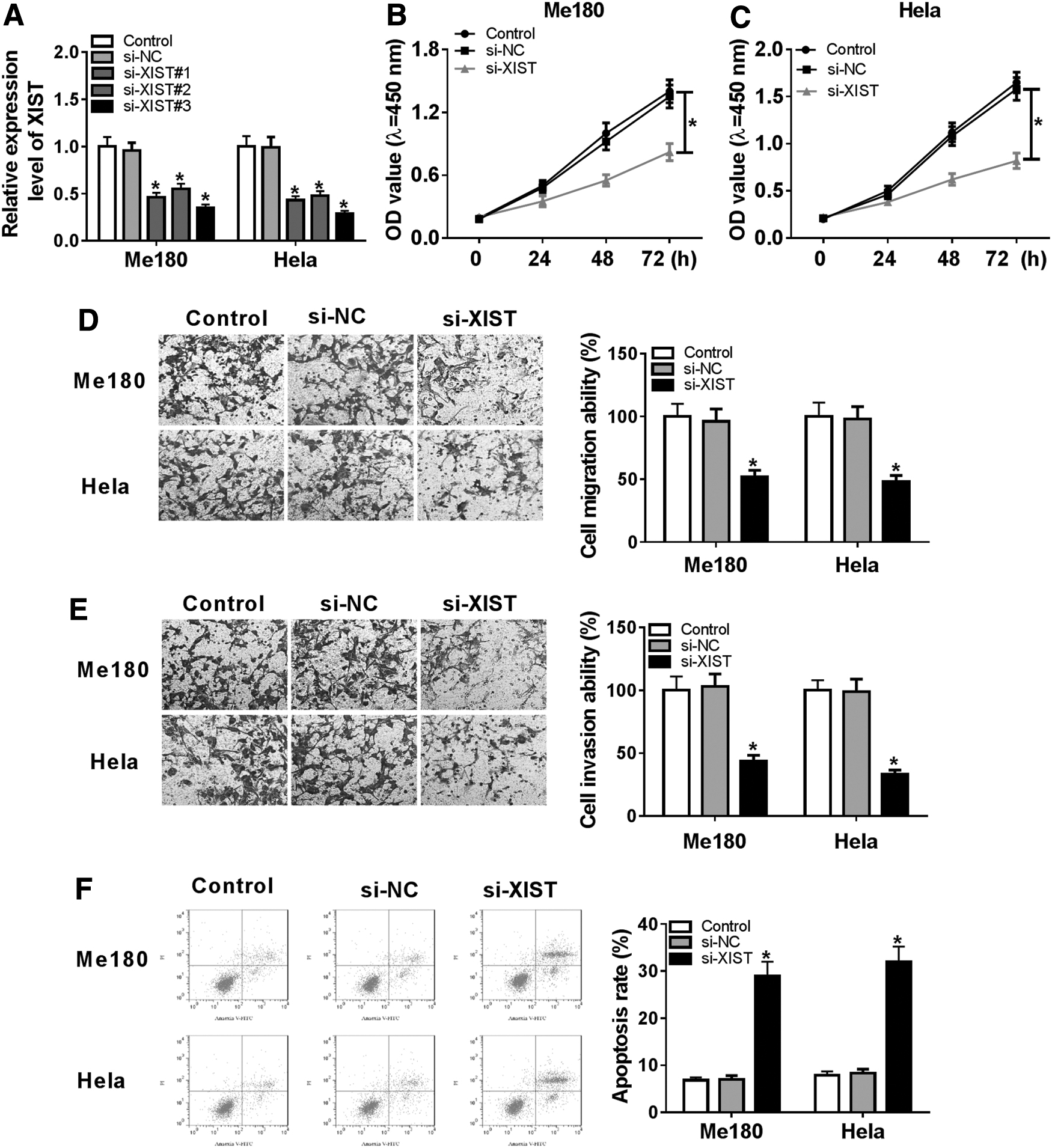
XIST knockdown hindered the proliferation, migration, and invasion and induced apoptosis.
XIST knockdown impeded the tumor growth in vivo
To illuminate the role of XIST in vivo, nude mice models were generated by injection with sh-NC or sh-XIST. Tumor volume and weight were both drastically reduced by XIST silencing (Fig. 3A, B). In addition, qRT-PCR assay revealed that the expression of XIST was obviously decreased by the downregulation of XIST (Fig. 3C). The in vivo experiment suggested that XIST silencing could inhibit tumor growth.
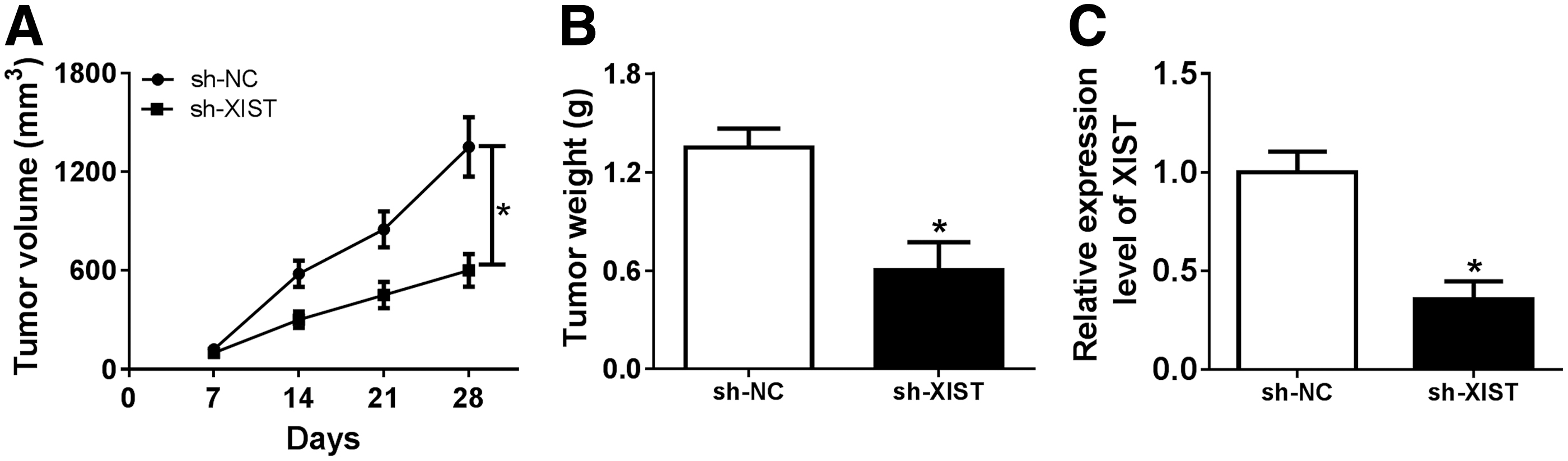
XIST downregulation inhibited tumor growth in vivo.
XIST directly bound to miR-889-3p and inversely regulated miR-889-3p expression
To expound the relationship between XIST and miR-889-3p, LncBase Predicted v2 tool was conducted to predict the binding sites and the mutant sequences of XIST were designed to confirm the binding activity (Fig. 4A). Dual-luciferase reporter assay indicated that miR-889-3p significantly limited the luciferase activities of Me180 and Hela cells in XIST WT group compared with the negative control, whereas for empty vector and XIST MUT groups, luciferase activities remained unchanged (Fig. 4B, C). Furthermore, RIP assay showed that the introduction of miR-889-3p markedly increased the enrichment of XIST in RIP–Ago2 complex relative to RIP–IgG complex in both Me180 and Hela cells (Fig. 4D). Besides, qRT-PCR assay demonstrated that miR-889-3p expression was decreased by XIST upregulation and was promoted by transfection with XIST knockdown (Fig. 4E). In total, XIST was directly interacted with miR-889-3p and negatively regulated miR-889-3p expression.
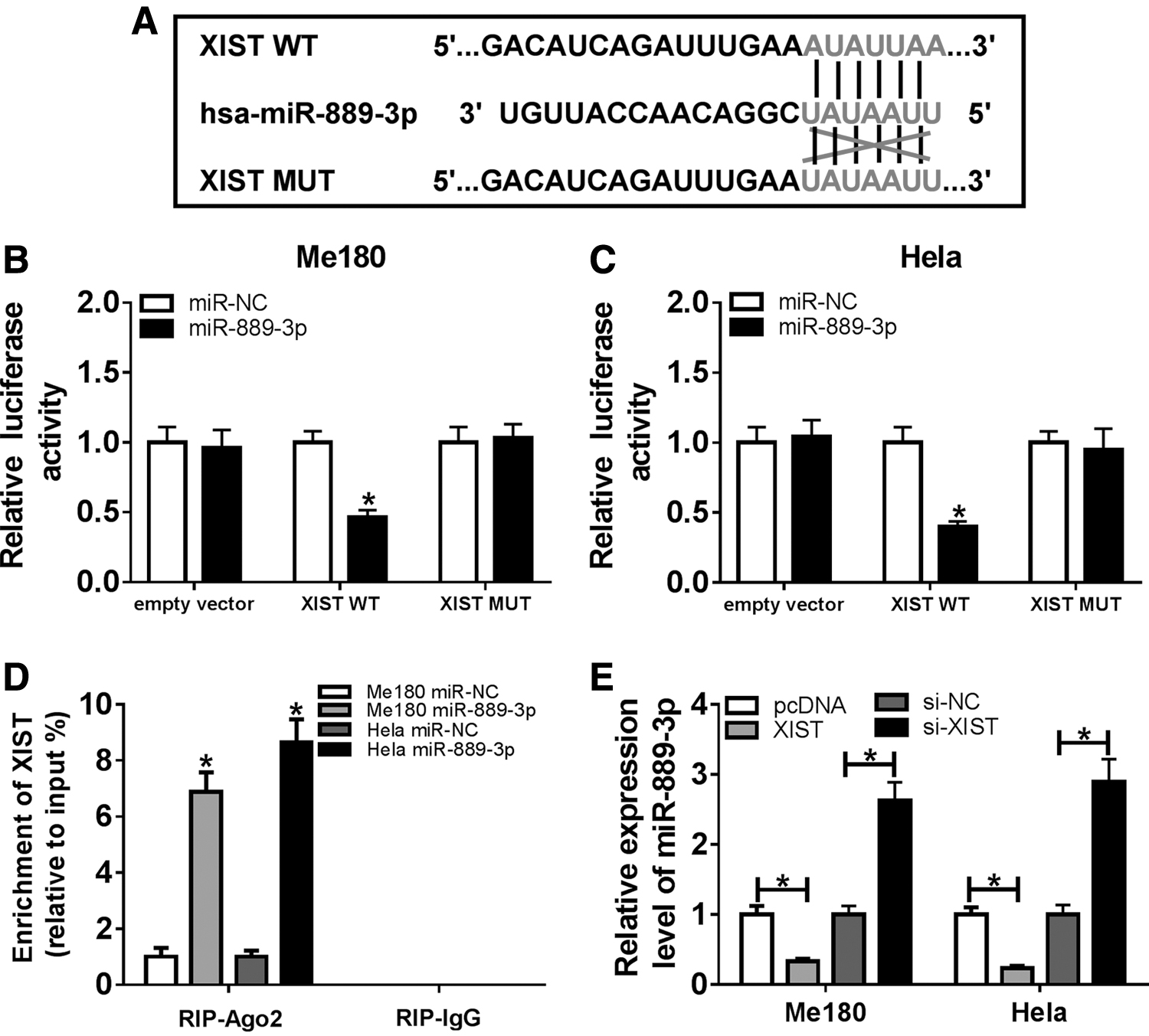
XIST targeted miR-889-3p and negatively regulated miR-889-3p expression.
XIST repressed proliferation, migration, and invasion and facilitated apoptosis by suppressing miR-889-3p expression
To explore the interaction between XIST and miR-889-3p, the expression of miR-889-3p was detected by qRT-PCR assay. The results revealed that the expression of miR-889-3p was enhanced in Me180 and Hela cells transfected with miR-889-3p, whereas XIST overexpression reduced miR-889-3p level by transfection with XIST (Fig. 5A). CCK-8 assay determined that miR-889-3p hindered the proliferation of Me180 and Hela cells, which was abated by XIST upregulation (Fig. 5B, C). Moreover, transwell assay indicated that both migration and invasion abilities of Me180 and Hela cells were reduced by miR-889-3p and were promoted by cotransfection with XIST (Fig. 5D, E). In addition, the apoptosis of Me180 and Hela cells was triggered by miR-889-3p, which was abrogated after transfection with XIST (Fig. 5F). In a word, XIST reversed the effects of miR-889-3p on proliferation, migration, invasion, and apoptosis of CC cells.
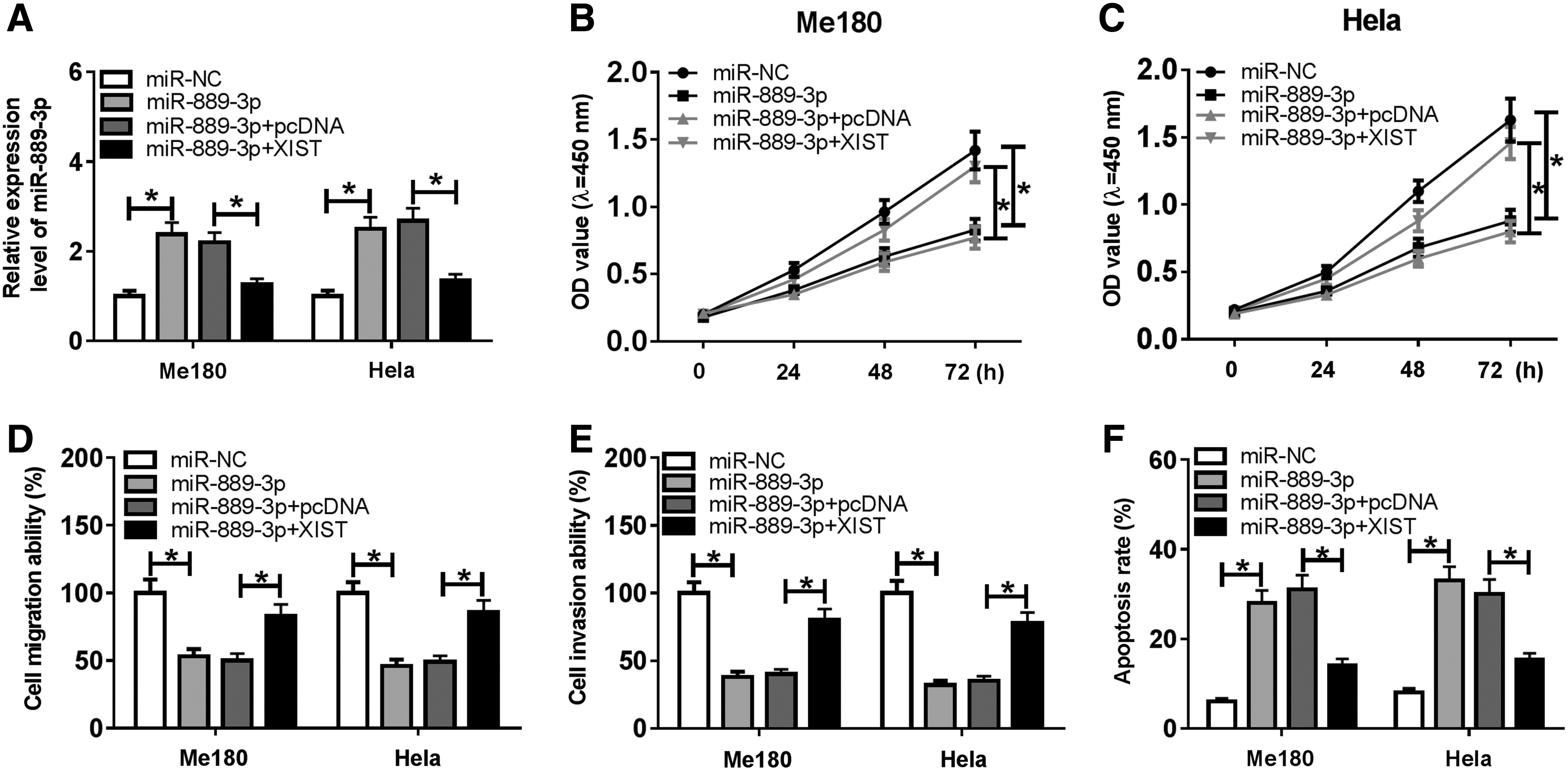
XIST promoted the proliferation, migration, and invasion and blocked apoptosis by sponging miR-889-3p.
SIX1 was a target of miR-889-3p
Micro-T CDS online tool was performed to predict the potential target of miR-889-3p and revealed that SIX1 contains the binding sites of miR-889-3p (Fig. 6A). To verify the prediction, luciferase vectors (SIX1 3′-UTR WT and SIX1 3′-UTR MUT) were constructed. Dual-luciferase reporter assay demonstrated that luciferase activities of Me180 and Hela cells were sharply decreased by cotransfection with miR-889-3p and SIX1 3′-UTR WT compared with miR-NC control, whereas no obvious changes were observed in the empty vector and SIX1 3′-UTR MUT groups (Fig. 6B, C). Of note, the enrichment of SIX1 in RIP–Ago complex was enhanced by miR-889-3p relative to miR-NC, whereas there was no difference in RIP–IgG complex (Fig. 6D). In addition, the protein level of SIX1 was measured by Western blot assay and the results showed that SIX1 protein level was significantly reduced by miR-889-3p upregulation in Me180 and Hela cells (Fig. 6E). To sum up, miR-889-3p targeted SIX1 and repressed SIX1 expression.
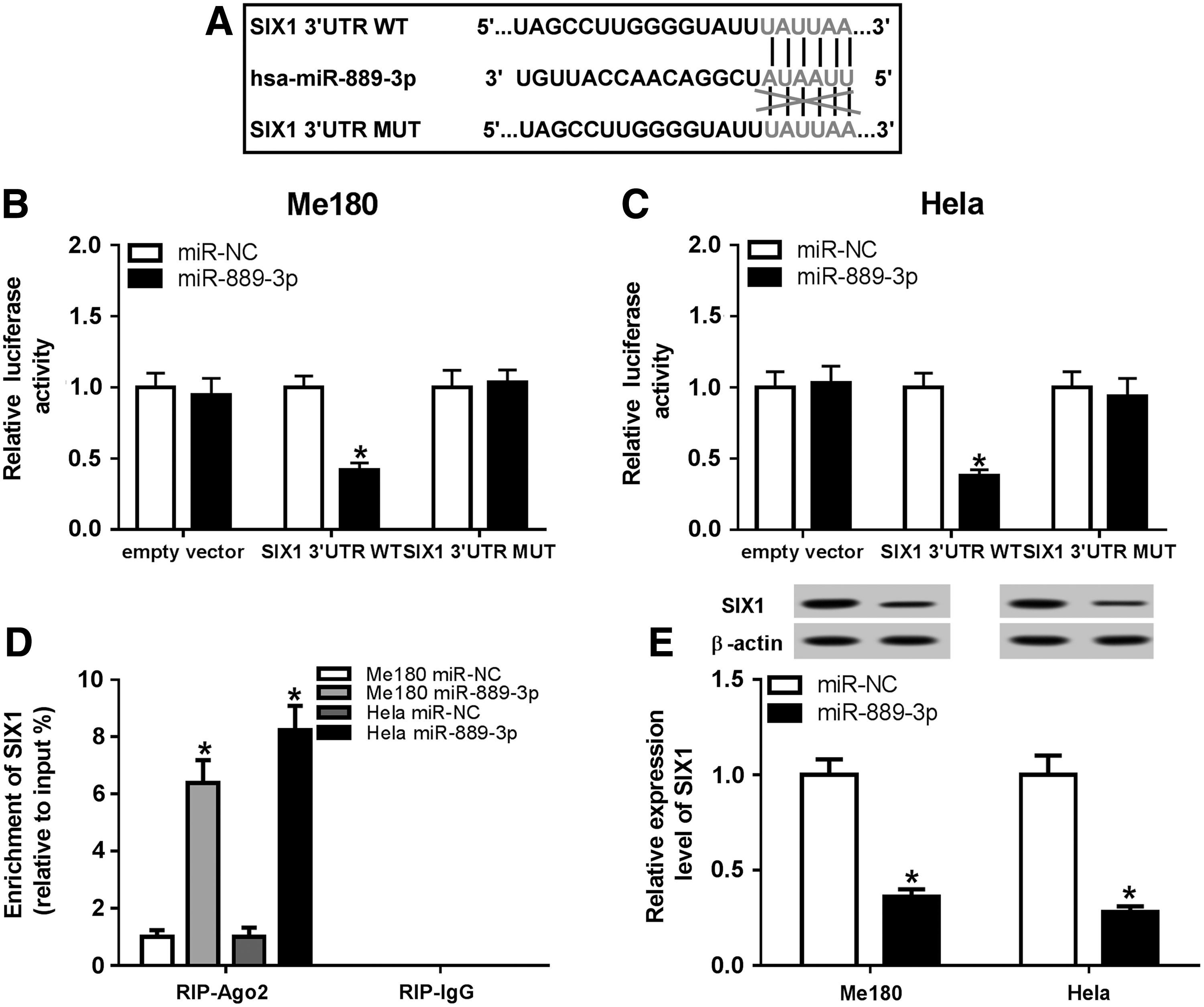
MiR-889-3p targetedly regulated SIX1 expression.
MiR-889-3p expression was decreased and SIX1 expression was elevated in CC tissues and cells
To investigate the relationship among XIST, miR-889-3p, and SIX1 in CC, the expression of miR-899-3p and SIX1 was detected by qRT-PCR assay. Compared with the normal tissues, miR-899-3p expression was largely decreased in CC tissues (Fig. 7A). In addition, the expression of miR-889-3p was reduced in Me180 and Hela cells relative to Ect1/E6E7 cell (Fig. 7B). Based on the expression of XIST measured previously, a negative correlation between XIST and miR-889-3p was found by Pearson analysis (Fig. 7C). Meanwhile, SIX1 level was prominently enhanced in CC tissues (Fig. 7D). In addition, a higher level of SIX1 was observed in Me180 and Hela cells relative to Ect1/E6E7 cell (Fig. 7E). Of interest, miR-889-3p expression was inversely correlated with SIX1 expression (Fig. 7F). Besides, there is a positive correlation between XIST expression and SIX1 expression (Fig. 7G). Western blot assay showed that SIX1 level was reduced by XIST silencing, whereas the interference of miR-889-3p reversed the effect of silenced XIST (Fig. 7H). Collectively, downregulation of miR-889-3p and upregulation of SIX1 were found in CC tissues and cells and there were correlations among XIST, miR-889-3p, and SIX1.
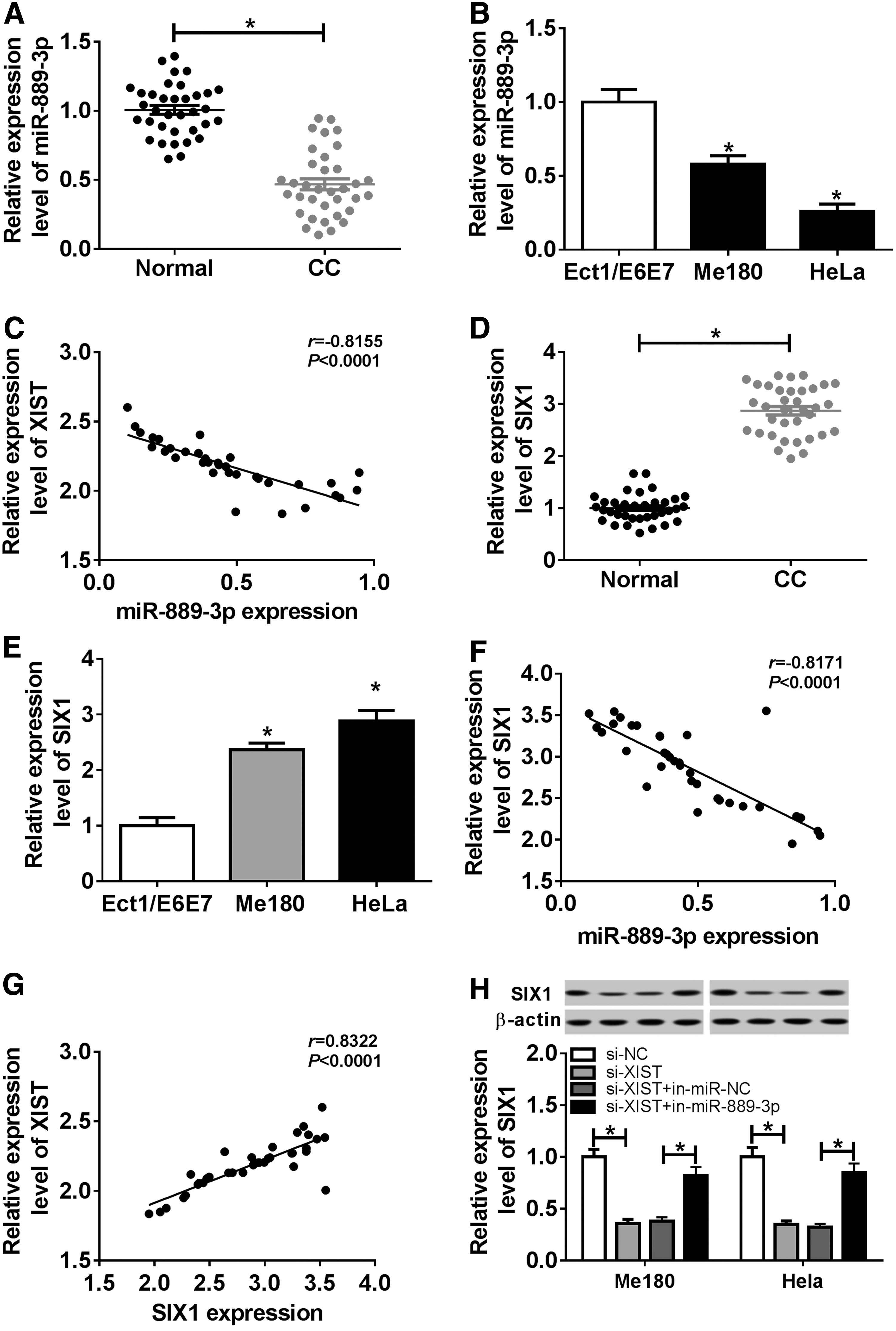
MiR-889-3p was lowly expressed and SIX1 was highly expressed in CC tissue and cells.
MiR-889-3p inhibition abrogated si-SIX1-mediated antiproliferation, antimigration, anti-invasion and proapoptosis of CC cells
To make clear the functional effect of SIX1 in CC, the level of SIX1 was detected by Western blot assay. The results showed that SIX1 level was decreased by SIX1 knockdown and was promoted by transfection with miR-889-3p inhibition (Fig. 8A). CCK-8 assay displayed that silenced SIX1 retarded the proliferation of Me180 and Hela cells, which was relieved by miR-889-3p downregulation (Fig. 8B, C). Furthermore, transwell assay indicated that both migration and invasion abilities of Me180 and Hela cells were inhibited by SIX1 silencing and were then promoted by cotransfection with miR-889-3p inhibition (Fig. 8D, E). In addition, silenced SIX1 enhanced the apoptosis of Me180 and Hela cells, which was attenuated by the downregulation of miR-899-3p (Fig. 8F). Overall, miR-899-3p weakened the activities of proliferation, migration, invasion, and apoptosis caused by SIX knockdown in CC.

The interference of miR-889-3p enhanced the proliferation, migration, and invasion and repressed apoptosis by targeting SIX1.
Discussion
The incidence of CC is in second place in women tumors, as one of the most frequent cancers worldwide. More than 10% of all cancer deaths were caused by CC. 23 High metastasis and easy recurrence largely reduced the overall survival of CC patients. Unfortunately, less than half of patients with stage III or IV could survive >5 years, even after standardized treatments. Besides, lncRNAs have been reported to be related to the pathogenesis of CC. 24 Hence, it is essential to elucidate the mechanism of lncRNA XIST in CC.
As the first identified noncoding gene from XIC, XIST served as a predictor of cancer prognosis, which was expressed aberrantly in many human cancers. Chen and his colleagues disclosed that the upregulation of XIST was observed in CC and the downregulation of XIST repressed cell proliferation in vitro and in vivo. 13 Zhu et al. also found that XIST expression was increased and silenced XIST impeded cell invasion and EMT in CC. 25 Similarly, XIST expression in CC patient tissues and cells was determined. Results revealed that highly expressed XIST in CC tissues and cells was discovered and XIST knockdown inhibited cell progression of CC cells, which were consistent with previous studies. Moreover, the in vivo experiment further demonstrated the functional effects of XIST in CC. These data implicated that XIST was negatively associated with cell progression and tumor growth in CC.
MiR-889 was widely studied in various cancers, including CC. A recent study demonstrated that the expression of miR-889 was decreased in CC, whose expression was closely related to cancer staging and metastasis, and miR-889 significantly reduced cell viability and invasion. 26 In this study, miR-889-3p with lower expression in CC tissues and cells was confirmed. These results were in line with our study. Of interest, miR-889-3p was targeted by lncRNA XIST, which was confirmed by the dual-luciferase reporter and RIP assays, and miR-889-3p expression was inversely regulated by XIST. More importantly, XIST could recover the inhibitory effects on cell growth and metastasis and promotional effect on the apoptotic rate of CC cells mediated by miR-889-3p. All these data suggested that XIST contributed to the biological behaviors and the progression of CC by suppressing miR-889-3p expression.
Increasing researches have determined that SIX1 overexpression enhanced cell motility, induced EMT, and led to malignant conversion. 27 Furthermore, Xu et al. reported that SIX1 expression was elevated and SIX1 promoted progression and EMT in CC. 28 In this study, we found the upregulation of SIX1 level in CC tissues and cells, showing no difference with the recent study. Besides, bioinformatics analysis showed that SIX1 was a target of miR-889-3p, and both dual-luciferase reporter and RIP assays verified the combination between them. In addition, miR-889-3p negatively modulated SIX1 expression. Moreover, miR-889-3p inhibition recuperated the activities of SIX1 knockdown on cell growth and metastasis, as well as cell apoptosis. Overall, miR-889-3p suppressed SIX1 expression to regulate cell progression in CC.
Of note, we also found that silenced XIST reduced SIX1 expression through targeting miR-889-3p, suggesting that XIST could regulate miR-889-3p/SIX1 axis to influence cell progression in CC. However, only XIST silencing experiment was conducted in mice models, and there was no more in vivo evidence to explain the interaction among XIST/miR-889-3p/SIX1 axis in CC. Therefore, more researches in the model or even in clinical patients are needed in the future.
Conclusion
XIST and SIX1 were upregulated, whereas miR-889-3p was lowly expressed in CC tissues and cells. XIST silencing blocked cell growth and metastasis and exerted a promotion effect on cell apoptosis in vitro and reduced tumor growth in vivo. In addition, XIST bound to miR-889-3p and negatively regulated miR-889-3p expression. XIST weakened the effects of miR-889-3p on cell growth, apoptosis, and metastasis. Furthermore, miR-889-3p targeted SIX1 and suppressed SIX1 expression. MiR-889-3p abolished SIX1 knockdown-mediated inhibition of cell growth and metastasis and the promotion of cell apoptosis.
Authors' Contribution
X.L.: project development, data collection, data analysis, and article writing; S.X.: project development, data collection, data analysis, and article editing; J.Z.: data collection and data analysis; Y.K.: project development, funding acquisition, data analysis, and article editing.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.