
Abstract
Background:
The incidence of thyroid cancer has increased dramatically in recent decades due, in large part, to identifications of subclinical diseases. Literature on thyroid cancer has examined the pathogenesis of high invasive papillary thyroid cancer (PTC) and has improved the prevention and treatment of PTC. This study aims to investigate the effects of metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) on PTC migration and invasion, and clarify the regulatory mechanisms between miR-146b-5p and MALAT1.
Materials and Methods:
In this study, we examined the differential expression of MALAT1, miR-146b-5p, and DNA methyltransferases 3A (DNMT3A) in PTC tissues. The effect of MALAT1 on the proliferation and invasion ability of PTC cells was verified by constructing a sh-MALAT1 knockdown cell model. Correlations between MALAT1, miR-146b-5p, and DNMT3A were analyzed by the Pearson correlation method. Finally, we verified the regulatory relationship between miR-146b-5p and MALAT1 by the luciferase assay and rescue assay.
Results:
The expression of MALAT1 was upregulated in PTC tissues and cells, while a MALAT1 knockdown counteracted cellular activity, migration, and invasion of B-CPAP and K1 cells. The relationship between miR-146b-5p and DNMT3A was negative, while the relationship between miR-146b-5p and MALAT1 was positive. Both genes were separately detected using the Pearson correlation method. The luciferase assay and rescue assay demonstrated that a binding site in miR-146b-5p was existent in the 3′ untranslated region of DNMT3A, while a knockdown of DNMT3A partially rescued si-miR-146b-5p induced proliferation, migration, and invasion effects on PTC cells.
Conclusions:
The MALAT1 gene is highly expressed in PTC, while the knockdown MALAT1 gene attenuates the cellular activity and invasive ability of PTC cells. The microRNA miR-146b-5p can promote a MALAT1 expression by negatively regulating DNMT3A in PTC.
Introduction
The incidence of thyroid cancer has increased dramatically in recent decades due in large part to identifying subclinical diseases. 1 According to the latest data, thyroid cancer accounts for 5% of new cancer tested in women, ranking fifth in all kinds of cancers. 2 Papillary thyroid cancer (PTC) is the most common subtype of thyroid cancer and carries the best overall prognosis. 3 Some PTC may invade tissues, such as peripheral nerves, blood vessels, and muscles. This part of PTC has a poor prognosis and may recur, metastasize, and cause death in some cases. 4 Studying the pathogenesis of highly invasive PTC will contribute to the diagnosis and treatment of such cancer.
The use of noncoding RNA (ncRNA) as a diagnostic marker has become a hot research topic in the field of PTC. MicroRNA (miRNA) and long noncoding RNA (lncRNA)—two types of ncRNA—play important roles in biological processes, such as cell differentiation, signal transduction, cell growth, and tumorigenesis. 5,6 In thyroid tumors, studies have found that a variety of miRNAs are differentially expressed in benign and malignant thyroid nodules, such as miR-221 and miR-222 in the expression of patients with malignant PTC. 7,8 In the preliminary study of this project group, 51 specimens of thyroid tumor tissue were detected by miRNA microarray and the quantitative real-time polymerase chain reaction (qRT-PCR) technique. It was found that miR-146b-5p was significantly elevated in PTC patients with extracapsular invasion and cervical lymph node metastasis, with the stronger the invasiveness, the more obvious its expression. 9 The high expression of miR-146b-5p has high sensitivity and specificity in thyroid cancer and can be used as an independent and poor prognostic risk assessment factor for thyroid cancer. 10 We used bioinformatics technology to predict that miR-146b-5p can target oncogene metastasis-associated lung adenocarcinoma transcript 1 (MALAT1), one of the earliest discovered lncRNAs. 11 Some studies suggest that MALAT1 affects cancer invasion, metastasis, and epithelial–mesenchymal transition (EMT) by affecting a gene's expression. 12 In thyroid cancer, MALAT1 has been identified as a contributing factor to cancer development and can be used as a novel biomarker for early diagnosis and even treatment. 13
miRNAs and lncRNA can regulate and target gene expression at the transcriptional and post-transcriptional levels through target gene sequence complementarity or epigenetic regulation. 14,15 We speculate that the expression of miR-146b-5p and lncRNA MALAT1 may be involved in the regulation, progression, invasion, and metastasis of PTC. As one of the DNA methylases, DNA methyltransferases 3A (DNMT3A) can modulate the DNA methylation by directing to specific gene promoters and participating in the epigenetic regulation of lncRNA by miRNA. 16 In cancer, DNMT3A was identified as a crucial regulator of metastasis-related genes in gastric cancer. 17 Zheng et al. have reported that hepatitis B virus X protein (HBx) can directly interact with DNMT3A to promote or inhibit the binding of DNMT3A to the promoter region of oncogenes and tumor suppressors. 18 In this study, we first examined the role of MALAT1 in PTC and then used gene editing techniques and luciferase assays to verify its possible regulatory correlation with miR-146b-5p in vitro.
Materials and Methods
Patients and specimens
From March 2017 to February 2018, we collected PTC tissues and paracancerous tissue specimens from the surgical patients in Affiliated Hangzhou First People's Hospital. Both cancer tissues and normal tissue specimens were confirmed by postoperative pathologies; after the examination of both tissue samples, a group detection was performed. Each tissue sample was quickly frozen, stored in liquid nitrogen, and subjected to quantitative RT-PCR analysis and a Western blot analysis. All patients included in this study did not receive any chemotherapy before surgery. Each patient provided a written informed consent form. A complete clinical pathology along with follow-up data was administered for patients with PTC who provided the specimens. This study was approved by the Ethics Committee of Affiliated Hangzhou First People's Hospital, Zhejiang University School of Medicine.
Cell culture
A human normal thyroid follicular epithelial cell line (Nthy-ori 3-1), and PTC cell lines (TPC-1, B-CPAP, and K1) were purchased from Shanghai Institutes for Biological Sciences and were used for this study. Cell lines were cultured using Dulbecco's modified Eagle's medium (Gibco BRL, NY) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 μg/mL streptomycin (Invitrogen, San Diego). The cells were cultured in a humidified cell culture incubator at 37°C in 5% CO2. All cell lines were passaged for less than 6 months.
Total RNA extraction, reverse transcription, and qPCR
Total RNA from tissues and cells was extracted using TRIzol reagent (Invitrogen). Specimens that qualified for nucleic acid quantification and purity were assayed, while the total RNA was reverse transcribed into cDNA using a Primer-Script one-step RT-PCR kit (Takara, Dalian, China). A real-time quantitative RT-PCR detection was performed using an SYBR Premix Dimmer Eraser kit (Takara) on an ABI 7500 system (Applied Biosystems, Waltham). The expression profile of miR-146b-5p (Assay ID. 001097; Thermo Fisher Scientific, Carlsbad) was examined using TaqMan microRNA assays. miRNA-specific primer sequences were designed and synthesized based on miRNA sequences obtained from the miRBase database: hsa-miR-146b-5p, UGAGAACUGAAUUCCAUAGGCU. Glyceraldehyde 3-phophate dehydrogenase (GAPDH) was used as an internal reference gene, while the mRNA value of the target gene was normalized by comparing it to GAPDH. The primer sequences are presented in Table 1. The 2−ΔΔCt method was used to calculate the relative expression fold change of the target gene. Finally, a statistical analysis was performed.
Primer Sequences
PCR, polymerase chain reaction.
Plasmid construction
The MALAT1 expression plasmid or the miRNA-146-5p seed region binding site mutation plasmid was obtained by a PCR amplification using human genomic DNA as a template. TA cloning is one of the most popular methods of cloning the amplified PCR product using Taq and other polymerases. The PCR product was verified by TA cloning sequencing, subcloned into the mammalian expression vector pcDNA3.1 (Invitrogen) for routine validation, expanding culture, and plasmid extraction. A plasmid transfection of target cells was performed using lipofectamine 2000 (Invitrogen). The promoter region sequence of MALAT1 and DNMT3A were obtained using NCBI, while cloning primers were designed and synthesized. The MALAT1 or DNMT3A promoter truncation mutant plasmid and the binding site point mutation plasmid were cloned using PrimerStar polymerase (Takara), cloned with the pGL3 promoter plasmid as a template and subcloned into the pGL3 luciferase plasmid backbone.
Cell transfection
Both MALAT1 siRNA and negative control siRNA (si-NC) were purchased from Qiagen (Hilden, Germany). Hsa-miRNA-146b-5p mimic, hsa-miRNA-146b-5p inhibitor, and negative control mimic were purchased from Genechem (Shanghai, China). Cells were then plated in a six-well plate for 24 h, and cell transfection was performed according to the appropriate instructions. After 6 h of transfection, the medium was changed to a fresh basal medium, and culture was continued for 48 h. Cells were harvested and subjected to real-time quantitative RT-PCR or a Western blot analysis.
Luciferase experiment
The sequence of wild-type miR-146b-5p (miR-146b-5p-wt) and wild-type 3′ untranslated region (3′-UTR) of DNMT3A mRNA was obtained by NCBI. A mutant type of DNMT3A (DNMT3A-mut) without binding sites with miR-146-5p was obtained via an overlap extension PCR with mutant primers. For the dual-luciferase activity assay, DNMT3A wild-type or mutation type was cloned into the 3′-UTR region of the Renilla luciferase gene of the pRL-TK plasmid. The above plasmids were transfected into cells and co-transfected with the corresponding miRNAs mimics or negative control mimic. Firefly luciferase was used in pGL3-control to monitor transfection efficiency. A luciferase assay was performed according to the dual-luciferase reporter assay kit instructions.
Western blotting
Western blotting was performed according to a conventional method for detecting expression levels of DNMT3A and GAPDH proteins. DNMT3As primary antibody (ab232391; Abcam, Cambridge, United Kingdom) and GAPDH primary antibody (ab181602) were both purchased in Abcam.
Cell migration and invasion experiment
After 48 h of cell transfection, 50,000 cells diluted in serum-free medium were seeded in an upper chamber (8.0 μm; Millipore, Burlington) and further coated with Matrigel (Sigma, St. Louis). The upper chamber was then placed in the bottom chamber of the medium (which contained 10% FBS) for a duration of 24 h. The cells on the upper surface were then lightly scraped off and rinsed, while the lower surface was invaded into the cell fixative and stained with 0.05% crystal violet for 2 h. Finally, the invading cells were counted under a microscope, and the relative numbers were calculated. The experiment was repeated separately three times.
Statistical analysis
All statistical analyses were performed using SPSS, version 19.0 (SPSS, Chicago). The MALAT1 gene expression levels for tumor tissues and adjacent normal tissues were compared using a paired-sample t-test. Other results were analyzed using an independent sample t-test or a one-way ANOVA. All data are expressed as a mean ± standard deviation. p < 0.05 was considered to be statistically significant.
Results
The expression of MALAT1 was upregulated, and DNMT3A was downregulated in PTC tissues and cells
qRT-PCR was used to compare the expression of MALAT1 in cancer tissues and adjacent tissues in 46 patients with PTC. It was found that the expression of MALAT1 in cancer tissues was significantly higher than that in adjacent tissues (Fig. 1A). After the detection of different PTC cell lines, the same results were obtained whereby the expression level of MALAT1 was significantly increased (Fig. 1B). Western blot analysis and qRT-PCR were used to detect the protein and mRNA expression of DNMT3A in the tissues of 46 patients. It was found that the protein and mRNA expression of DNMT3A in PTC tissues was lower than that in adjacent tissues (Fig. 1C, D). In the PTC cell line, furthermore, the mRNA expression level of DNMT3A was also significantly decreased (Fig. 1E).
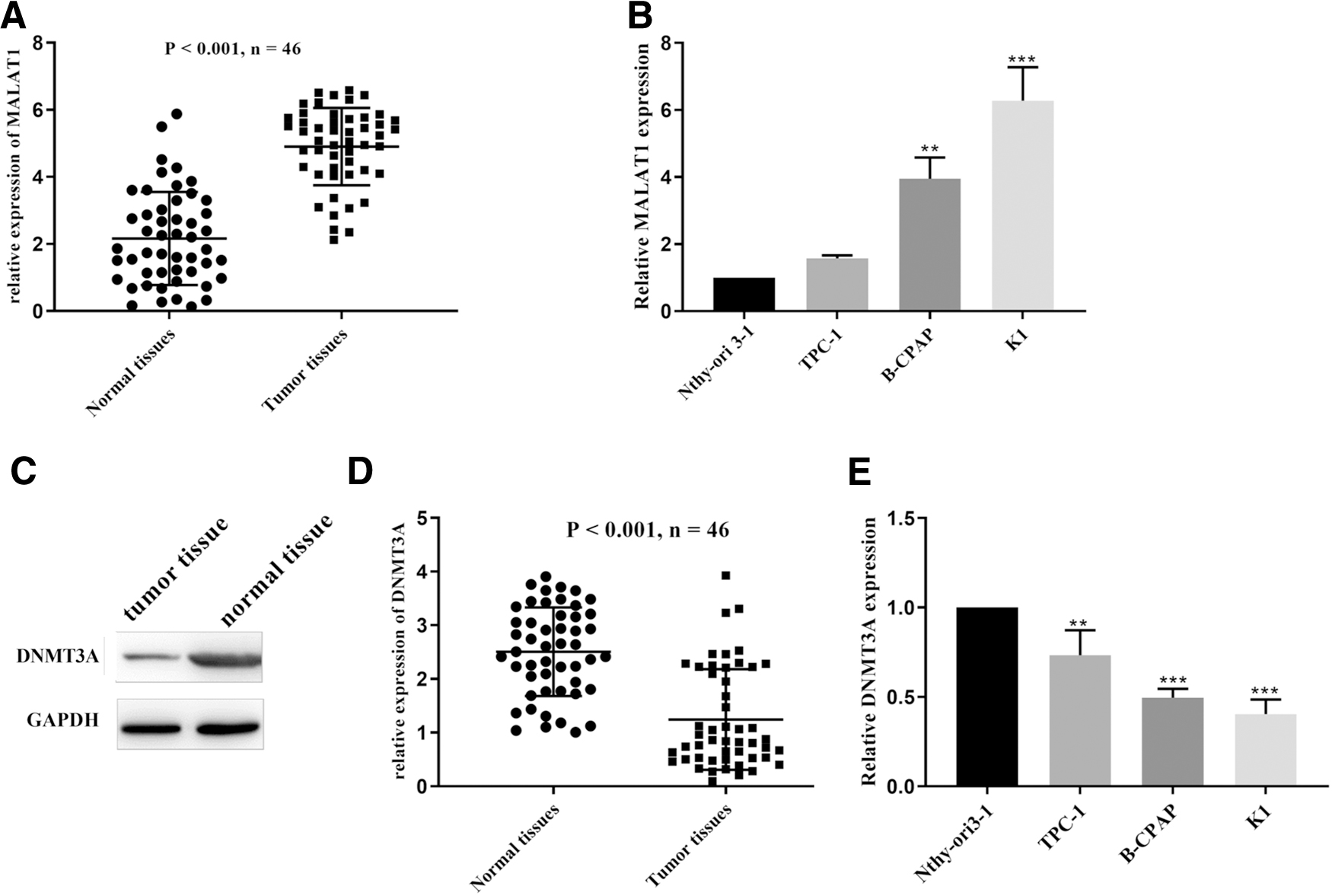
The expression of MALAT1 was upregulated, and DNMT3A was downregulated in PTC tissues and cells.
Knockdown of MALAT1 counteracted proliferation, migration, and invasion of B-CPAP and K1 cells
To further validate the mechanism of MALAT1 in PTC, we selected two cell lines, B-CPAP and K1, for knockout validation. The expression of MALAT1 was reduced in both cell lines by siRNA interference (Fig. 2A). After the expression of MALAT1 was decreased, the cell viability of the two cell lines was significantly reduced, relative to the si-NC group (Fig. 2B, C). The results of the Transwell assay demonstrated that the migration and invasion of B-CPAP and K1 cells were significantly reduced after a knockdown of MALAT1 (Fig. 2D).
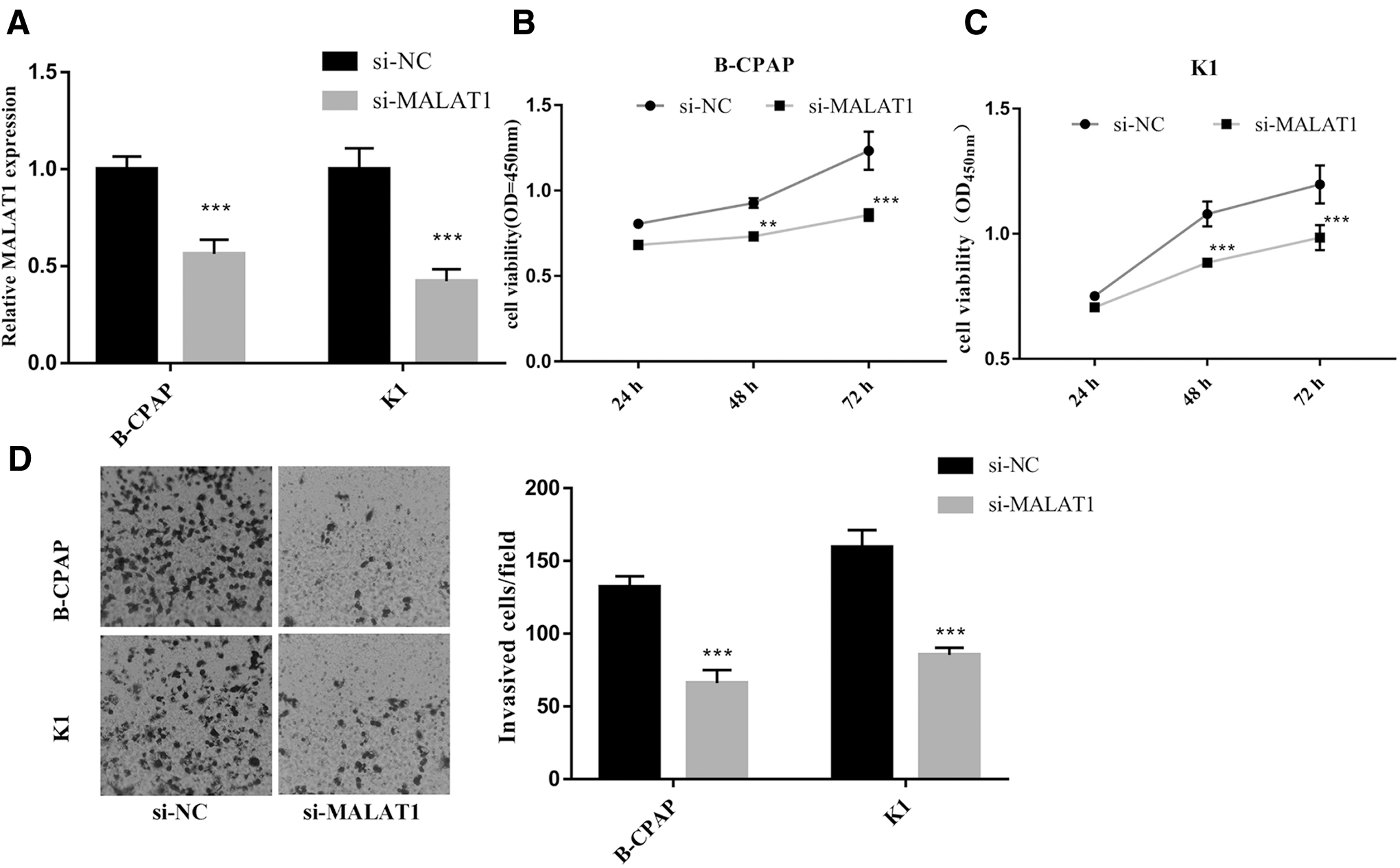
Knockdown of MALAT1 counteracted proliferation, migration, and invasion of B-CPAP and K1 cells. B-CPAP and K1 cells were treated with si-NC and si-MALAT1, respectively. The mRNA level of MALAT1 was detected
MiR-146b-5p was positively associated with MALAT1, and miR-146b-5p could silence the expression of DNMT3A mRNA
The expression level of miR-146b-5p in PTC was significantly higher than that in adjacent tissues (Fig. 3A). After the analysis of the Pearson correlation method, miR-146b-5p showed a negative correlation with DNMT3A and a positive correlation with MALAT1 (Fig. 3B, C). After a bioinformatics analysis was completed, we obtained the binding sites between the wild-type DNMT3A and miR-146b-5p; afterward, we mutated the sequence of DNMT3A given that they could not bind with one another anymore (Fig. 3D). The relationship between miR-146b-5p and DNMT3A target was verified by a luciferase assay. The results showed that miR-146b-5p-mimic attenuated the luciferase activity on the DNMT3A wild-type group in both cell lines, but had no effect on the DNMT3A mutant type group. After the addition of miR-146b-5p-inhibitor, luciferase activity was significantly increased in either DNMT3A-wt or DNMT3A-mut groups (Fig. 3E).
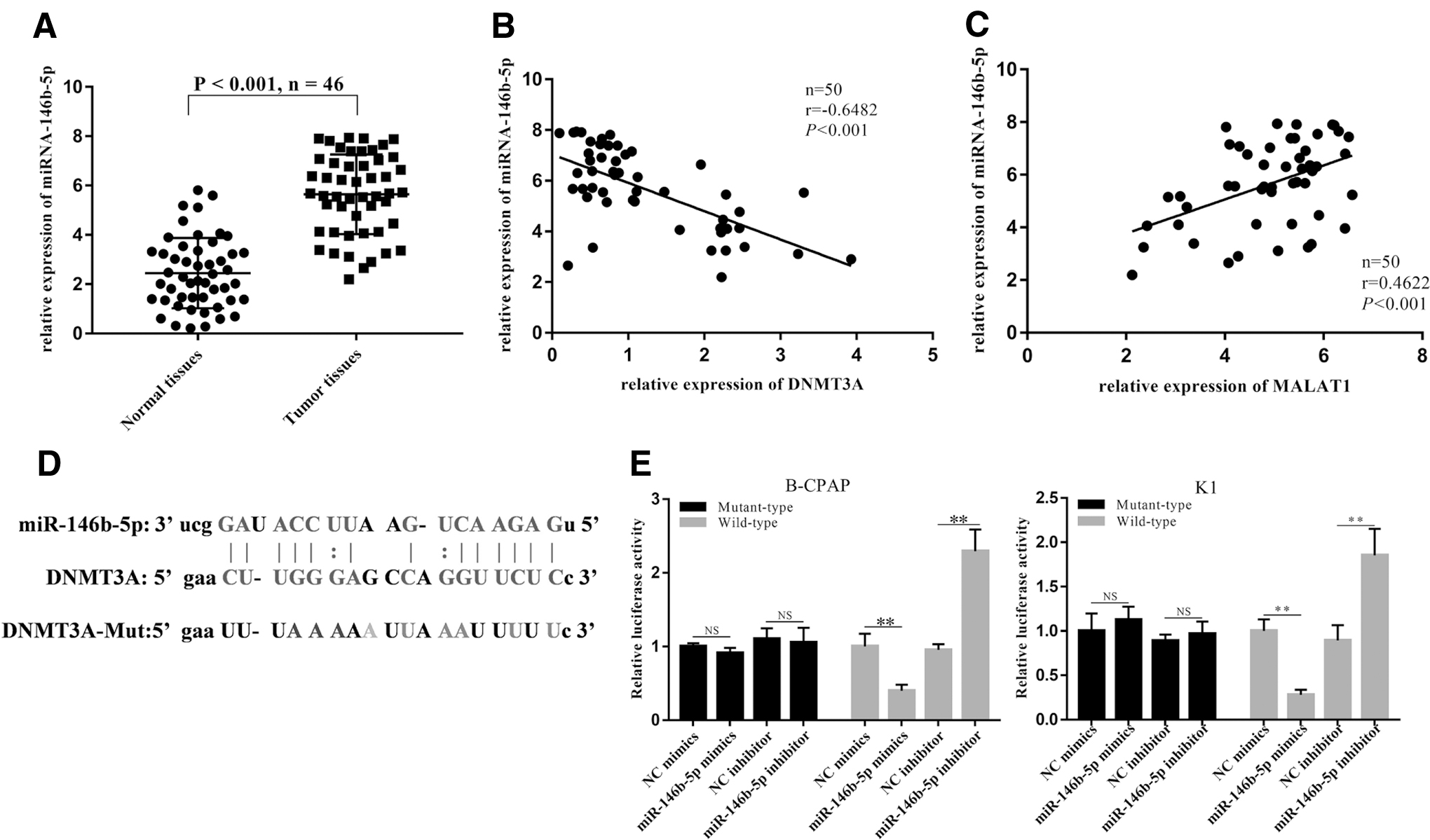
MiR-146b-5p was positively associated with MALAT1, and miR-146b-5p could silence the expression of DNMT3A mRNA.
MiR-146b-5p promoted the expression of MALAT1 by negatively regulating DNMT3A
The rescue experiment was performed to verify the relationship between miR-146b-5p, DNMT3, and MALAT1. The results showed that the expression of MALAT1 was significantly increased after knocking down DNMT3A alone. After knocking down miR-146b-5p alone, the expression of MALAT1 decreased. After knocking down DNMT3A and miR-146b-5p together, the expression of MALAT1 was increased, but it was lower than that of DNMT3A's knockdown alone (Fig. 4A). The cell viability, migration, and invasion ability of the two cell lines were the same tendency as Figure 4A (Fig. 4B–E).

MiR-146b-5p promoted the expression of MALAT1 by negatively regulating DNMT3A.
Discussion
In the present study, we mainly found that miR-146b-5p promoted the expression of MALAT1 by negatively regulating DNMT3A. Combined with our previous studies and bioinformatics predictions, it was found that miR-146b-5p was significantly elevated in patients with PTC and bioinformatics technology predicted that miR-146b-5p may regulate MALAT1. 9 Here, we first determined the high expression of miR-146b-5p, MALAT1, and DNMT3A in 46 PTC tissues. The results showed that the expressions of miR-146b-5p and MALAT1 in cancer tissues were significantly higher than those in adjacent tissues, and the expression levels of DNMT3A were therefore significantly reduced. Then we found that knockdown of MALAT1 counteracted proliferation, migration, and invasion of B-CPAP and K1 cells. After using the Pearson correlation method as a means of analysis, we found the correlations between miR-146b-5p, MALAT1, and DNMT3A. It was then confirmed by a luciferase assay and rescue experiments that miR-146b-5p could regulate the expression of lncRNA MALAT1 and its effect on the invasion and proliferation of PTC by targeting DNMT3A. Given the relevance of PTC in the health care field, our research can potentially provide additional information for practitioners in its treatment.
Due to various causes, from environmental factors to attribution bias, the incidence of papillary thyroid carcinoma is gradually increasing. 19 Differentiated thyroid cancer is the most frequent subtype of thyroid cancer, accounting for more than 95% of cases in America, and PTC is one of these differentiated thyroid cancers, carrying the best overall prognosis. 3,20 In our previous clinical work, some PTCs show higher levels of invasiveness, while pathological examinations have revealed cervical lymph node metastasis and even an invasion of peripheral nerves, blood vessels, and muscles. 21 These PTCs have not been proven to invade thyroid capsules at the time of the initial diagnosis. The misdiagnosis of thyroid papillary carcinoma and the recurrence of tumors can further lead to reoperation. Research on thyroid cancer is pressing and requires further study to clarify its highly invasive nature and improve prevention and treatment.
Two kinds of ncRNA (i.e., miRNA and lncRNA) participate in the physiological and pathological processes of the organism and play an essential role in the occurrence of tumors. 22,23 miRNAs mainly participate in the gene's post-transcriptional regulation by targeting the 3′-UTR of mRNA transcripts. 24,25 LncRNA mainly regulates gene expression from three lenses: epigenetics; transcriptional regulation; and post-transcriptional regulation. 26,27 In thyroid tumors, some research results show that the expression of miR-146b-5p in thyroid cancer was significantly higher than that in adjacent nontumor tissues and benign hyperplastic thyroid nodules. It means that miR-146b-5p can be used for the differential diagnosis of benign and malignant thyroid nodules. 28,29 The overexpression of miR-146b-5p in normal rat follicular cells (PCCL3) decreased SMAD4 (SMAD family member 4/Mothers against decapentaplegic homolog 4) levels and disrupted the transforming growth factor β signal transduction. 29 The expression level of miR-146b-5p is associated with extra-glandular invasion and metastasis of thyroid cancer, and has been significantly higher in invasive thyroid carcinoma than in noninvasive thyroid cancer. 30 The MALAT1 gene is named for its initial discovery in human nonsmall cell lung cancer. Studies have suggested that MALAT1 affects cancer invasion, metastasis, and EMT by affecting gene expression. 12 Recent studies, moreover, have shown that MALAT1 could upregulate the expression of IQGAP1; while IQGAP1 knockdown has reversed the decreasing cell proliferation and invasion of thyroid cancer. 31 Our study found a high expression of MALAT1 in PTC along a positive correlation between miR-146b-5p with MALAT1.
There is a regulation relationship between miRNA and lncRNA, which plays an important role in tumor development and treatment. miRNAs target a large number of protein-coding genes and also target lncRNA. 32 miRNA is able to regulate lncRNA expressions through RISC (RNA induced silencing complex), much like how miR-21 can inhibit lncRNA-GAS5. 33 The competing endogenous RNA hypothesis suggests that lncRNA can act as endogenous miRNA target mimics, directly binding to miRNAs, inhibiting the regulation of miRNAs on mRNAs, and thereby allowing mRNAs to increase content. 34 Cesana et al. have demonstrated that lncRNA-MD1 can competitively bind to miR-133 and miR-135 to inhibit the interaction of miR-133 and miR-135 with target genes, thereby regulating the expression of MAML1 and MEF2C in human and murine muscles. 35 miRNAs also positively regulate lncRNA through epigenetic regulation. For example, miR-29a overexpressed in liver cancer, increases lncRNA-MEG3 expression by regulating DNA methyltransferases 1 and 3 (DNMT1 and DNMT3). 15 The miR-146-5p in this study is similar to this, and regulates the expression of MALAT1 by regulating DNMT3A. Studies have indicated that miRNAs activate gene transcription epigenetically as an enhancer trigger. 36 The role of lncRNA and miRNA in cell differentiation and signal transduction is a research hotspot in the postgene era.
In summary, this study investigated the effect of miR-146b-5p on the expression of lncRNA MALAT1 and its effect on PTC invasion. It expanded the molecular mechanism of PTC invasiveness and provided experimental evidence for the prevention and treatment of PTC. We hope to further study the molecular mechanisms of miR-146b-5p regulation in MALAT1 in vivo through nude mouse tumor formation experiments, laying the foundation for the treatment of highly aggressive PTC.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by Hangzhou Major Science and Technology Innovation Projects (Grant No.: 20131813A08), Zhejiang Medical Science and Technology Program (Grant No.: 2018KY576), and Hangzhou Health and Medicine Science and Technology Plan Project (Grant No.: OO20191087).