
Abstract
Background:
Many patients with advanced cervical cancer (CC) have a poor prognosis and their mortality rank the first among women with malignant tumors. It's essential to explore the molecular mechanism of CC in clinical practice. Long noncoding RNA maternally expressed gene 3 (MEG3) has been reported to downregulate in CC tissues. However, the underlying mechanism of MEG3 in CC remains poorly elaborated. The current study aimed to explore the potential mechanism of MEG3 inducing endoplasmic reticulum stress (ERs)-mediated apoptosis of CC cells.
Methods:
The expression of MEG3 and miR-7-5p in CC tissues and cell lines was verified by quantitative reverse transcription/polymerase chain reaction (qRT-PCR). The vector of MEG3, miR-7-5p inhibitor, and sh-SCT1 were transfected into CC cell lines, and their expression was tested by qRT-PCR. Flow cytometry was used to detect apoptosis, and ERs-related protein expression was performed by Western blot. The regulatory relationship between MEG3/SCT1 and miR-7-5p was validated by Dual luciferase reporter assay.
Results:
CC tissues and cell lines showed downregulated MEG3 and STC1, and upregulated miR-7-5p. Overexpression of MEG3 or miR-7-5p inhibition induced ERs-triggered apoptosis of CC cells. In addition, sh-STC1 can reverse the effects of overexpressing MEG3 on CC cell apoptosis. In addition, dual luciferase reporter assay revealed that miR-7-5p can directly target to MEG3 and STC1.
Conclusion:
MEG3, act as a competing endogenous RNA of miR-7-5p, accelerates ERs-mediated apoptosis of CC cells through regulating SCT1 expression.
Introduction
Cervical cancer (CC) is a common gynecological malignancy and the generating causes of cancer induced death worldwide, and it is the main cancer of the female reproductive system. 1,2 The location of this disease is concealed and the early symptoms are not obvious, which makes the differential diagnosis difficult. 3 When the symptoms appear, the patient is approximately in advanced stage, which is difficult for treatment. 4 Studies demonstrated that a long-term exposure to high-risk human papillomavirus is considered to be one of the risk factors for CC. 5,6 At present, many patients with advanced disease still have poor tolerance, with no significant effect of chemotherapy, besides its easy recurrence and metastasis after surgery, resulting in a poor prognosis. 7 Therefore, clarifying the molecular mechanism of CC is particularly important in clinical practice.
Recently, studies have shown that long noncoding RNAs (lncRNAs) are genomic noncoding transcripts that regulate gene expression at the epigenetic, transcriptional, or translational level and playing an important role in disease progression. Accordingly, lncRNAs were used to be biomarkers in diagnosis and prognosis analysis in various cancers. 8 Maternally expressed gene 3 (MEG3) is a lncRNA and imprinted gene with tumor-inhibiting function, which is located on chromosome 14q32.3. 9 Previous reports have found that the downregulated expression of lncRNA MEG3 in CC was associated with poor prognosis. 10 LncRNA MEG3 can regulate the apoptosis and proliferation of CC cells by banding to miR-21. 11 However, the research of the function and the regulated molecular mechanism of MEG3 in CC is still less.
The endoplasmic reticulum (ER) is an organelle that undergoes protein synthesis, fold, modification, and regulates intracellular Ca2+ homeostasis. 12 Various stimuli can interfere with the homeostasis of the ER, resulting in the accumulation of unfolded or misfolded proteins and pathological outcomes, such as ER stress (ERs). 13 However, lncRNA MEG3 has not been reported on the regulation of ERs in CC cells, which is worthy of further investigation.
The dysregulated expression of MiR-7-5p was reported in various tumors, such as colorectal cancer, breast cancer, and glioblastoma. 14 –16 Anja Honegger et al. showed that silencing the E6/E7 oncogene reduced the expression of miR-7-5p in HeLa cells, which may play a procancer role in CC. 17 Besides, bioinformatics predictions suggested that lncRNA MEG3 can target miR-7-5p. However, the correlation between MEG3 and miR-7-5p in CC has not been reported, which deserves further study.
In this study, the authors confirmed that overexpression of MEG3 or miR-7-5p inhibition facilitated CC cell apoptosis. In addition, their results also demonstrated that lncRNA MEG3 mediates ER stress through the miR-7-5p/STC1 axis, which in turn causes apoptosis in CC cells. In addition, miR-7-5p can bind to MEG3 or STC1, directly. Taken together, our findings further revealed the molecular mechanism of the tumor suppressor MEG3 in CC, which provide a new theoretical basis for the CC treatment.
Methods
Patient samples and cell lines
In the study, 12 cases of CC tissue and adjacent normal tissue were collected in surgical patients with CC with the approval of the Ethics Committee of The Third Xiangya Hospital of Central South University. All tissue specimens were put in liquid nitrogen immediately after surgical resection. The patients could not receive any tumor-specific treatment before surgical resection and all patients received written informed consent.
The normal cervical epithelial cell line (H8) and four CC cell lines (HeLa, MS750, Caski, SiHa) were obtained from American Type Culture Collection (ATCC). All cells were cultured in RPMI 1640 medium (Gibco; CatNo: 21870-076), supplemented with 10% fetal bovine serum (FBS) (Gibco; CatNO: 16140071) and added penicillin (100 U/mL) and streptomycin (100 Ug/mL). All cells were cultured in a 5% CO2 incubator at 37°C.
Cell transfection
For MEG3 upregulation, full-length MEG3 gene sequence was cloned into the pcDNA3.1 vector (Invitrogen, Carlsbad, CA) and then transfected CC cells. An empty pcDNA3.1 vector was used as a negative control. For miR-7-5p inhibition or overexpression, cells were transfected with miR-7-5p inhibitor/mimic (Genechem, Shanghai, China). To interrupt STC1 expression, small hairpin RNA targeting STC1 was constructed by pRNAT-U6.1/Neo vector in CC cells. Lipofectamine 3000 reagent (Invitrogen) was used for the transient transfection into cells.
Quantitative reverse transcription/polymerase chain reaction
Total RNAs from cultured cells or CC samples was extracted using TRIzol reagent (Invitrogen) and synthesize the cDNA ( TaKaRa, Tokoyo, Japan). Quantitative reverse transcription/polymerase chain reaction (qRT-PCR) was performed on an ABI 7500 system (Applied Biosystems, Carlsbad, CA) using SYBR Green reagent (TOYOBO, Osaka Prefecture, Japan).
Specific primers for qPCR amplification were as follows: GAPDH-F: 5′-CCAGGTGGTCTCCTCTGA -3′; GAPDH-R: 5′- GCTGTAGCCAAATCGTTGT -3′; U6- F: 5′- CTCGCTTCGGCAGCACA -3′; U6-R: 5′- AACGCTTCACGAAT-TTGCGT-3′; MEG3-F: 5′-CCATCACCTGGATGCCTACG-3′; MEG3-R: 5′-GGGAATAGGTGCAGGGTGTC-3′; miR-7-5p-F: 5′-GGCGCTGGAAGACTAG-TGATTT-3′; miR-7-5p-R: 5′-GTGCAGGGTCCGAGGT-3′; STC1-F: 5′-CCTGAA-GCCATCACTGAGGT-3′; STC1-R: 5′-GAAGAGGCTGGCCATGTTAG-3′; In all experiments, the reference gene GAPDH or U6 were used to quantify the level of each sample using qRT-PCR. The relative gene expression for each sample was analyzed by the 2−ΔΔCt method.
Annexin V-FITC/PI double-stained assays
The Annexin V-FITC Apoptosis Detection Kit (BD Bioscience, Becton Dickinson Co.) was used to detect the apoptosis of cells. The cells were treated with trypsin without EDTA, collected by centrifugation, washed with prechilled PBS, and adjusted to 1–5 × 105 cells. One hundred microliters of 1 × Binding Buffer was resuspended. The sample also required the addition of 5 μL of Annexin V-FITC and 10 μL of PI Staining Solution and gently mixed. The reaction was allowed at room temperature for 10–15 min away from light. Four hundred microliters of 1 × Binding Buffer was added to the above reaction sample, mixed, and placed on ice. The sample was detected by flow cytometry by using a FACSCalibur flow cytometer (BD Biosciences, San Jose, CA) within 1 h.
Measurement of mitochondrial membrane potential detection
Mitochondrial membrane potential (MMP) was measured by flow cytometry using the MMP Detection Kit (JC-1) (Sigma; CS0390). Mitochondrial electrochemical potential gradient (Δψ) depletion is considered a symptom of ERs. The kit uses a cationic lipophilic dye JC-1 to conveniently and rapidly detect changes in the electrochemical potential of mitochondrial inner membranes in living cells. Thirty micromole JC-1 reagent was added to the treated cells, and cultured in the dark at 5% CO2 in a humid atmosphere for 20 min at 37°C. The cells were analyzed in 30 min by FACSCalibur flow cytometer (BD Biosciences).
Intracellular calcium ion concentration detection
The cells were seeded onto slides in a six-well plate, and the cell density was 50%–80% overnight. A fluorescent calcium indicator of Fluo-4 AM at a final concentration of 2 μM was added, and incubation was conducted at 37°C for 60 min for fluorescent probe loading, and then washed with PBS. The intracellular calcium ion was measured by FACSCalibur flow cytometry (BD Biosciences).
Western blot assays
The expression of apoptosis-related proteins and ERs-related proteins in CC cells was measured by Western blotting. Separation of proteins from cells was by dodecyl sulfate sodium salt/polyacrylamide gel electrophoresis, followed by standard techniques for immunoassays.
Antibodies referred to Bax (Abcam, # ab32503, 1:2000 dilution), Bcl-2 (Abcam; # ab185002, 1:2000 dilution), cleaved caspase-3 (Abcam; # ab2302, 1:2000 dilution), protein kinase-like ER kinase (PERK, CST; #5683, 1:2000 dilution), Phospho-PERK (CST; #3179, 1:2000 dilution), Cleaved activating transcription factor 6 (ATF6, Abcam; #ab62576, 1: 2000 dilution), glucose-regulated protein (GRP78, Abcam; #ab21685, 1: 2000 dilution), inositol-requiring enzyme 1 (IRE1, CST; #3294, 1:2000 dilution), Phospho-IRE1 (Abcam; #ab48187, 1: 2000 dilution), HRP-conjugated secondary antibodies (Invitrogen; G-21040). Controls were referred to β-actin (CST; #4970, 1:5000 dilution). ImageJ software (NIH, Bethesda, MD) was used to analyze the expression of each protein and recorded as integral density (ID).
Dual luciferase reporter assay
The wild-type sequence of MEG3 (WT-MEG3) or 3′UTR of STC1 (WT-STC1) and their corresponding mutant-type (MUT-MEG3 or MUT-STC1) sequence were cloned into pGL4 luciferase reporter plasmids (Promega, Madison, WI). In addition, the reporter plasmids (WT-MEG3, WT-STC1, MUT-MEG3, or MUT-STC1) were cotransfected into HeLa cells with miR-7-5p mimics. After transfection of 48 h, the luciferase activity was detected using the Dual Luciferase Reporter Assay Kit (Promega Corporation).
Statistical analysis
All data are expressed as the mean ± Standard Deviation (SD). Each assay was performed in triplicate. The differences between two groups were analyzed by Student's t-test. Multiple sets of data were compared using one-way analysis of variance (GraphPad version 7.0, GraphPad Software, CA). P < 0.05 indicates that the difference is statistically significant.
Results
MEG3/miR-7-5p/STC1 were dysregulated in CC tissues and cancer cells
MEG3 expression (Fig. 1A) and STC1 mRNA expression (Fig. 1C) were notably reduced in tumor tissues than that in adjacent tissues, whereas miR-7-5p expression level was remarkably increased in tumor tissues than that in adjacent tissues (Fig. 1B). Next, they assessed the expression level of MEG3, miR-7-5p, and STC1 mRNA in normal cervical epithelial cell line (H8) and CC cell lines (HeLa, MS750, Caski, SiHa). The expression levels of MEG3 and STC1 mRNA in CC cell lines were significantly decreased, however, miR-7-5p expression was crucially increased compared with H8 cells, especially in HeLa cells (Fig. 1D–F). According to the detection results, HeLa with relatively low expression of MEG3 and Caski cells with relatively high MEG3 expression were selected for subsequent experiments.

MEG3/miR-7-5p/STC1 were dysregulated in CC tissues and cancer cells
Overexpression of MEG3 promotes the apoptosis of CC cells
We transfected HeLa and CaSki cells with the overexpression MEG3 plasmid (pcDNA3.1-MEG3) and empty pcDNA3.1 vector (vector), control represents that the cells were not transfected. The results of qRT-PCR showed that the vector group showed no significant changes in MEG3 and miR-7-5p expression than the control group in the HeLa and CaSki cell lines. The MEG3 levels was expressively increased in the overexpressed MEG3 group comparison with the vector or control group (Fig. 2A). However, the expression of miR-7-5p was significantly reduced in the overexpressed MEG3 group compared with the vector or control group (Fig. 2B), and the result indicated that there was a certain relationship between MEG3 and miR-7-5p, which needed to be further verified.

Overexpression of MEG3 promotes the apoptosis of CC cells
To explore whether overexpressed MEG3 affected CC cell apoptosis, flow cytometry was selected for detection. After 48 h of transfection, the number of apoptosis in overexpressed MEG3 group was notably increased compared with the vector group, indicating that MEG3 overexpression promoted cell apoptosis (Fig. 2C, D). To further validate the effect of MEG3 on CC cells, Western blot was used to detect the CC cell apoptosis-related proteins. Our results revealed that the protein level of cleaved caspase-3 expression remarkably increased, and the Bcl-2/Bax expression ratio was reduced in the overexpressed MEG3 group than that in vector group (Fig. 2E, F).
Overexpression of MEG3 regulates ERs response
The overexpressed MEG3 (pcDNA3.1-MEG3) and empty pcDNA3.1 vector (vector) were transfected into HeLa and CaSki cells. Compared with the vector and control groups, the Ca2+ concentration in the overexpressed MEG3 group was significantly increased (Fig. 3A, B). MMP measurements showed a significant loss of mitochondrial activity in the overexpressed MEG3 group compared with the vector or control groups (Fig. 3C, D). The ERs-related protein (GRP78, PERK, IRE1, ATF6) expression was confirmed (Fig. 3E, F). Our results proved that the protein levels of GRP78, cleaved ATF6, and the ratio of p-PERK/PERK and p-IRE1/IRE1 were prominently promoted in the overexpressed MEG3 group compared with the vector or control groups of CC cells. The results indicated that MEG3 overexpression could on occasion cause ERs of CC cells.

Overexpression of MEG3 regulates ERs response
Inhibition of MiR-7-5p triggers ERs-induced apoptosis of CC cells
We transfected HeLa and CaSki cells with the miR-7-5p inhibitor or inhibitor NC; control represents that the cell was not transfected. The expression of miR-7-5p and STC1 were detected by qRT-PCR. Inhibitor NC group showed no significant changes in miR-7-5p and STC1 expression compared with the control group in the HeLa and CaSki cell lines. The expression of miR-7-5p was observably reduced in the miR-7-5p inhibitor group than the inhibitor NC group (Fig. 4A). However, the STC1 mRNA levels were expressively increased in the miR-7-5p inhibitor group than inhibitor NC group (Fig. 4B). MiR-7-5p with low expression could increase the expression of STC1, indicating that there was a certain relationship between the two, which needed further verification.
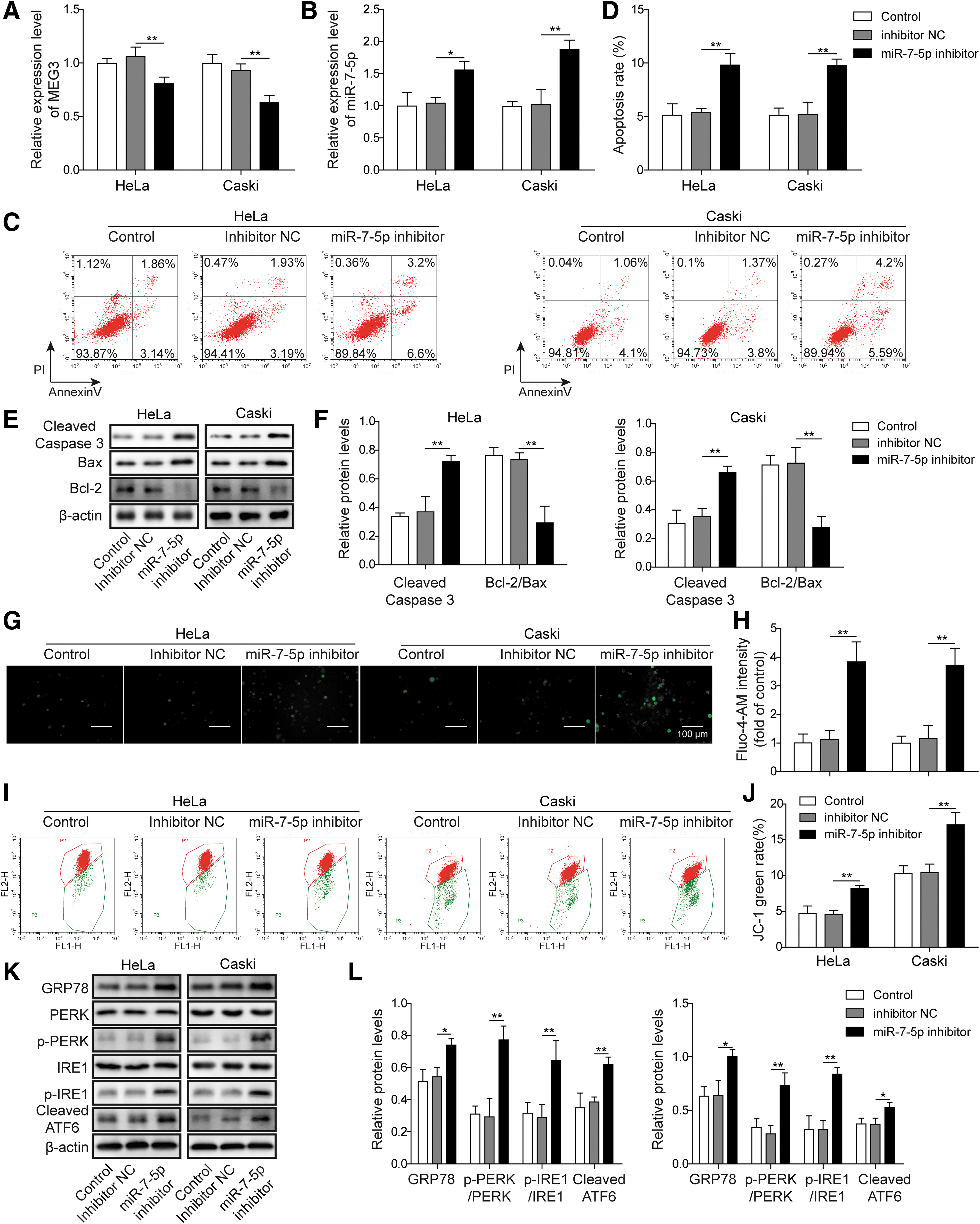
Inhibition of miR-7-5p regulates STC1 expression and triggers apoptosis of CC cells
To explore whether miR-7-5p inhibitor affected CC cell apoptosis, they used flow cytometry for detection. After 48 h of transfection, the level of apoptosis in miR-7-5p inhibitor group was notably increased than the inhibitor NC or control group (Fig. 4C, D), pointing that the decreased expression of miR-7-5p could promote cell apoptosis. Western blot was used to detect the expression of Bcl-2, Bax, and cleaved caspase-3. The results from this study revealed that the protein level of cleaved caspase-3 expression remarkably increased, and the Bcl-2/Bax expression ratio was reduced in miR-7-5p inhibitor group than that of the inhibitor NC or control group (Fig. 4E, F).
The intracellular calcium concentration was detected by Fluo-4-AM fluorescent labeling. Compared with the inhibitor NC or control groups, the Ca2+ concentration in the miR-7-5p inhibitor group was obviously promoted (Fig. 4G, H). Besides, the MMP was significantly decreased in the miR-7-5p inhibitor group than the inhibitor NC or control groups (Fig. 4I, J). In addition, the ERs-related protein (GRP78, PERK, IRE1, and ATF6) expression was confirmed by Western blot. In the HeLa and CaSki cells, the protein levels of GRP78, cleaved ATF6, and the ratio of p-PERK/PERK and p-IRE1/IRE1 were lower in the inhibitor NC and control groups than the miR-7-5p inhibitor group (Fig. 4K, L).
Knockdown of STC1 reverses the effect of MEG3 on CC cells
The qRT-PCR analysis was used to analyze the expression of STC1 mRNA in CC cells. Figure 5A showed that the STC1 mRNA levels were expressively increased in the overexpressed MEG3 group compared with the vector or control group. However, the expression of STC1 was significantly reduced in the sh-STC1 group than sh-NC group (Fig. 5B). To explore the network between MEG and STC1 and their effect on CC cell apoptosis, they used flow cytometry for detection. The sh-STC1 and pcDNA3.1-MEG3 were cotransfected into HeLa and CaSki cells. After 48 h of transfection, the cell apoptosis in sh-STC1+MEG3 group was notably inhibited. The result indicated that knockdown of STC1 expression could reverse the proapoptotic effect of MEG3 overexpression on CC cells (Fig. 5C, D).
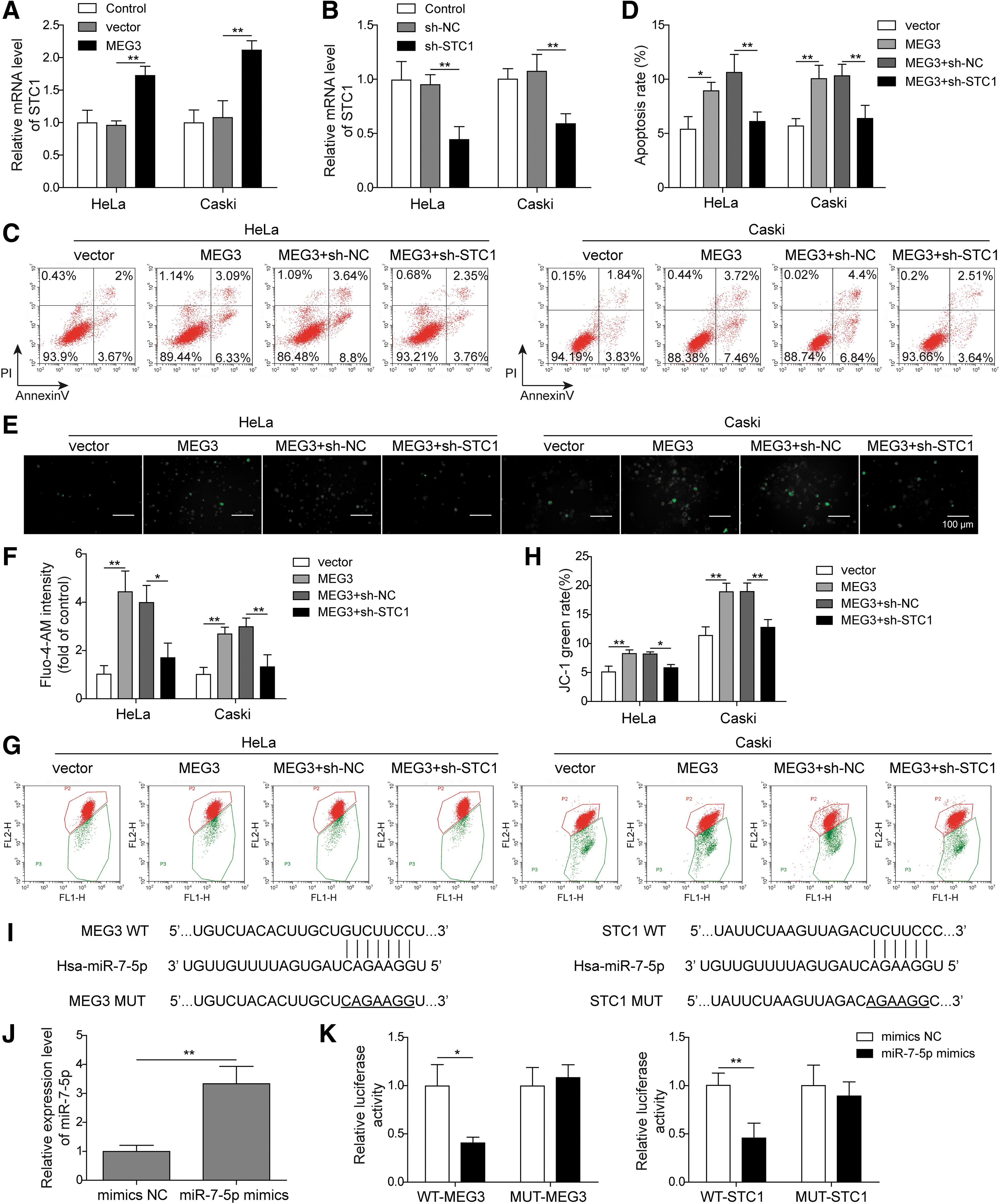
Knockdown of STC1 reverses the effect of MEG3 on CC cells
The intracellular calcium concentration detection results showed that the Ca2+ concentration in the sh-STC1 and pcDNA3.1-MEG3 cotransfection group was significantly reduced. Knockdown of STC1 reversed the increase in intracellular calcium that was caused by overexpression of MEG3 (Fig. 5E, F). MMP measurements showed a significant increase in sh-STC1 and pcDNA3.1-MEG3 cotransfection groups. Knockdown of STC1 expression could reverse the decrease in MMP that was caused by MEG3 overexpression (Fig. 5G, H).
The prediction database shows that miR-7-5p is the target of MEG3 and STC1, and their binding sites are shown in Figure 5I. Additionally, luciferase reporter assay was used to validate the relationship between miR-7-5p and MEG3 or STC1. MiR-7-5p mimic was transfected into HeLa cells, and the expression level of miR-7-5p was detected (Fig. 5J). In the WT-MEG3 and WT-STC1 groups, the luciferase activity was decreased after transfection with miR-7-5p mimic than mimic NC group. However, the luciferase activities were unchanged in the MUT-STC1 and MUT-MEG3 groups after transfection (Fig. 5K). This result illustrated that miR-7-5p could directly target MEG3 and STC1.
These results show that MEG3 is the upstream target gene of miR-7-5p, which regulates the expression of miR-7-5p, and that STC1 is a downstream target of miR-7-5p. MEG3 binds to miR-7-5p, resulting in decreased binding of free miR-7-5p to target STC1 mRNA, thereby upregulating STC1 expression and mediating ERs-induced apoptosis in CC cells.
Discussion
Although numerous studies have explored the pathogenesis of CC and greatly improved the diagnosis and treatment strategy of CC, many patients have poor prognosis due to unclear early diagnosis, which affects the survival of patients. 18,19 Tumor development is a complex process that involves the activation of proto-oncogenes and the mutation of antioncogenes. During the disease development, abnormal transcription and gene mutation play a vital role, and the function changes of cells can eventually lead to CC. 20 Therefore, it is particularly important to study the pathogenesis of CC.
It is well known that lncRNA can participate in the regulation of gene expression, dose compensation, genomic imprinting, transcriptional activation, transcriptional interference and nuclear transport, X chromosome silencing, and chromatin modification in a variety of important biological processes. 21,22 In recent years, lncRNA has become an increasingly important component of cancer transcriptome, and it plays a pivotal role in the occurrence and development of various cancer cells through the interaction with protein, DNA and RNA. 23 The study has showed the clinical value of lncRNA MEG3 in patients with CC, and a series of molecular experiments identified epigenetic regulation of lncRNA MEG3 in CC tissues and cells. 24,25
The results from this study indicated that MEG3 was underexpressed in CC tissues and cells, and overexpression of MEG3 promoted ERs-induced CC cell apoptosis.
MiRNAs can bind with mRNAs, causing gene silencing and thus affecting cell fates. The previous study found miR-7-5p dysregulation in CC. miR-7-5p promotes autophagy by targeting BCL2 and PARP-1, reducing DNA repair activity and maintaining energy homeostasis in cisplatin-resistant CC cells. 26 In this study, the results demonstrated that inhibition of miR-7-5p can accelerate the ERs-related apoptosis of CC cells.
STC is a glycoprotein hormone that plays an important role in the homeostasis of calcium and phosphate. 27 Its human homologous genes, STC1 (247 amino acids) and STC2 (302 amino acids) expressed in various tissues have been used in endocrinology and cardiovascular diseases' treatment. 28,29 Studies have demonstrated that STC1 plays a key role in tumor and disease progression, such as breast cancer, thyroid cancer, and gastric cancer, and is associated with disease progression. 30 –32 Studies have confirmed that STC1 has the effect of correcting the inappropriate relationship between epithelial/mesenchymal cells. Mesenchymal stem cells can inhibit pulmonary fibrosis by secreting STC1 to regulate ER stress. 33,34
The findings from this study proved that MEG3 overexpression reduced the level of STC1 in CC cells, and after cotransfection with pcDNA3.1-MEG3 and sh-STC1, reverses the effect of MEG3 on ERs-induced proapoptosis in CC cells.
Competing endogenous RNAs (ceRNAs) hypothesis reveals a new interaction mechanism between RNAs. Act as a ceRNAs, lncRNA, or circRNAs regulate gene expression by competitively binding to miRNAs. 35,36 For example, lncRNA-ATB combining with miR-200 promotes ZEB1/ZEB2 expression to induce the invasion/metastasis cascade of hepatocellular carcinoma. 37 MEG3 inhibits vascular endothelial cell senescence by regulating miR-128-dependent Girdin expression. 38 The relationship between MEG3 and miRNA-7 has been demonstrated in clear cell renal cell carcinoma (CCRCC). MEG3 competitively binds to miR-7 and inhibits cell proliferation, invasion, and migration in CCRCC by regulating RASL11B expression. 39 However, the relationship between MEG3 and miR-7 in CC remains unclear.
In this study, miR-7-5p proved the target of MEG3 and STC1. The results depicted a potential that MEG3 upregulated the level of STC1 by binding miR-7-5p, thereby mediated ERs-triggered apoptosis of CC cells.
Conclusion
In conclusion, the results from this study demonstrated that upregulation of MEG3 triggers apoptosis of CC cells associated with ERs. MEG3 may act as a ceRNA of miR-7-5p and affect the expression of STC1, in turn. The findings further revealed the molecular mechanism of the tumor suppressor MEG3 in CC, which provide a new theoretical basis for the CC treatment.
Footnotes
Disclosure Statement
The authors declare no competing interests.
Funding Information
This work was financially funded by the Natural Science Foundation Youth Fund Project of China Hunan Province (2019JJ50907).