
Abstract
Background:
Long noncoding RNA (lncRNA)-PRLB (progression-associated lncRNA in breast cancer) has been identified to enhance the drug resistance of breast cancer cells. In this study, the authors explored PRLB effect in the paclitaxel (Tax) resistance of ovarian cancer cells and revealed the role of RSF1 (remodeling and spacing factor 1)/nuclear factor kappaB (NF-κB) signaling in this process.
Materials and Methods:
Tax resistance was established in CAOV3 and SKOV3 cell lines. The expressions of PRLB in Tax resistant tissues and cells of ovarian cancer were detected using the real time polymerase chain reaction assay. MTT [3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide] and flow cytometry were used to detect cell survival and apoptosis. The RNA-binding protein immunoprecipitation (RIP) assay and/or the luciferase gene reporter assay were used to assess the cross talk among miR-150-5p and PRLB/RSF1.
Results:
PRLB expression was obviously enhanced in the Tax resistant ovarian cancer tissues and cells. Depletion of PRLB induced a significant decrease in the IC50 value of the CAOV3/Tax and SKOV3/Tax cells and increased cell apoptosis, as well as increased miR-150-5p expression through a direct binding. In addition, miR-150-5p upregulation decreased the luciferase activity of PRLB and RSF1, whereas this effect was abolished when the putative binding sites were mutated. And overexpression of RSF1 significantly rescued the effect of PRLB downregulation-caused decrease in the IC50 value and the increase in cell apoptosis and the decreased expressions of RSF1 and p-p65.
Conclusion:
This study reveals that knockdown of PRLB improves the sensitivity of ovarian cancer cells to Tax, at least in part, through inhibiting the activation of RSF1/NF-κB signaling through targeting miR-150-5p.
Introduction
Ovarian cancer ranks the seventh in all kinds of cancers and the leading cause of death in women in the world. It's estimated that there are roughly 295, 414 new cases and 184, 799 mortalities every year. 1 Most patients are at advanced stages when they are first diagnosed due to the lag in the effective screening methods, 2 for whom the 5-year overall survival rate was only 29% in United States. 3 The combination of surgery resection and chemotherapy is needful for this kind of ovarian cancer. Although the first-line chemotherapy with platinum drugs and paclitaxel (Tax) has a response rate of >80%, 70% patients will have a recurrence within 2–3 years due to the resistance to chemotherapy. 4 Therefore, elucidation of the mechanism underlying chemotherapy resistance in ovarian cancer is needed.
Long noncoding RNAs (lncRNAs) are a class of noncoding RNAs (ncRNAs), which have ≥200 nucleotides in length. 5 Although lacking of cross-species conservation, 6 lncRNAs have been demonstrated to play vital roles in multiple biological processes, such as cell proliferation, survival, differentiation, and cell cycle; and their deregulation facilitates the initiation, development, and chemoresistance of many kinds of malignant cancers, including ovarian cancer. 7 –9 For example, lncRNA GAS5 (growth arrest-specific transcript 5) was reported to be expressed at a low level in ovarian cancer samples and correlated with poor prognosis; in addition, GAS5 was downregulated in ovarian cancer cells with cisplatin (DDP) resistance, and upregulation of GAS5 dramatically increased the sensitivity of ovarian cancer cells to DDP both in vivo and in vitro. 10 The lncRNA-PRLB (progression-associated lncRNA in breast cancer) was demonstrated to play a pro-tumor role in breast cancer and significantly reinforced cell resistance to 5-fluorouracil. 11 However, whether PRLB is implicated in the chemotherapy resistance of ovarian cancer still remains unknown.
Using the bioinformatics methods, miR-150-5p was identified as a predicted target of PRLB and has been reported to play a role in the drug resistance of cancers. 12 For instance, Palagani et al. 12 reported that combination of miR-150-5p with low doses of glucocorticoids in multiple myeloma cell line MM1S was found to sensitize therapy response, whereas opposite effects could be observed with a miR-150-5p downregulation. In addition, Wuerkenbieke et al. 13 found that miR-150 downregulation enhanced the resistance of ovarian cancer cells to pertuzumab. Similarly, Kim et al. 14 reported that miR-150 enhanced the paclitaxel sensitivity in paclitaxel-resistant ovarian cancer cells. However, whether PRLB modulates chemotherapy resistance of ovarian cancer by targeting miR-150-5p still remains unclear.
RSF1, also known as a histone-binding protein, is of importance in transcriptional regulation and chromatin remould. 15 Noticeably, it is well documented that RSF1 contributes to the chemoresistance of many types of cancers through activating the nuclear factor kappa B (NF-κB) pathway, such as nonsmall cell lung cancer (NSCLC), 16 nasopharyngeal carcinoma, 17 glioma, 18 and ovarian cancer, 19 suggesting that RSF1/NF-κB signaling might be a potent target to prevent the development of drug resistance. The bioinformatics predicted that RSF1 was a direct target of miR-150-5p; hence, the authors speculated that PRLB could modulate the chemotherapy resistance of ovarian cancer by targeting miR-150-5p/RSF1.
In this study, the authors explored whether PRLB implicates in the Tax resistance of ovarian cancer cells by modulating miR-150-5p/RSF1/NF-κB signaling. Their results showed that PRLB1 displayed an amplified expression profile in ovarian cancer cells, which were resistant to Tax; and overexpression of PRLB significantly enhanced the resistance of ovarian cancer SKOV3 and CAOV3 cells to Tax, at least in part, through activating the RSF1/NF-κB signaling by targeting miR-150-5p.
Materials and Methods
Tissue samples
Forty-five ovarian cancer tissue samples were collected from 45 patients with epithelial ovarian cancer (EOC), among which 25 tissues were from treatment responsive patients and 20 tissues were from treatment resistance patients. The information of the patients is demonstrated in Supplementary Table S1. All patients signed the informed consent. The treatment-resistant tumors were those persistent or recurrent diseases within 6 months after the initiation of first-line combination chemotherapy of Tax and platinum. Treatment-responsive tumors were those with a complete response to chemotherapy and a more than 6 months platinum-free interval. 7 The experimental protocols involving human samples were approved by the Ethics Committee of Renmin Hospital of Wuhan University.
Cell culture and culture
Two human ovarian cancer cell lines SKOV3 and CAOV3 were acquired from American Type Culture Collection (Manassas, VA). The cells were cultured in RPMI-1640 medium, including 10% fetal bovine serum at 37°C with 5% CO2. The products used in this part were all obtained from Thermo Fisher Scientific (Waltham, MA).
Establishment of the Tax resistant cell lines
The Tax-resistant ovarian cancer cell lines SKOV3/Tax and CAOV3/Tax were generated according to a previous study. 20 In brief, the parental CAOV3 and SKOV3 cells were exposed to different concentrations of Tax (from 1 to 30 nM). The Tax concentration was increased by 2 nM until it reached 10 nM every other week. Following 2 months of incubation with 10 nM Tax, Tax concentration was increased by 5 nM every month until it reached 30 nM. The SKOV3/Tax and CAOV3/Tax cell lines were maintained in 30 nM Tax to keep the drug-resistant phenotype. The above obtained drug resistant cell lines were maintained in Tax-free medium for at least 2 weeks before experiment.
Western blotting
The total protein extraction was carried out using the Lysis Buffer containing protease inhibitor (Beyotime Biotechnology, Beijing, China), followed by concentration measurement with the BCA Protein Assay Kit (Thermo Fisher Scientific). Then the proteins were selected by polyacrylamide gel and then transferred to the polyvinylidene fluoride membrane (Millipore, Billerica, MA), followed by being blocked with 5% fat-free milk diluted in Tris-HCl buffer salt solution-Tween 20 (TBST) for 1 h and incubated with the indicated first antibodies and secondary antibodies. The protein contents were determined using the ImageJ software with normalization to β-actin. The first antibodies used in this experiment included anti-RSF1 antibody (No. ab109002; Abcam, Cambridge, MA), anti-p-p65 antibody (No. 3033; Cell Signaling Technology, MA) and anti-p65 antibody (No. 8242; Cell Signaling Technology).
Cell transfection
The short hairpin RNAs (shRNAs) used to silence PRLB in CAOV3/Tax and SKOV3/Tax cells, and the negative control vector (shRNA-NC), together with the mimic and inhibitor used to overexpress (miR-150-5p, miR-NC) and downregulate miR-150-p (inh-miR-150-5p, inh-NC) were obtained from GenePharma (Shanghai, China). The plasmid used to upregulate RSF1 and the Control vector were all purchased from the OriGene (Beijing, China). The shRNAs were infected into cells with the help of polybrene, and other vectors were introduced into cells using the Lipofectamine 2000 transfected reagent (Invitrogen, Carlsbad, CA) referring to the manufacturer's instruction.
Real time polymerase chain reaction
The total mRNA was extracted from cells using the TRIzol reagent (Invitrogen) and was then reversely transcribed into the complementary DNA (cDNA) followed by detection using the RT-PCR Kit (Thermo Fisher Scientific) for mRNA detection and TaqMan microRNA (miRNA) assays (Applied Biosystems, Foster City, CA) for miRNA based on the following conditions: 95°C for 5 min, 95°C for 15 s, and 60°C for 1 min for a total of 40 cycles. GAPDH and U6 were used as an internal control for the quantification of mRNA and miRNA level.
PRLB-sense: 5′-ACGCCATGTTGGGAGACTTC-3′, antisense: 5′-TGAAAGCCCAGGGTCAACTC-3′; GAPDH-sense: 5′-GGAGTCAACGGATTTGGTCG-3′, antisense: 5′-TTCCCGTTCTCAGCCATGTAG-3′.
Cell survival detection
Ovarian cancer cells were plated into 96-well plates with 5000 cells in each well. Post 24 h of transfection, the cells were treated with different concentrations of Tax. Post 48 h of transfection, MTT [3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide] solution (5 mg/mL; Beyotime) was added into each well with a dosage of 20 μL, and the cells were incubated at 37°C for 4 h. Then, 150 μL dimethyl sulfoxide (DMSO) (Beyotime) was added into each well for 10 min. The absorbance at 490 nm was measured with a microplate reader. The median inhibitory concentration (IC50) for Tax in ovarian cancer was evaluated using the relative survival curves.
Cell apoptosis detection
Cell apoptotic rates were detected using the flow cytometry with the help of an Annexin V-FITC/propidium iodide (PI) Cell Apoptosis Detection Kit (BD Biosciences, CA). In brief, about 5 × 105 ovarian cancer cells were seeded into the six plates and submitted to different cell transfections, followed by treatment with 100 nM Tax. After 48 h, the cells were harvested and washed with phosphate buffered saline once. Then, the cells were resuspended with 500 μL of 1 × binding buffer consisting of 5 μL Annexin V-FITC and maintained for 15 min in the dark. Next, the cells were incubated with 10 μL PI solution and incubated for another 5 min. The cell apoptosis rate, including both early and late, was measured on a flow cytometer (Beckman Coulter, Inc.).
Luciferase reporter assay
Thewild type (WT) and mutant type (MUT) of PRLB/RSF1 sequence, including the putative miR-150-5p binding site, were inserted into the pmirGLO vector (Promega, Madison, WI). Then, the SKOV3/Tax and CAOV3/Tax cells were cotransfected with the recombinant WT/MUT plasmids and miR-150-5p or miR-NC. Post 48 h of cell transfection, the Renilla luciferase activity was measured using the Dual-Luciferase Reporter Assay System (Promega), with firefly luciferase as a control.
RNA-binding protein immunoprecipitation assay
The RNA-binding protein immunoprecipitation (RIP) assay was executed using the Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore) based on the descriptions. The coprecipitated RNAs were measured by the real time polymerase chain reaction (RT-PCR) technology with total RNA (input controls) and corresponding species IgG controls being detected at the same time.
Statistical analysis
Statistical analysis was executed using the SPSS 21.0 software. The results were present with the mean ± standard deviation and were analyzed with Student's t test or one way analysis of variance (ANOVA). If the p-value was less than 0.05, the differences between groups were identified as significant.
Results
The expression of PRLB is upregulated in the Tax-resistant ovarian cancer tissues and cell lines
To explore the effect of PRLB in the drug resistance of ovarian cancer, the authors first detected the expression patterns of PRLB in the Tax-resistant ovarian cancer tissues and cell lines. Compared with the treatment-responsive ovarian cancer tissues (n = 25), the expression of PRLB was dramatically increased in the treatment-resistance tissues (n = 20) (Fig. 1A). The authors then established the Tax-resistance cell lines in ovarian cancer CAOV3 and SKOV3 cells. The IC50 value of CAOV3 cells to Tax was 4.064 nM, and it was significantly increased in the CAOV3/Tax cells with an IC50 value of 1566 nM (Fig. 1B). Consistently, the IC50 value of SKOV3 cells to Tax was 8.695 nM, and it was significantly increased in SKOV3/Tax cells with an IC50 value of 3135 nM (Fig. 1B). In addition, the expression level of PRLB was obviously elevated in the SKOV3 and CAOV3 cell lines compared with their parental cell lines (Fig. 1C). These results demonstrated that PRLB was overexpressed in the drug-resistant tissues and cell lines of ovarian cancer.
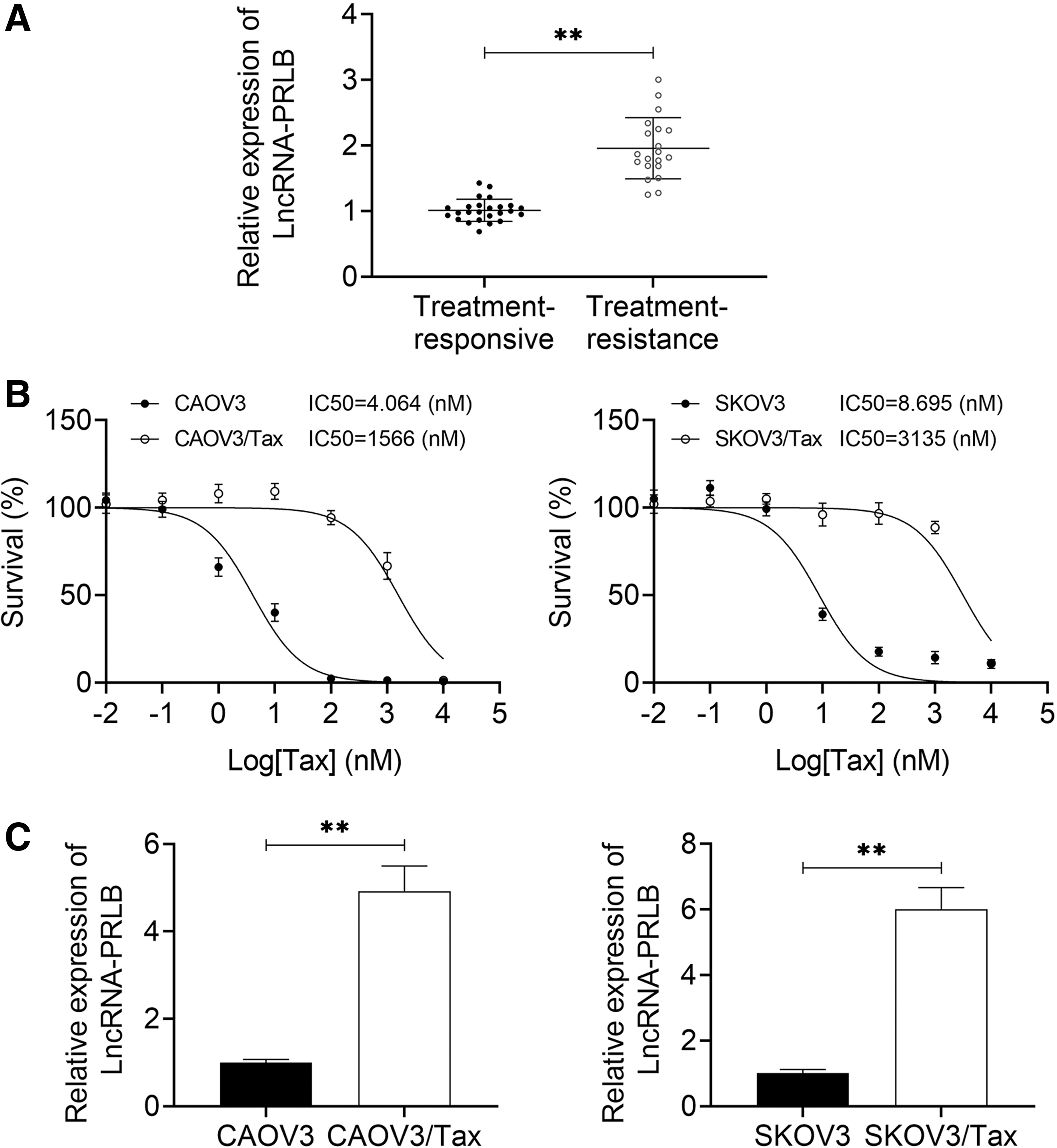
PRLB was overexpressed in the Tax-resistant ovarian cancer tissues and cell lines.
Downregulation of PRLB reverses the Tax resistance in SKOV3/Tax and CAOV3/Tax cells
To further explore the role that PRLB plays in the Tax resistance in ovarian cancer cells, the authors then downregulated PRLB expression in CAOV3/Tax and SKOV3/Tax cells. The expression of PRLB was significantly reduced when CAOV3/Tax and SKOV3/Tax cells were infected with the shRNA-1/2 targeting PRLB, with shRNA-1 showing the best knockdown efficiency (Fig. 2A), which was chosen for the following experiments. Downregulation of PRLB reduced the IC50 value of SKOV3/Tax and CAOV3/Tax cells (Fig. 2B). In addition, knockdown of PRLB induced a significant increase in cell apoptosis rate in CAOV3/Tax and SKOV3/Tax cell lines (Fig. 2C). These results indicated that downregulation of PRLB improved the sensitivity of Tax-resistant ovarian cancer cells to Tax.
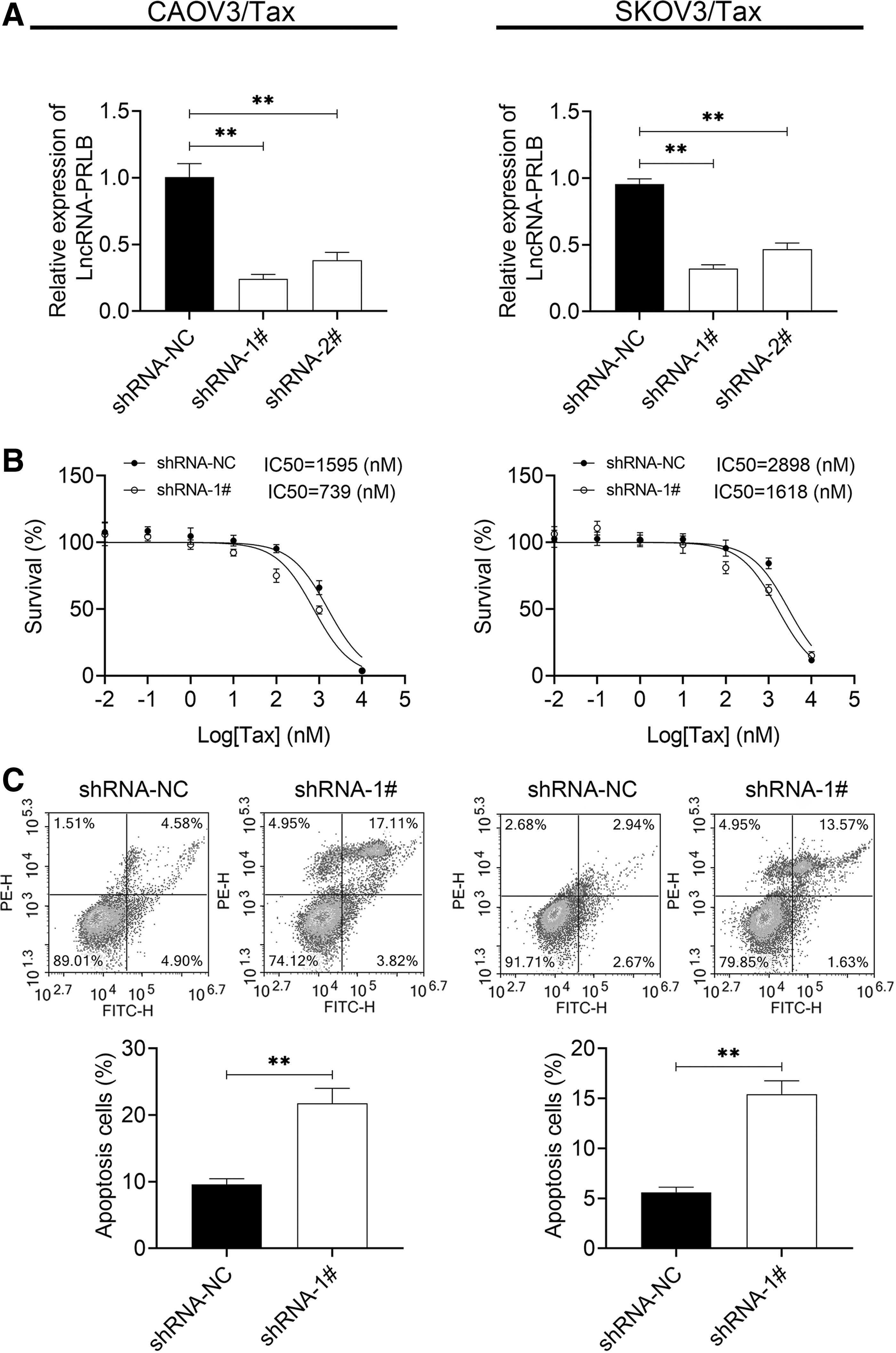
Knockdown of PRLB enhanced the sensitivity of CAOV3/Tax and SKOV3/Tax cells to Tax.
PRLB targets miR-150-5p in CAOV3/Tax and SKOV3/Tax cells
To reveal the underlying mechanism of PRLB in the drug resistance of ovarian cancer cells to Tax, the authors then applied the bioinformatics and found that miR-150-5p was a predicted target of PRLB. The putative binding sites were displayed (Fig. 3A). To further explore the relationship between miR-150-5p and PRLB, the authors then carried out the luciferase reporter and RIP assays. The expression of miR-150-5p was apparently increased following CAOV3/Tax and SKOV3/Tax cell transfection with miR-150-5p compared with miR-NC transfection (Fig. 3B). The luciferase activity was repressed after cells were overexpressed in miR-150-5p compared with the miR-NC group with PRLB-WT plasmid transfection, but it was abolished when the cells were transfected with the PRLB-MUT plasmid in both CAOV3/Tax and SKOV3/Tax cell lines (Fig. 3C). In addition, the RIP result confirmed that there was a combination of PRLB and miR-150-5p in CAOV3/Tax and SKOV3/Tax cell lines (Fig. 3D). And downregulation of PRLB significantly increased the expression of miR-150-5p (Fig. 3E). These findings suggested that miR-150-5p was a direct target of PRLB in ovarian cancer.
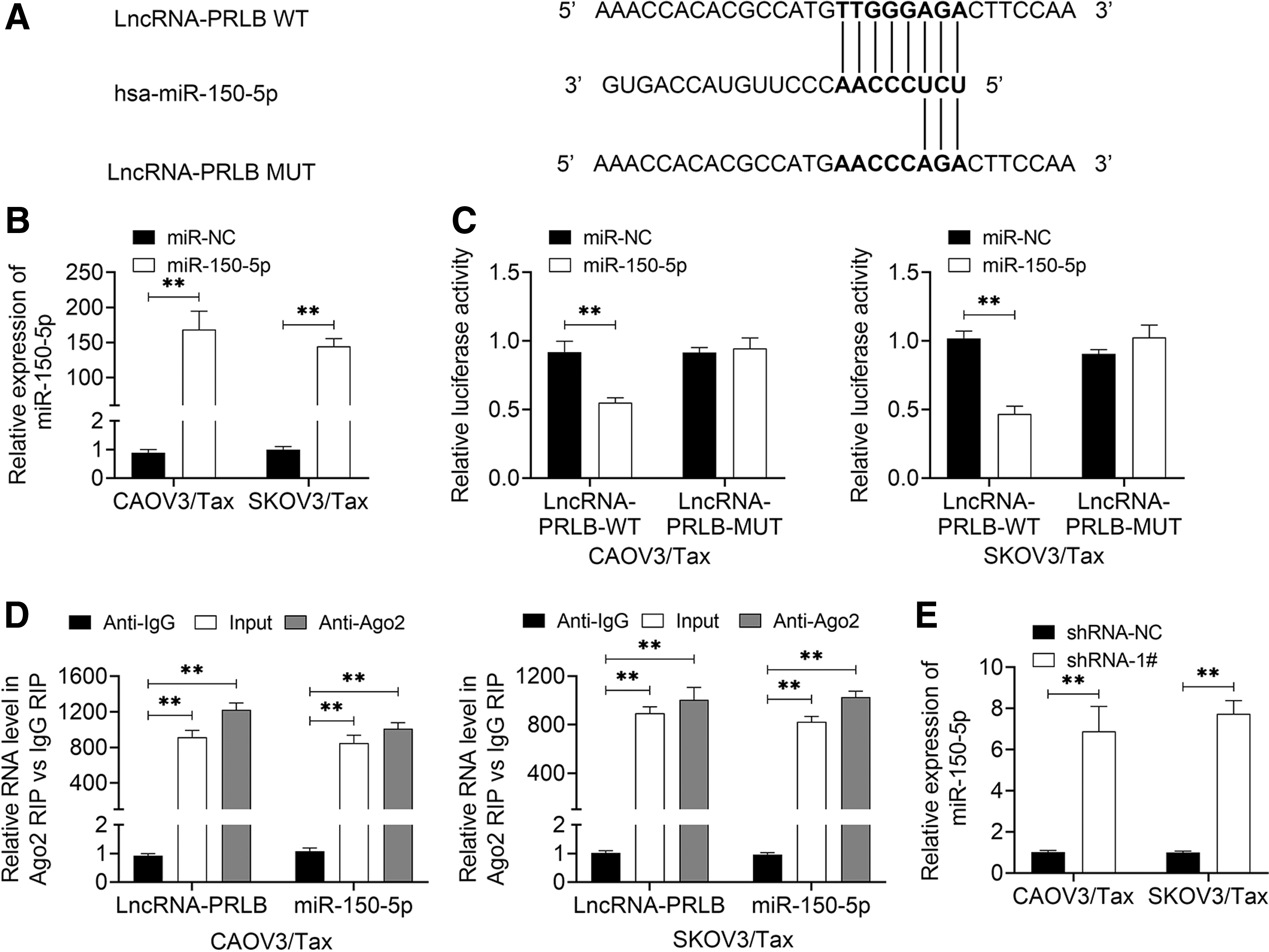
miR-150-5p was a direct target of PRLB in ovarian cancer.
Downregulation of PRLB downregulates RSF1 expression by targeting miR-150-5p
Then, the authors explored the relationship between RSF1 and PRLB/miR-150-5p. Compared with the parent SKOV3 and CAOV3 cell lines, RSF1 expression levels were significantly increased in both CAOV3/Tax and SKOV3/Tax cell lines at mRNA and protein levels (Fig. 4A, B). The bioinformatics software TargetScan (
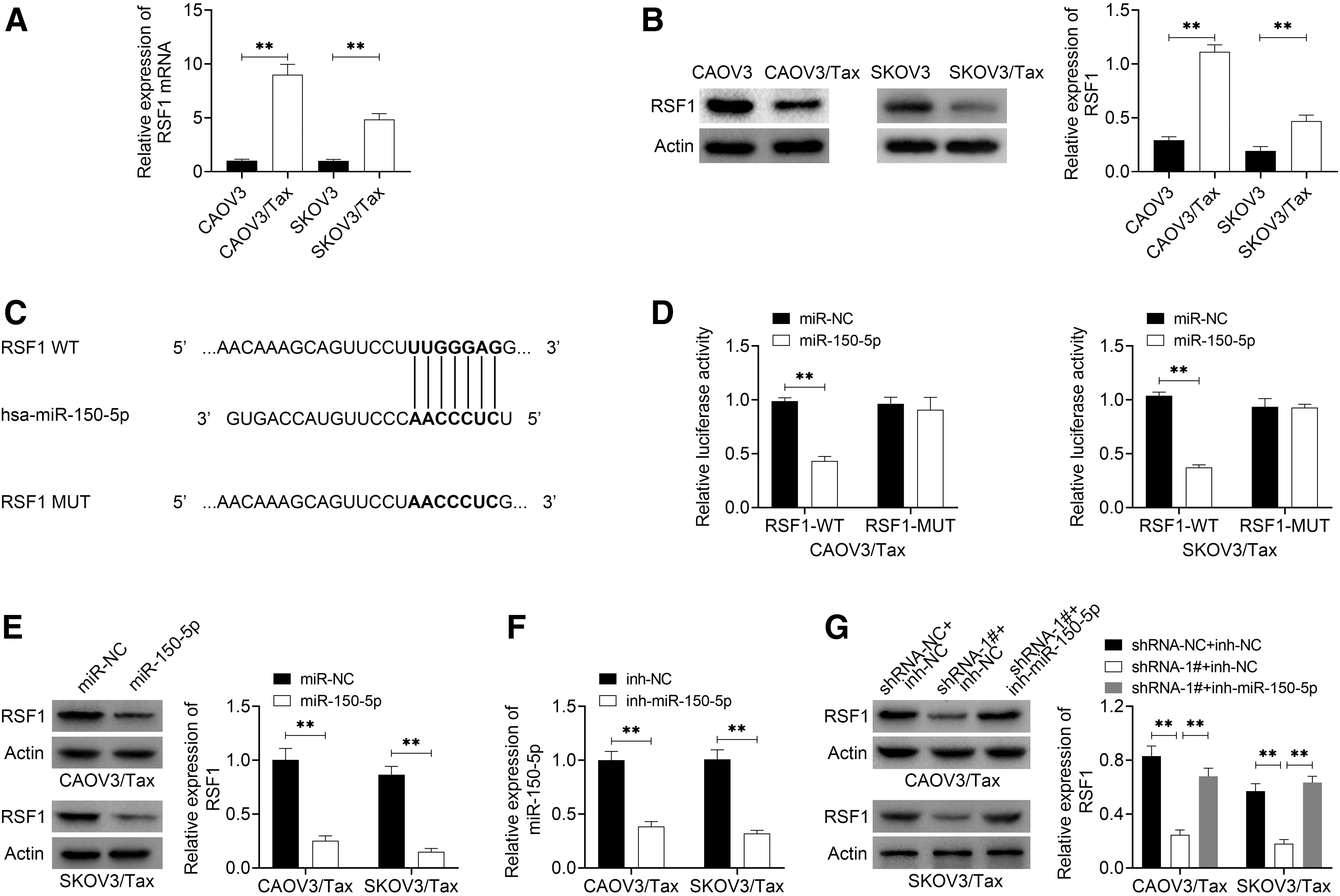
Downregulation of PRLB inhibited RSF1 expression by targeting miR-150-5p.
Knockdown of PRLB enhances the Tax sensitivity of CAOV3/Tax and SKOV3/Tax cells, at least in part, through inhibiting RSF1/NF-κB signaling
As the RSF1/NF-κB signaling plays a vital role in the chemoresistance in many kinds of cancers, including ovarian cancer, the authors then investigated whether PRLB was involved in the Tax resistance of ovarian cancer by modulating the RSF1/NF-κB signaling. The results showed that the IC50 decrease and cell apoptosis increase caused by PRLB downregulation were all significantly rescued when RSF1 was overexpressed in both CAOV3/Tax and SKOV3/Tax cell lines (Fig. 5A, B). In addition, RSF1 overexpression neutralized the decreases in the expression of RSF1 and p-p65 induced by PRLB downregulation (Fig. 5C). These findings illustrate that knockdown of PRLB enhanced the Tax sensitivity of CAOV3/Tax and SKOV3/Tax cells, at least in part, by inhibiting RSF1/NF-κB signaling.
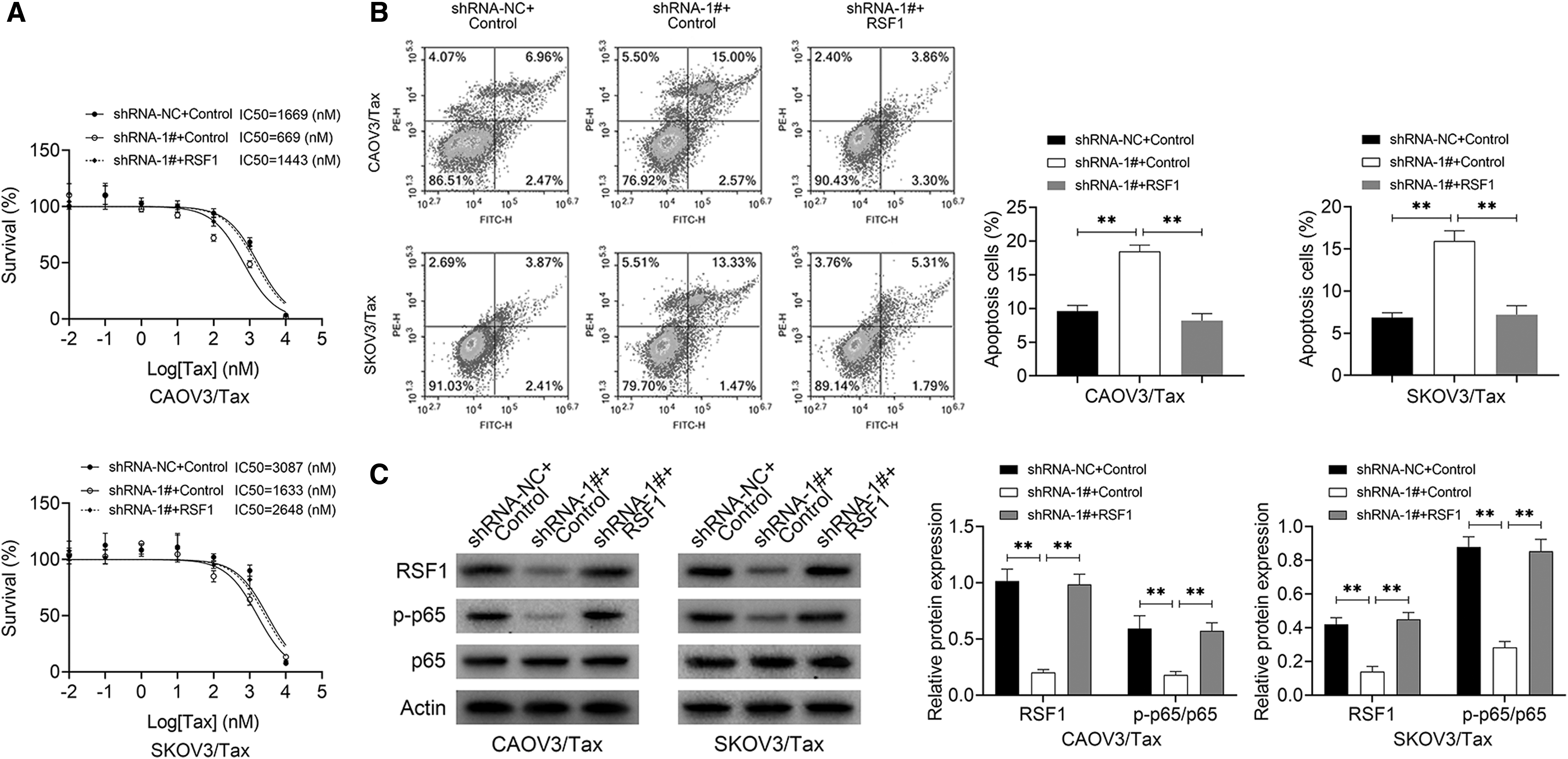
Knockdown of PRLB enhanced the Tax sensitivity of CAOV3/Tax and SKOV3/Tax cells, at least in part, by inhibiting RSF1/NF-κB signaling.
Discussion
EOC accounts for almost 90% of ovarian cancer cases in Asian populations. 21 Tax as a class of diterpenoid alkaloids generally serve as a first-line chemotherapy regimen for ovarian cancer. 22 However, the occurrence and development of drug resistance significantly reduce the curative effect and are a main reason for the poor prognosis of EOC patients. 23 In this study, through the in vitro experiments, the authors elucidated that the lncRNA PRLB was overexpressed in the Tax-resistant ovarian cancer tissues and cells, and downregulation of PRLB could induce a significant increase in the sensitivity of CAOV3/Tax and SKOV3/Tax cells to Tax by inhibiting RSF1/NF-κB signaling through targeting RSF1.
Currently, more lncRNAs have been identified as being involved in the drug resistance of ovarian cancer. 24 For instance, lncRNA HOTAIR was highly expressed in ovarian cancer cells. 25 LINC01118 was also detected to be overexpressed in the EOC tissues and chemoresistant cells, and LINC01118 upregulation significantly enhanced the resistance of ovarian cancer cells to Tax, as well as facilitated cell migration and invasion and reduced cell apoptosis. 26 Liu et al. 27 demonstrated that lncRNA FER1L4 expression was reduced in human ovarian cancer cells with drug resistance, and FER1L4 overexpression apparently increased the Tax sensitivity of ovarian cancer cells. In the current study, the authors revealed for the first time that lncRNA PRLB was overexpressed in the ovarian cancer tissues and cells with Tax resistance compared with the nonresistance tissues and cells, and knockdown of PRLB significantly increased the Tax sensitivity of ovarian cancer SKOV3 and CAOV3 cells.
miRNAs are a cluster of short ncRNAs, with probably 19–24 nucleotides in length. 28 The pre-miRNAs are cleaved to form the mature miRNAs in cytoplasm, followed by being integrated into the RNA-induced silencing complex and binding to the 3′-untranslated region (3′-UTR) of the target mRNA, resulting in the translational inhibition or degradation of the target mRNA. 29 Noticeably, lncRNAs could serve as miRNA sponges or decoys to repress the activities of target miRNAs, then intervening the interaction between the miRNAs and the target mRNAs. 30 The miR-4766-5p was identified to serve as a target of PRLB in breast cancer. 11 In this study, the authors identified that PRLB bonds to miR-150-5p, which could target the 3′-UTR of RSF1 mRNA. In NSCLC, miR-150-5p was confirmed to function as a target of linc00673, and downregulation of miR-150-5p abrogated the effects of linc00673 silence mediated repressions in cell proliferation, invasion, migration, and epithelial–mesenchymal transition, 31 suggesting that miR-150-5p plays an inhibitory role in NSCLC. MiR-150-5p was also found to serve as a tumor suppressor in head and neck squamous cell carcinoma by targeting SPOCK1. 32 In ovarian cancer, Xia et al. 33 reported that miR-150-5p was a target of the lncRNA ZFAS1, which promoted the EOC cell migration, invasion, and proliferation and enhanced the chemoresistance to cisplatin, whereas these effects were impaired by miR-150-5p, indicating that miR-150-5p could enhance the chemosensitivity of EOC cells to cisplatin. Moreover, RSF1 was reported to activate NF-κB signaling and then enhance the chemoresistance in ovarian cancer. 19 In this study, the authors verified that miR-150-5p regulated by PRLB could bind to RSF1 and inhibit the activation of RSF1/NF-κB signaling. Moreover, to deeply investigate the role of RSF1/NF-κB signaling in PRLB downregulation-mediated drug sensitivity enhancement, the authors also performed the rescued experiment. The authors found that overexpression of RSF1 abrogated the role of PRLB silence in the weakness of drug resistance and promotion of cell apoptosis, suggesting that RSF1/NF-κB is closely implicated in PRLB/miR-150-5p-mediated Tax resistance repression.
In conclusion, this study reveals that knockdown of PRLB improves the sensitivity of ovarian cancer cells to Tax, at least in part, through inhibiting the activation of RSF1/NF-κB signaling by targeting miR-150-5p. This study might provide an efficient target to prevent the resistance of ovarian cancer to Tax.
Ethics Approval and Consent to Participate
The experimental protocols that used human samples were approved by the Ethics Committee of Renmin Hospital of Wuhan University, and all the patients signed written informed consent.
Footnotes
Authors' Contributions
L.H. conceived and designed the experiments, Y.Z.Z. analyzed and interpreted the results of the experiments, and Y.Z.Z. and L.H. performed the experiments.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.