
Abstract
Background:
Breast cancer (BC) is a common tumor in women worldwide, and irradiation (IR) resistance is a major obstacle for BC therapy. Circle RNAs (circRNAs) were identified as implicated in the progression of cancer and IR resistance. However, the role of circABCB10 in BC progression and IR resistance is not well defined.
Materials and Methods:
The levels of circABCB10, miR-223-3p, and profilin-2 (PFN2) were detected by quantitative real-time polymerase chain reaction. The cell viability and survival rate were monitored by MTT assay. The glucose consumption, lactic acid production, LDH-A activity, and ATP production were evaluated to measure glycolysis. The protein levels of hypoxia inducible factor-1α (HIF-1α), hexokinase 2 (HK2), lactate dehydrogenase A chain (LDH-A), and PFN2 were estimated by Western blot assay. The colony formation rate was tested by colony formation assay. Dual-luciferase reporter assay was constructed to validate the interaction between miR-223-3p and circABCB10 or PFN2. The mice xenograft assay was performed to further verify the effects of circABCB10 on BC progression in vivo.
Results:
CircABCB10 and PFN2 were elevated, while miR-223-3p was reduced in BC tissues and cells. CircABCB10 sponged miR-223-3p, and PFN2 was a target of miR-223-3p in BC cells. CircABCB10 silencing inhibited cell proliferation, glycolysis, colony formation, and decreased IR resistance in BC cells by modulating miR-223-3p. Meanwhile, circABCB10 depletion restrained xenograft tumor growth in vivo. Also, miR-223-3p overexpression refrained cell proliferation, glycolysis, and colony formation while improving IR sensitivity in BC cells by regulating PFN2. Besides, circABCB10 knockdown declined PFN2 in BC cells via miR-223-3p. The glycolysis inhibitor 2-deoxy-D-glucose enhanced IR sensitivity in BC cells via circABCB10.
Conclusion:
CircABCB10 knockdown contributed to irradiation sensitivity by negatively regulating glycolysis via the miR-223-3p/PFN axis in breast cancer.
Introduction
Breast cancer (BC) is a common cancer among women, while there is still a small chance to happen in men. Despite the improvement in BC therapy, such as medication, radiation therapy, and surgery, it is still the second most lethal cancer worldwide. 1 Radiotherapy is an important treatment approach for BC patients, while the body would develop radioresistance with the increasing radiation time. 2 Therefore, it is crucial to elucidate the mechanism of irradiation (IR) resistance in BC.
Circular RNAs (circRNAs), a family of covalently closed loop structure RNAs without 3′-end and 5′-end, have been identified to play vital roles in diverse types of cancer development. 3 For example, Wu et al. reported that circ-ZNF609 modulated Gli1 expression to accelerate colorectal cancer progression via miR-150. 4 Another study implied that circ-ATP8A2 promoted cervical cancer progression through miR-433/EGFR axis. 5 Also, circRNAs were involved in IR resistance. For instance, a document in colon cancer indicated that circ_0001313 depletion increased IR sensitivity in colon cancer. 6 CircABCB10 (hsa_circ_0008717) was also implicated in BC progression. 7 However, the molecular mechanism of circABCB10 in BC progression and IR resistance was rarely reported.
The glycolytic metabolism in tumor cells also called “the Warburg effect”. It features increasing glucose uptake and accumulation of lactic acid in tumor cells. 8 There are reports that accumulation of lactic acid is an indication for tumor development, and also the aggravation for IR resistance in tumor cells. 9 Moreover, it was previously reported that the production of aerobic glycolysis was related to the development of radioresistance in cancer cells, hinting that the combination of glycolysis inhibitors has a remarkable therapeutic benefit in radiotherapy. 10 –12 Furthermore, the Warburg effect inhibitor 2-deoxy-D-glucose (2-DG) was identified as an effective drug to elevate IR sensitivity. 13,14 Synchronously, 2-DG and 6-aminonicotinamide (6-AN, an inhibitor of the pentose phosphate pathway) were confirmed to elevate radiosensitivity through activating ASK1/JNK/p38MAPK signaling pathway in neck cancer cells. 15,16 Hence, the development of effective glycolysis targeted drugs may provide a new therapeutic strategy to improve radiosensitivity of BC.
MicroRNAs (miRNAs), a form of small RNAs (18–25 nucleotides [nts]) without translation capacity, affect target gene expression by repressing mRNA translation or mediating mRNA degradation. 17 Also, current studies indicated that miRNAs were related to cancer progression and IR resistance. For example, a publication related to BC presented that miR-186 confined BC progression through modulating Twist1. 18 Another report showed that miR-33a-5p elevated IR sensitivity in melanoma through glycolytic metabolism. 19 Profilin-2 (PFN2), located on human chromosome 3q25.1, was identified to regulate actin polymerization in response to extracellular signals. 20 Besides, the aberrant expression of PFN2 was involved in BC progression. 21 However, the mechanisms of miR-223-3p and PFN2 in cancer progression and IR resistance have not been fully reported in BC.
In the current research, the authors explored the roles of circABCB10 in BC progression and IR resistance. This research may shed light on further study on IR resistance in BC.
Materials and Methods
Tissue sample collection
The study was permitted by the Ethics Committee of Guiyang Hospital of Guizhou Aviation Industry Group and performed according to the Declaration of Helsinki Principles. Thirty-three BC tissue samples were collected from BC patients in Guiyang Hospital of Guizhou Aviation Industry Group, as well as their corresponding adjacent normal tissue samples. All samples were frozen at −80°C until further use. Written informed consent was provided by all patients.
Cell culture and transfection
Four human BC cell lines MCF-7, MDA-MB-231, MDA-MB-468, and MDA-MB-453 and normal breast epithelial cell line MCF-10A were purchased from BioVector NTCC, Inc. (Beijing, China). The cells were cultured in minimum essential medium (MEM; Solarbio, Beijing, China) containing 10% fetal bovine serum (FBS; Solarbio) and incubated in an incubator with 5% CO2 at 37°C.
Small interfering RNA (siRNA) targeting circABCB (si-circABCB10, 5′- TAGAAGACCATGGGGGATGTCAAGAGCATCCCCCATGGTCTTCTATTTTTT-3′) and its negative control (si-NC), miR-223-3p mimics (miR-223-3p) and its control (miR-NC), and miR-223-3p inhibitor (anti-miR-223-3p) and its matched control (anti-miR-NC) were obtained from GenePharma (Shanghai, China). The fragments of circABCB10 and PFN2 were amplified and then inserted into pLCDH-cir (pLCDH; Geneseed, Guangzhou, China) or pcDNA (pcDNA-NC; Invitrogen, Carlsbad, CA) vector to construct overexpression plasmid, namely circABCB10 and pcDNA-PFN2, respectively. Then, 50 nM of synthetic oligonucleotides or vectors were transfected into MCF-7 and MDA-MB-231 cells using Lipofectamine 3000 (Invitrogen) according to the manufacturer's manual.
Quantitative real-time polymerase chain reaction
The RNA in BC tissue samples and cells was extracted using TriQuick Reagent (Solarbio). The random primers (TaKaRa, Dalian, China) were used for reverse transcription of circABCB10 and PFN2, and miRNA 1st Strand cDNA Synthesis Kit (Vazyme, Nanjing, China) was utilized for reverse transcription of miR-223-3p. The quantitative PCR was performed using SYBR Premix Ex Taq II (TaKaRa). The level of miR-223-3p was normalized by small nuclear RNA U6, while circABCB10 and PFN2 were analyzed by glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as control, and then calculated with the method of 2−ΔΔCt. The oligonucleotides were bought from Beijing Genomics Institute (BGI, Shenzhen, China) and listed as follows: circABCB10: (F, 5′-CTAAGGAGTCACAGGAAGACATC-3′, and R, 5′-GTAGAATCTCTCAGACTCAAGGTTG-3′); miR-223-3p: (F, 5′-CGCUAUCUUUCUAUUAACUGACCAUAA-3′, and R, 5′-CGCUAUCUUUCUAUUAUGACUCCAUAA-3′); PFN2: (F, 5′-ATGATTGTAGGAAAAGACCGGGA-3′, and R, 5′-GCAGTCACCATCGACGTATAGAC-3′); GAPDH: (F, 5′-TGTTCGTCATGGGTGTGAAC-3′, and R, 5′-ATGGCATGGACTGTGGTCAT-3′); and U6: (F, 5′-ATTGGAACGATACAGAGAAGATT-3′, and R, 5′-GGAACGCTTCACGAATTTG-3′).
3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide assay
MCF-7 and MDA-MB-231 cells were injected into a 96-well plate at the density of 6 × 103 per well and incubated overnight at 37°C. Subsequently, the cells were incubated with 0.5 mg/mL MTT for 4 h at 37°C. Then, 100 μL of dimethyl sulfoxide (DMSO; Solarbio) was added to solubilize the formazan. The absorbance at 490 nm was detected using a microplate reader. Each sample was prepared in triplicate.
Measurement of glycolysis
The glucose consumption, lactic acid production, LDH-A activity, and ATP production were monitored using glucose assay kit (Solarbio), LA assay kit (Solarbio), LDH activity assay kit (Solarbio), and ATP detection kit (Solarbio), respectively. All protocols were performed according to the manual.
Western blot assay
The protein in the BC tissues and cells was extracted using RIPA buffer (Solarbio). The concentration of protein samples was assessed by a BCA Kit (Beyotime, Shanghai, China). After the denaturation at 100°C for 5 min, the equal amount of 20 μg protein samples were separated by sodium dodecyl sulfonate-polyacrylamide gel electrophoresis, and then transferred onto a polyvinylidene fluoride (PVDF) membrane (Solarbio). Then the membrane was sealed in the nonfat milk for 2 h at room temperature and incubated with primary antibody against hypoxia inducible factor-1α (HIF-1α, 1/2000; Abcam, Cambridge, MA), hexokinase 2 (HK2, 1/1000; Abcam), lactate dehydrogenase A chain (LDH-A, 1/1000; Abcam), PFN2 (1/2000; Abcam), and GAPDH (1/10,000; Abcam) at 4°C overnight, and then maintained with goat anti-rabbit IgG-HRP secondary antibody (1/20,000; Abcam) at 37°C for 2 h. The protein signaling was visualized via eyoECL plus kit (Beyotime).
IR and drug treatment
A Siemens Primus linear accelerator (Bolin, Germany) was utilized for X-ray irradiation. The MCF-7 and MDA-MB-231 cells (6 × 103 cell per well) were transfected with oligonucleotides or vectors for 48 h, and then treated with 0, 2, 4, 6, and 8 Gy/min of X-rays irradiation at 100 cm.
The glycolysis inhibitor 2-DG (Solarbio) was dissolved in deionized water to 5 mM. The MCF-7 and MDA-MB-231 cells were disposed with dilute 2-DG for 24 h.
Colony formation assay
The transfected MCF-7 and MDA-MB-231 cells were added into six-well plate (500 cells per well) containing MEM and 10% FBS, and cultivated at 37°C for 15 d. The fresh medium was replaced for every 3 d. Then, the colonies were stained with crystal violet (Sigma-Aldrich, St. Louis, MO) for 30 min at room temperature and fixed with 4% paraformaldehyde. The number of colonies with >50 cells was counted.
Bioinformatics analysis
The interaction between miR-223-3p and circABCB10 or PFN2 was predicted by starBase v3.0 online database.
Dual-luciferase reporter assay
The dual-luciferase reporter assay was performed using pmirGLO vector (Youbio, Changsha, China). The wild-type sequences containing binding sites and mutant sequences of circABCB10 and 3′-untranslated regions (3′UTR) of PFN2 were amplified and inserted into pmirGLO vector to construct luciferase reporter, named as circABCB10-WT, circABCB10-MUT, PFN2 3′UTR-WT, and PFN2 3′UTR-MUR, respectively. The BC cells were cotransfected with 100 ng corresponding luciferase reporter and 40 nM miR-NC or miR-223-3p using Lipofectamine 3000 (Invitrogen). After transfection for 48 h, Dual-Lucy Assay Kit (Solarbio) was used for luciferase activity analysis. The Renilla activity was used as internal reference.
In vivo experiment
The experiment in nude mice was authorized by the Animal Care Committee of Guiyang Hospital of Guizhou Aviation Industry Group and performed according to the guidelines of National Institutes of Health. The short hairpin RNA (shRNA) against circABCB10 (sh-circABCB10) and its matched negative control (sh-NC) were synthesized from Genechem (Shanghai, China). The nude mice were obtained from Shanghai Laboratory Animal Company (SLAC, Shanghai, China). The MCF-7 cells (3 × 106) stably transfected with sh-NC or sh-circABCB10 were implanted into nude mice (n = 5 per group), and the volume of xenograft tumor was measured every week for 5 weeks according to the formula: volume (mm3) = width 2 × length/2. Weight measurement was first taken for the resected xenograft tumor and then stored in −80°C for further research.
Statistical analysis
All data are exhibited as the means ± standard deviation (SD) from three independent experiments using GraphPad Prism 7 (GraphPad, Inc., San Diego, CA). The comparisons between two groups were analyzed by Student's t-test, while among multiple groups, they were processed via one-way analysis of variance (ANOVA) followed by Dunnett's test. The difference was considered as statistically significant when p value was less than 0.05.
Results
CircABCB10 was elevated in BC tissues and cells
To explore the roles of circABCB10 in BC, the level of circABCB10 was first examined in BC tissues. As shown in Figure 1A, the level of circABCB10 was increased in BC tissue samples compared with that in normal tissue samples. Also, circABCB10 was upregulated in BC cell lines (MCF-7, MDA-MB-231, MDA-MB-468, and MDA-MB-453) related to that in normal breast epithelial cell line MCF-10A (Fig. 1B). These results implicated that circABCB10 was increased in BC tissues and cells.
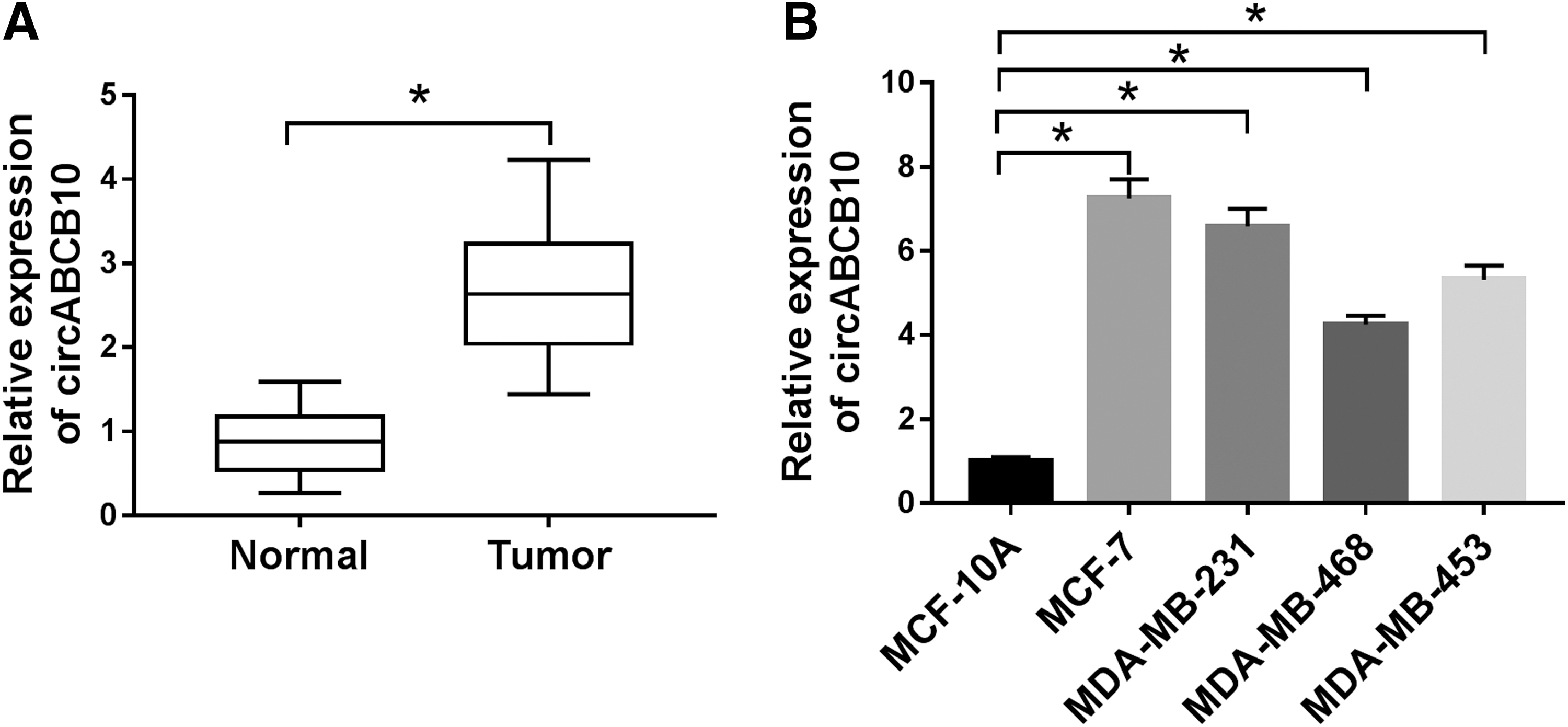
CircABCB10 was elevated in BC.
CircABCB10 silencing confined cell proliferation, glycolysis, and colony formation, and improved IR sensitivity in BC cells
Based on the above results, the authors further explored the functions of circABCB10. As presented in Figure 2A and B, the level of circABCB10 was declined in MCF-7 and MDA-MB-231 cells transfected with si-circABCB10 in comparison with that in si-NC group, suggesting the high knockdown efficiency. Furthermore, the transfection of si-circABCB10 resulted in the reduction of cell viability in MCF-7 and MDA-MB-231 cells (Fig. 2C, D). Meanwhile, glucose consumption, lactic acid production, LDH-A activity, and ATP production were all decreased in MCF-7 and MDA-MB-231 cells transfected with si-circABCB10 compared with that in si-NC group (Fig. 2E–H). Since HIF-1α, HK2, and LDH-A are glycolysis-associated factors, 22 the levels of these proteins were detected in si-circABCB10-transfected MCF-7 and MDA-MB-231 cells. As exhibited in Figure 2I and J, the protein levels of HIF-1α, HK2, and LDH-A were all reduced in si-circABCB10 group. In addition, the survival rate of MCF-7 and MDA-MB-231 cells was gradually decreased in si-circABCB10 group under IR condition compared to that in si-NC group (Fig. 2K, L). Also, the transfection of si-circABCB10 or IR treatment downregulated colony formation rate of MCF-7 and MDA-MB-231 cells, and IR aggravated the constraint effect on colony formation rate induced by si-circABCB10 (Fig. 2M, N). These data demonstrated that circABCB10 silencing impeded cell proliferation, glycolysis, and colony formation, but enhanced IR sensitivity in MCF-7 and MDA-MB-231 cells.
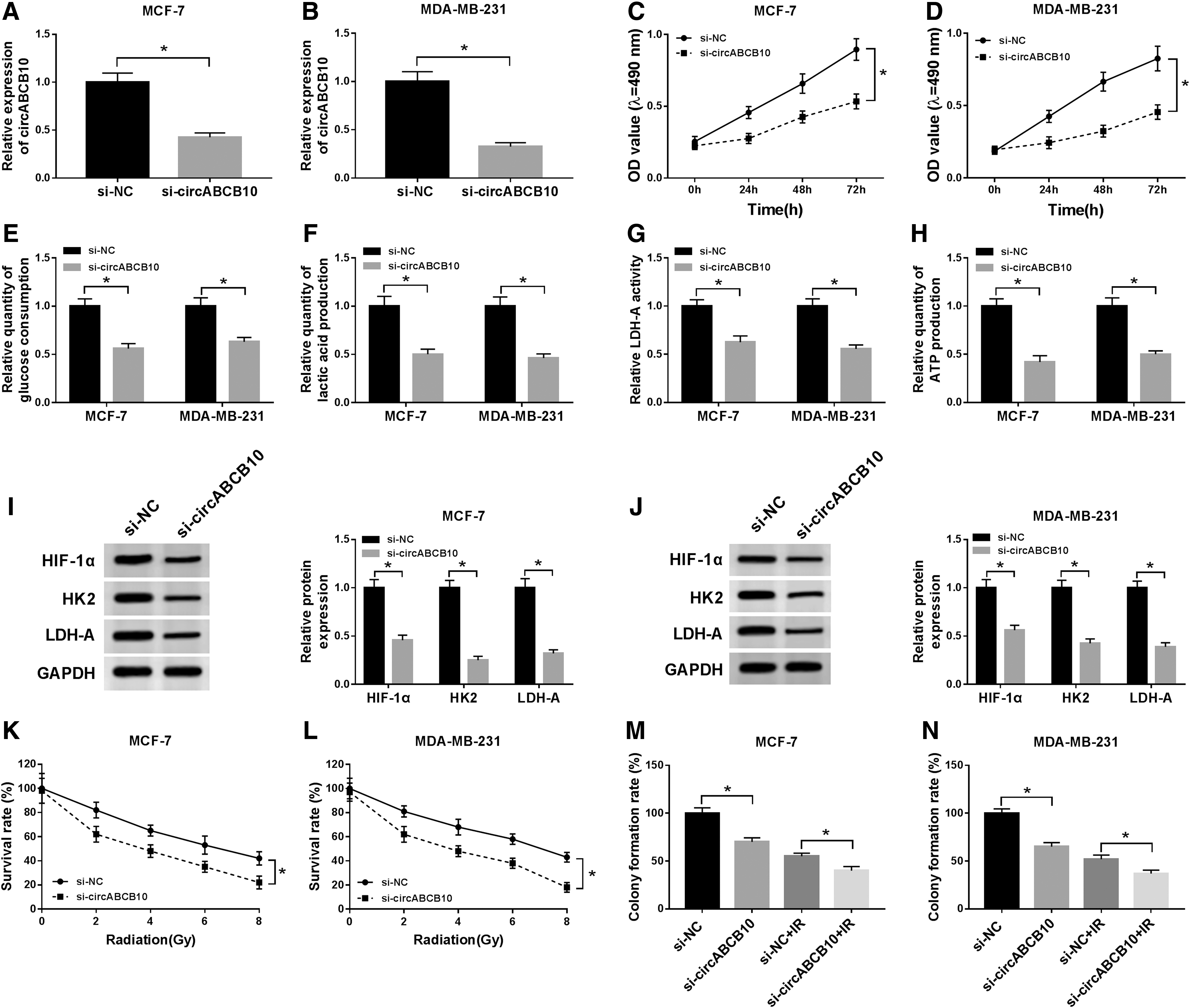
CircABCB10 silencing confined cell proliferation, glycolysis, and colony formation, and improved IR sensitivity in BC cells.
MiR-223-3p was targeted by circABCB10
To further explore the molecular mechanism of circABCB10 in BC, starBase v3.0 was utilized to search the putative target of circABCB10. As exhibited in Figure 3A, miR-223-3p had complementary base pairing with circABCB10. Following dual-luciferase reporter assay indicated that the luciferase activity of circABCB10-WT was declined in MCF-7 and MDA-MB-231 cells transfected with miR-223-3p compared with that miR-NC group; however, the luciferase activity of circABCB10-MUT reporter had no fluctuation in any group (Fig. 3B, C). Besides, the level of miR-223-3p was reduced in BC tissues and cell lines (Fig. 3D, E). Also, the level of miR-223-3p was negatively correlated with circABCB10 (Fig. 3F). In addition, miR-223-3p was enhanced in si-circABCB10-transfected MCF-7 and MDA-MB-231 cells, while decreased in circABCB10 overexpression group (Fig. 3G–J). These data unraveled that circABCB10 sponged miR-223-3p in BC cells.
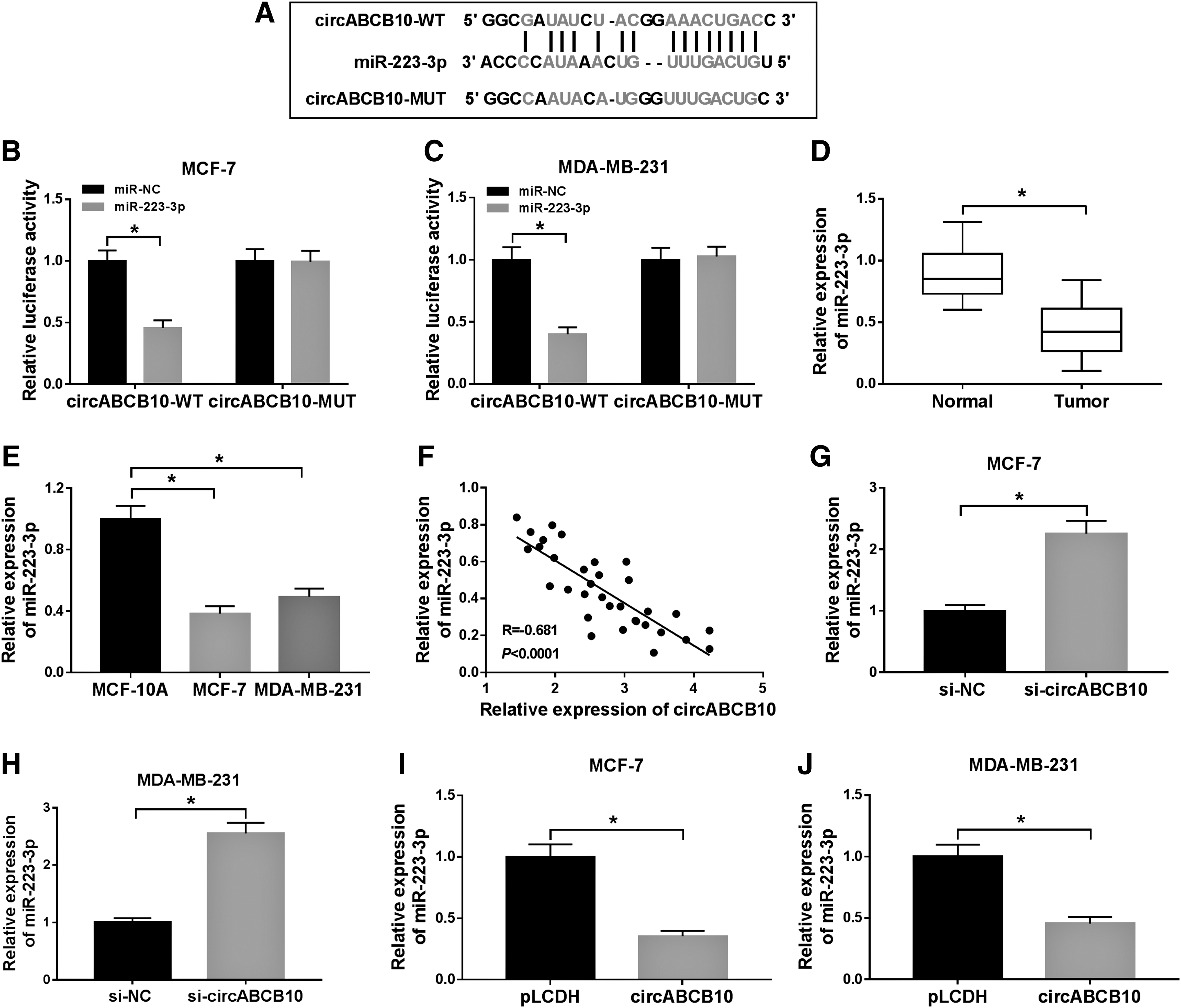
MiR-223-3p acted as a target of circABCB10.
CircABCB10 silencing refrained cell proliferation, glycolysis, and colony formation, but enhanced IR sensitivity in BC cells by regulating miR-223-3p
To elucidate whether circABCB10-induced cell proliferation, glycolysis, colony formation, and IR resistance in BC was mediated by miR-223-3p, the restoration experiment of miR-223-3p was performed. As shown in Figure 4A and B, the level of miR-223-3p was upregulated in si-circABCB10-transfected MCF-7 and MDA-MB-231 cells, while receded by the reintroduction of anti-miR-223-3p. Furthermore, the transfection of anti-miR-223-3p regained the restraint effects on cell viability, glucose consumption, LDH-A activity, the productions of lactic acid and ATP, and also the protein levels of HIF-1α, HK2, and LDH-A in MCF-7 and MDA-MB-231 cells retarded by si-circABCB10 (Fig. 4C–J). Besides, the survival rate of MCF-7 and MDA-MB-231 cells was declined in si-circABCB10 group with IR treatment, while the emergence of anti-miR-223-3p reversed this inhibitory impact (Fig. 4K, L). As presented in Figure 4M and N, IR inhibited the colony formation rate of MCF-7 and MDA-MB-231 cells as compared with the normal group; however, colony formation rate was reduced in MCF-7 and MDA-MB-231 cells transfected with si-circABCB10, but restored by the reintroduction of anti-miR-223-3p. These data revealed that circABCB10 depletion repressed cell proliferation, glycolysis, and colony formation, as well as increased IR sensitivity in BC cells by regulating miR-223-3p.
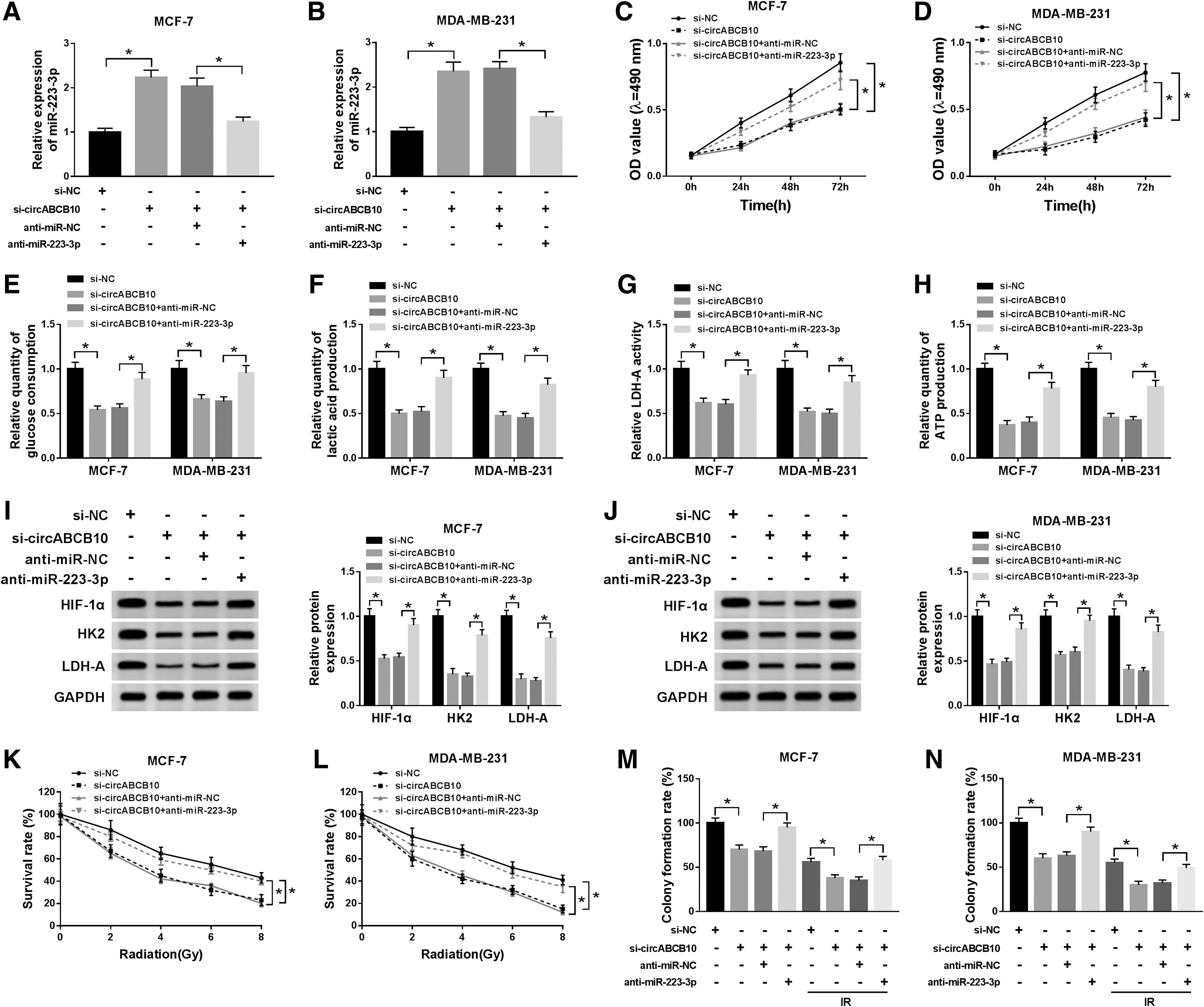
CircABCB10 silencing refrained cell proliferation, glycolysis, and colony formation, and enhanced IR sensitivity by regulating miR-223-3p in BC cells.
PFN2 was a target of miR-223-3p
To further illustrate the molecular mechanism of miR-223-3p in BC, starBase v3.0 database was used to search the putative targets of miR-223-3p. As shown in Figure 5A, PFN2 had complementary binding sequences with miR-223-3p. Meanwhile, the introduction of miR-223-3p contributed to the downregulation of luciferase activity of PFN2 3′UTR-WT reporter in MCF-7 and MDA-MB-231 cells in contrast to that of in miR-NC group; whereas the luciferase activity of PFN2 3′UTR-MUT reporter had no apparent change in any group (Fig. 5B, C). Besides, the levels of PFN2 mRNA and protein were upregulated in BC tissues and cells compared with that in normal tissues or MCF-10A cells (Fig. 5D, F, G). The scatter plot indicated that the level of miR-223-3p was negatively correlated with PFN2 (Fig. 5E). In addition, the mRNA and protein levels of PFN2 were dramatically upregulated in anti-miR-223-3p-transfected MCF-7 and MDA-MB-231 cells, but notably decreased in miR-223-3p group (Fig. 5H, I). These data disclosed that PFN2 was a direct candidate target of miR-223-3p in BC cells.
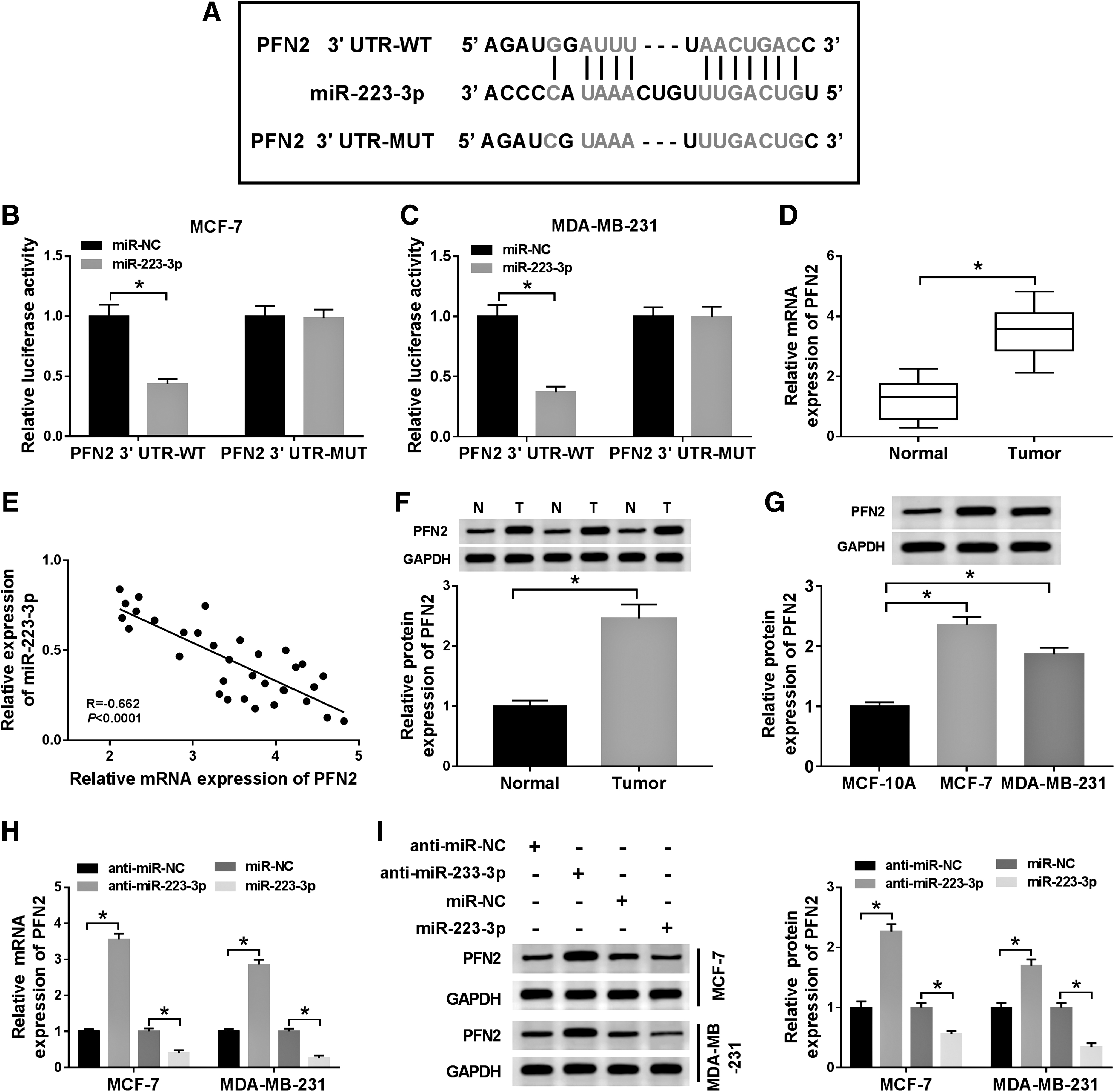
PFN2 was a target of miR-223-3p.
MiR-223-3p overexpression curbed cell proliferation, glycolysis, colony formation, and IR resistance in BC cells by regulating PFN2
Based on the above results, the authors found that PFN2 was a candidate target of miR-223-3p in BC cells. Next, they further explored whether miR-223-3p-stimulated cell proliferation, glycolysis, colony formation, and IR resistance in BC was mediated by PFN2. First, the levels of PFN2 mRNA and protein were reduced in MCF-7 and MDA-MB-231 cells transfected with miR-223-3p, but rescued by the reintroduction of pcDNA-PFN2 (Fig. 6A–D). Furthermore, the emergence of pcDNA-PFN2 restored the inhibitory effect on cell viability in MCF-7 and MDA-MB-231 cells retarded by miR-223-3p (Fig. 6E, F). Meanwhile, the levels of glucose consumption, LDH-A activity, the productions of lactic acid, and ATP were downregulated in miR-223-3p-transfected MCF-7 and MDA-MB-231 cells, while relieved in miR-223-3p + pcDNA-PFN2 group (Fig. 6G–J). Also, the protein levels of HIF-1α, HK2, and LDH-A showed the same trend. In brief, the transfection of miR-223-3p resulted in decreased levels of these proteins in MCF-7 and MDA-MB-231 cells, while alleviated by the emergence of pcDNA-PFN2 (Fig. 6K, L). In addition, the cell survival rate was downregulated in IR-treated MCF-7 and MDA-MB-231 cells transfected with miR-223-3p, while regained by the reintroduction of pcDNA-PFN2 (Fig. 6M, N). IR inhibited colony formation rate of MCF-7 and MDA-MB-231 cells compared with that in normal group; meanwhile, the introduction of pcDNA-PFN2 mitigated the suppressive impact on colony formation rate of MCF-7 and MDA-MB-231 cells caused by miR-223-3p (Fig. 6O, P). These results implicated that miR-223-3p inhibited cell proliferation, glycolysis, and colony formation, while enhanced IR sensitivity in BC cells by modulating PFN2.

MiR-223-3p curbed cell proliferation, glycolysis, colony formation, and IR resistance in BC cells by regulating PFN2.
CircABCB10 knockdown reduced PFN2 expression in BC cells by sponging miR-223-3p
To elucidate the relationship among circABCB10, miR-223-3p, and PFN2 in BC, si-circABCB10 and anti-miR-223-3p were cotransfected into MCF-7 and MDA-MB-231 cells. As presented in Figure 7A–D, the levels of PFN2 mRNA and protein were declined in si-circABCB10-transfected MCF-7 and MDA-MB-231 cells, while the emergence of anti-miR-223-3p counteracted this suppressive effect. Besides, the level of circABCB10 was positively correlated with PFN2 (Fig. 7E). These results showed that circABCB10 silencing decreased PFN2 in BC cells via miR-223-3p.
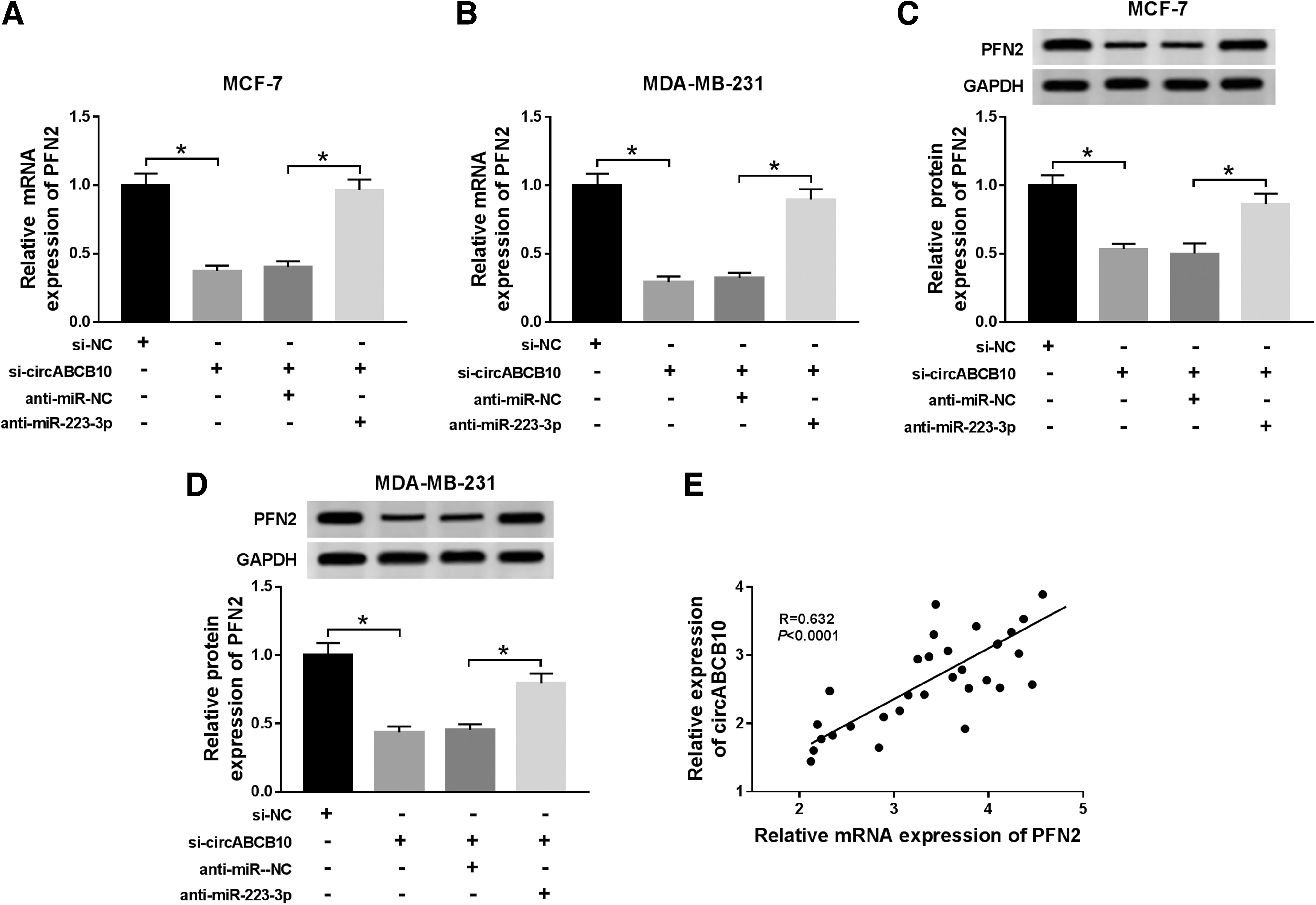
CircABCB10 knockdown reduced PFN2 expression in BC cells by sponging miR-223-3p.
CircABCB10 depletion constrained xenograft tumor growth in vivo
To further explore the effects of circABCB10 in BC, MCF-7 cells stably transfected with sh-NC or sh-circABCB10 were implanted into nude mice. As exhibited in Figure 8A and B, the volume and weight of xenograft tumor were both declined in sh-circABCB10 group related to that in sh-NC group. Meanwhile, the level of circABCB10 was reduced, while miR-223-3p was upregulated in sh-circABCB10 group compared to that in sh-NC group (Fig. 8C, D). Also, the protein level of PFN2 was decreased in sh-circABCB10 group (Fig. 8E). To sum up, these results unraveled that the depletion of circABCB10 blocked xenograft tumor growth in vivo.
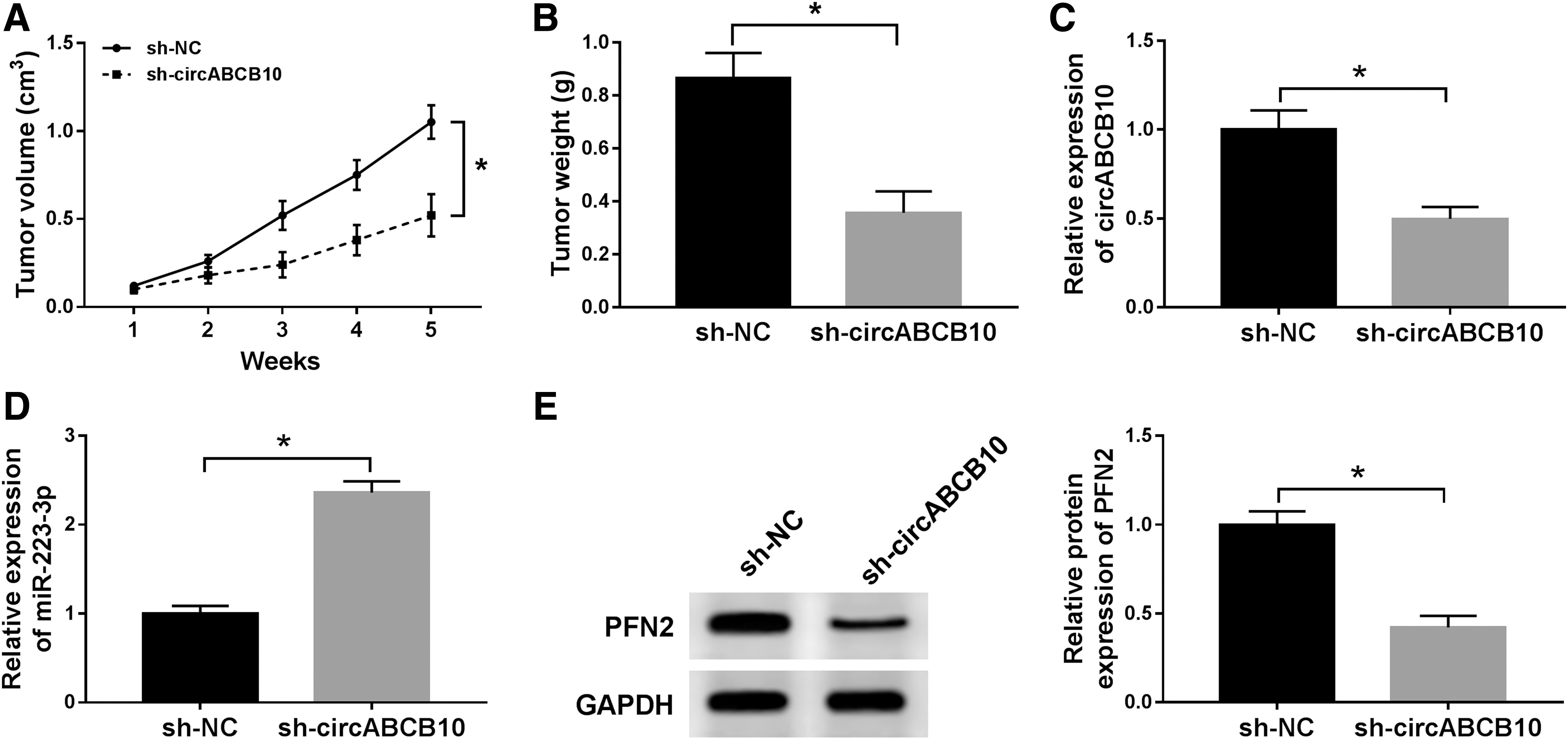
CircABCB10 depletion constrained xenograft tumor growth in vivo.
Inhibition of glycolysis attenuated the promoted effects on IR resistance in BC cells induced by circABCB10
To further investigate the effects of glycolysis on IR resistance in BC, the glycolysis inhibitor 2-DG was used to treat the MCF-7 and MDA-MB-231 cells transfected with circABCB10. As exhibited in Figure 9A, 2-DG had no influence on the survival rate of MCF-7 and MDA-MB-231 cells. Moreover, the treatment of 2-DG weakened the promoted impact on the levels of glucose consumption, LDH-A activity, and the productions of lactic acid and ATP in MCF-7 and MDA-MB-231 cells induced by circABCB10 (Fig. 9B–E). Also, the levels of HIF-1α, HK2, and LDH-A protein also showed the similar tendency. In brief, the levels of these protein were upregulated in MCF-7 and MDA-MB-231 cells transfected with circABCB10, but the treatment of 2-DG abated this accelerated effect (Fig. 9F, G). In addition, 2-DG attenuated the promoted effect on survival rate of MCF-7 and MDA-MB-231 cells under IR condition facilitated by circABCB10 (Fig. 9H, I). Also, 2-DG diminished the accelerated impact on colony formation rate of MCF-7 and MDA-MB-231 cells caused by circABCB10 in normal condition or IR condition (Fig. 9J, K). These data disclosed that glycolysis inhibition attenuated the facilitated effects on colony formation and IR resistance in BC cells promoted by circABCB10.
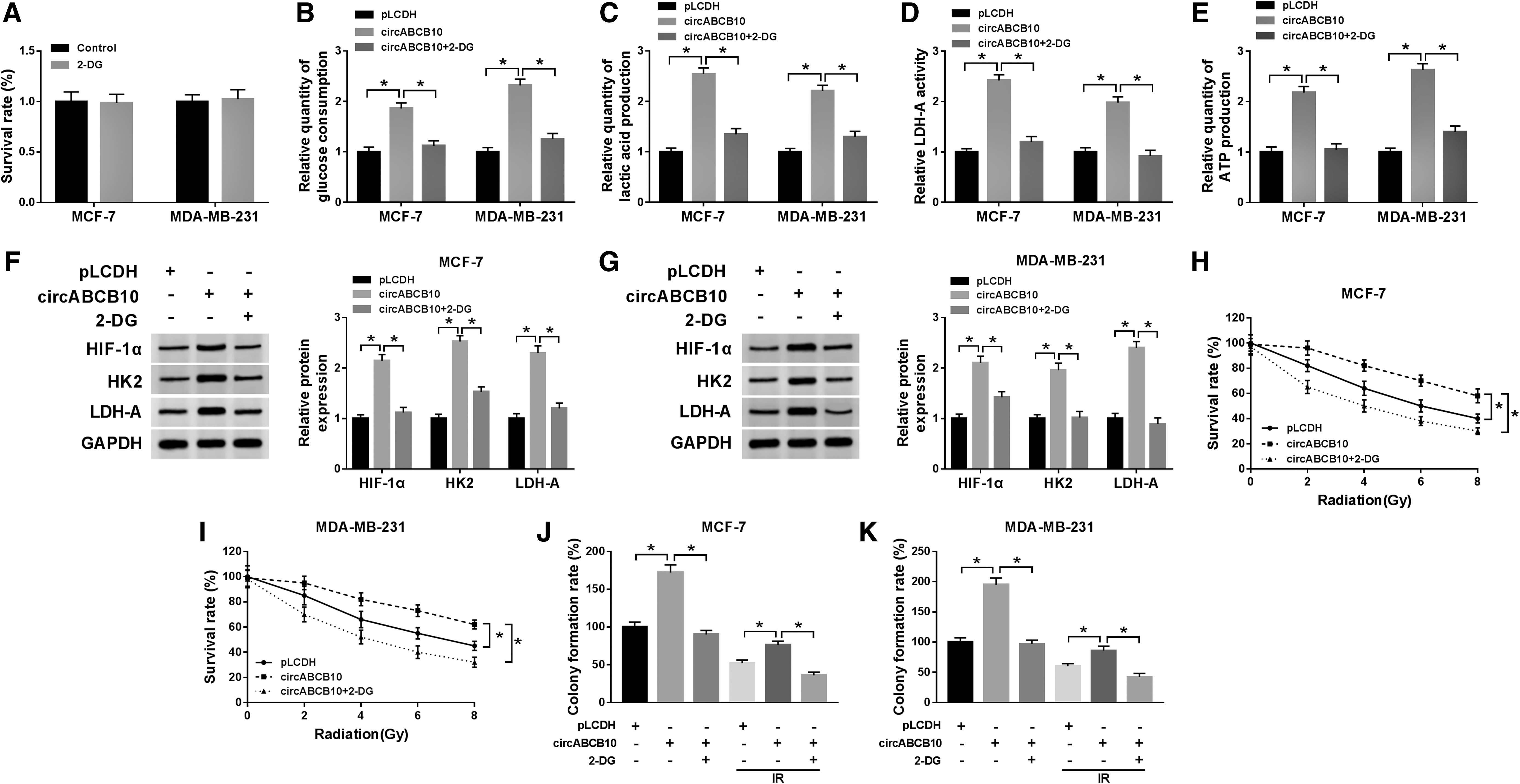
Inhibition of glycolysis attenuated the promoted effects on colony formation and IR resistance in BC cells induced by circABCB10.
Discussion
The progression of cancer and IR resistance were the main barriers to successful treatment for cancer patients. Emerging data indicated that circRNAs played vital roles in cancer progression and IR resistance. 23,24 In this research, the authors focused on the effects of circABCB10 on BC progression and IR resistance. These results unraveled that circABCB10 upregulated PFN2 expression to accelerate BC progression, and glycolysis to aggravate IR resistance by sponging miR-223-3p.
Recent studies indicated that circRNAs were related to the development and progression of cancer. For example, Ding et al. documented that circ-DONSON was upregulated in gastric cancer, and circ-DONSON accelerated gastric cancer progression through recruiting the NURF complex. 25 Also, another study in nonsmall cell lung cancer (NSCLC) presented that circABCB10 silencing retarded cell proliferation and migration in NSCLC cells through miR-1252/FOXR2 axis. 26 In the current study, circABCB10 was elevated in BC tissues and cells. Furthermore, circABCB10 knockdown hindered cell growth in BC cells. The similar results of circABCB10 in BC were in line with that in previous document. 7 In addition, circABCB10 silencing blocked xenograft tumor growth in vivo. These data indicated that circABCB10 depletion confined BC progression.
The glycolysis in tumor cells is different from that in normal cells, and termed “the Warburg effect”. Its main characteristics are the increase of glucose uptake and lactate output in tumor cells. 27 The dysregulation of circRNAs was implicated in glycolytic metabolism in many types of cancers. For example, a study in oral squamous cell carcinoma (OSCC) revealed that circRNA_100290 knockdown reduced the lactate production and glucose uptake in OSCC cells via miR-378a. 28 Another report in gallbladder cancer (GBC) exhibited that circFOXP1 accelerated the Warburg effect in GBC cells. 29 In this research, circABCB10 depletion decreased the levels of glucose consumption, LDH-A activity, and the productions of lactic acid and ATP production in BC cells. Moreover, the levels of HIF-1α, HK2, and LDH-A also showed the same tendency. Besides, the Warburg effect inhibitor 2-DG reversed the promoted effect on glycolysis in BC cells induced by circABCB10. IR resistance is one of the major problems in cancer treatment. Emerging evidence implied that glycolysis contributed to IR resistance of cancer cells. 30 Kamenisch et al. documented that IR promoted invasion in melanoma through enhancing the Warburg effect. 31 In the present exploration, the silencing of circABCB10 reduced the survival rate of BC cells in IR condition. Furthermore, the IR treatment aggravated the inhibitory effect on colony formation rate of BC cells induced by circABCB10. In addition, 2-DG declined colony formation rate and IR resistance in BC cells inhibited by circABCB10. The similar results of 2-DG in IR sensitivity were consistent with the reported documents. 10,11 These data disclosed that circABCB10 facilitated IR resistance through promoting glycolysis in BC.
Convincing evidence showed that the ectopic expression of miR-223-3p was related to cancer progression. For instance, a document displayed that miR-223-3p refrained cell proliferation and migration in lung squamous cell carcinoma. 32 In the present study, miR-223-3p was sponged by circABCB10. Meanwhile, miR-223-3p was downregulated in BC tissues and cells. Furthermore, circABCB10 knockdown constrained cell proliferation and colony formation of BC cells by regulating miR-223-3p. The similar results of miR-223 in BC were consistent with the past report. 33,34 Accumulating evidence showed that miRNAs were associated with IR resistance. For example, a document in NSCLC implied that miR-99a improved IR sensitivity in NSCLC cells through mTOR signaling pathway. 35 In this study, the silencing of circABCB10 suppressed glycolysis, while improved IR sensitivity in BC cells by modulating miR-223-3p. These data demonstrated that circABCB10 silencing repressed BC progression and glycolysis, as well as mitigated IR resistance by sponging miR-223-3p.
Recent studies demonstrated that PFN2 was also implicated in cancer progression. For example, Zhou et al. reported that PFN2 facilitated head and neck cancer progression. 36 Also, Kim et al. exhibited that adhesion-related molecule PFN2 was enriched in the integrin and epithelial–mesenchymal transition signaling pathway which associated with radioresistant. 37 In the current research, PFN2 was a target of miR-223-3p. MiR-223-3p overexpression curbed cell proliferation, colony formation, and glycolysis, while improved IR sensitivity in BC cells by regulating PFN2. The similar results of PFN2 in BC progression were in agreement with that in previous report. 21 Besides, circABCB10 depletion declined PFN2 expression in BC cells by sponging miR-223-3p. These data uncovered that circABCB10 depletion reduced PFN2 expression to repress BC progression and glycolysis, while enhanced IR sensitivity by sponging miR-223-3p.
In this study, the limitation was the limited sample size. In future exploration, the more BC tissue samples will be collected. Also, studies with nude mice under IR condition will be carried out to validate the effects of circABCB10 on IR sensitivity.
Conclusion
In conclusion, circABCB10 and PFN2 were increased, but miR-223-3p was declined in BC tissues and cells. The authors' findings discovered that circABCB10 knockdown could facilitate IR sensitivity through negatively modulating glycolysis via the miR-223-3p/PFN axis in BC, providing a novel direction for radiotherapy of BC.
Footnotes
Authors' Contributions
Conceptualization and Methodology: Rui Zhong and Chaoyue Deng; Formal analysis and Data curation: Chaoyue Deng and Zhenlin Zhou; Validation and Investigation: Yue Zhao and Zhenlin Zhou; Writing - original draft preparation and Writing - review and editing: Yue Zhao, Rui Zhong and Chaoyue Deng; Approval of final article: all authors
Availability of Data and Materials
The analyzed data sets generated during the present study are available from the corresponding author on reasonable request.
Ethics Approval and Consent to Participate
The present study was approved by the Ethics Review Committee of Guiyang Hospital of Guizhou Aviation Industry Group. Written informed consent was obtained from all enrolled patients.
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
No funding was received for this article.