
Abstract
Background:
Lung cancer was one of the most common malignant tumors worldwide. In China, lung cancer has become the leading reason of malignant tumors-related mortality in urban population, whereas nonsmall cell lung cancer (NSCLC) represented at least 80% of all lung cancers with poor 5-year survival rate. Long noncoding RNA (lncRNA) small nucleolar RNA host gene 20 (SNHG20) was reported to be associated with NSCLC, but the regulatory mechanisms of SNHG20 in NSCLC needed further investigation.
Methods:
The abundances of SNHG20 and E2F transcription factor 3 (E2F3) in NSCLC tissues and cells were measured with real-time quantitative polymerase chain reaction (RT-qPCR) and Western blot assays. 3-(4, 5-Dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazol-3-ium bromide (MTT) was applied to detect cells proliferation, whereas flow cytometry analysis was used to monitor cell apoptosis. In addition, cells capabilities of migratory and invasion were assessed with transwell assay. The association among miR-2467-3p, SNHG20, and E2F3 was analyzed by dual-luciferase reporter assay. The related protein expression levels were determined by Western blot.
Results:
SNHG20 and E2F3 was upregulation in NSCLC tissues and cell lines. Mechanical experiment displayed that knockdown of SNHG20 or E2F3 silencing could inhibit proliferation, motility, and improve apoptosis in NSCLC cell lines. Restored expression of E2F3 could effectively reverse reduction of proliferation, motility, and promotion of apoptosis caused by SNHG20 silencing in NSCLC cells. Besides, SNHG20 activated protein kinase B (AKT) signaling pathway and increased E2F3 level in NSCLC cells through targeting miR-2467-3p.
Conclusion:
SNHG20 contributed to NSCLC development through mediating AKT signaling pathway and sponging miR-2467-3p to elevate E2F3 expression in NSCLC cells.
Introduction
Lung cancer was the leading reason of cancer-induced mortality and more predominantly diagnosed cancers worldwide, which contributed to ∼1.2 million deaths from 2005 to 2015. 1 Nonsmall cell lung cancer (NSCLC) was a frequent class of lung cancer, almost reaching 85% of all lung cancer cases. Although treatment advances have been significant, clinical outcomes for NSCLC patients have remained disappointing owing to pervasion. 2 More recently, a series of studies reported long noncoding RNA (lncRNA) involved in progression of NSCLC through acting as biomarker, 3 potential therapeutic target, 4 prognostic indicator. 5 Incidentally, dysregulation of oncogenes, silencing of tumor suppressor genes, activating genes to enhance growth, and metastasis of tumor were major risks resulted in sensitization of NSCLC. 6 –8
LncRNA, a class of non-protein-coding capacity with >200 bp in length, had been recognized to serve central regulators in biological processes. 9 More importantly, emerging evidence suggested that lncRNA exerted its functions in multiple cancers by improving metastasis, 10 drug resistance. 11 Small nucleolar RNA host gene 20 (SNHG20), 2183 bp in length, has been broadly reported to play a significant role in carcinogenesis and cancer progression, serving as a reliable prognostic predictor for cancer patients, such as oral squamous cell carcinoma, 12 hepatocellular, 13 NSCLC, 14 and colorectal cancer. 15
In the light of previous results, the abnormal expression of SNHG20 could regulate cell growth and metastasis in various human cancers, for instance, downregulation of SNHG20 obviously inhibited cell growth in vivo in glioblastoma cells 16 ; conversely, elevated expression of SNHG20 increased growth in esophageal squamous cell carcinoma. 17 Guan et al. demonstrated that upregulation of SNHG20 functioned as a molecular sponge to facilitate proliferation and mobility through targeting miR-495 in breast cancer, 18 besides, miR-140-5p also was a potential target gene of SNHG20 in cervical cancer. 19
Moreover, Chen et al. found that SNHG20 level was markedly associated with poor overall survival and was a new candidate for diagnosis and therapy in NSCLC. 14 However, the functional role of SNHG20 contributed to NSCLC progression was still not fully clear. In this research, the authors planned to investigate functional role and potential regulatory mechanisms of SNHG20. The authors first examined SNHG20 level with real-time quantitative polymerase chain reaction (RT-qPCR) assay. Subsequently, loss of functional experiment was employed to probe the function of SNHG20 in two different NSCLC cells. The potential target gene of SNHG20 was analyzed with dual-luciferase reporter assay. In summary, this study intended to explore function and regulatory mechanism of SNHG20 in NSCLC.
Materials and Methods
Patient specimens and cells
The cancer tissues samples and adjacent normal tissues were isolated from 31 NSCLC patients who underwent resection surgery at The First People's Hospital of Guiyang hospital. The cancer tissues samples and adjacent normal tissues were frozen in liquid nitrogen after harvesting and then stockpiled at −80°C. The study was conducted with approval and supervision from The First People's Hospital of Guiyang and conformed to the Declaration of Helsinki. Written informed consent was provided by NSCLC patients.
A549 and H1299 cells were gained from the American Type Culture Collection (Rockville, MD).16HBE were acquired from Institute of Respiratory Diseases (Guangzhou, China). All cells were maintained in Dulbecco's modified Eagle's medium (GIBCO BRL, Grand Island, NY) supplement with 10% (v/v) fetal bovine serum (FBS; InvivoGen, San Diego, CA), 100 U/mL penicillin and 100 mg/mL streptomycin in an atmosphere at 37°C with 5% CO2.
Real-time quantitative polymerase chain reaction
NSCLC cell lines and tissues were lysed using Trizol Reagent (Solarbio, Beijing, China) to isolate total RNA. To evaluate the expression levels of SNHG20, E2F transcription factor 3 (E2F3) and miR-2467-3p, Prime Script RT Reagent kit (Takara, Dalian, China) were used to reversely transcribe RNA. After that, the expression level of RNA was computed using SYBR-Green Reagent kit (Qiagen, Hilden, Germany). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or nuclear RNA U6 was used as endogenous control. The expression level of RNA was relatively quantified by using 2−ΔΔCt method.
The primers were listed as follows:
SNHG20 (F-5′-ATGGCTATAAATAGATACACG-3′; R-5′-GGTACAAACAGGGAGGGA-3′);
E2F3 (F-5′-AATATGGCGTAGTATCTCCG-3′; R-5′-CTTCCCAAACATACACCCAC-3′);
miR-2467-3p (F-5′-GCCGAGGGACAGGCACCTGA-3′; R-5′-CTCAACTGGTGTCGTGGA-3′);
GAPDH (F-5′-TCCCATCACCATCTTCCAGG-3′; R-5′-GATGACCCTTTTGGCTCCC-3′);
U6 (F-5′-AACGAGACGACGACAGAC-3′; R-5′-GCAAATTCGTGAAGCGTTCCATA-3′).
Western blot assay
Whole protein was isolated from tissues or transfected cells with protein extraction kit (KeyGen, China). The extracts incubated on ice and centrifuged 12,000 rpm for 10 min at 4°C to divide cell debris. Equal amounts of total loaded on sodium dodecyl sulfate polyacrylamide gel electrophoresis and then blotted on polyvinylidene fluoride membranes (Schleicher & Schuell, Keene, NH). The membranes were blocked with 5% milk in phosphate-buffered saline (PBS) and then incubated with antibodies. Detection of protein was conducted overnight at 4°C using anti-E2F3 (sc-56665; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), antiphosphatidylinositol 3-kinase (PI3K, sc-374534; Santa Cruz Biotechnology, Inc.), anti-p-PI3K (ab182651; Abcam, Cambridge, United Kingdom), antiprotein kinase B (AKT, sc-81434; Santa Cruz Biotechnology, Inc.), anti-p-AKT (sc-377556; Santa Cruz Biotechnology, Inc.), antimammalian target of rapamycin (mTOR, sc-517464; Santa Cruz Biotechnology, Inc.), and anti-p-mTOR (sc-293089; Santa Cruz Biotechnology, Inc.), anti-GAPDH (sc-47724; Santa Cruz Biotechnology, Inc.) antibodies. After washing three times for 10 min with PBST, the membranes were incubated with the secondary horseradish peroxidase-conjugated antibody, including goat-antirabbit IgG (R1131; Sigma, St. Louis, MO) and goat-antimouse IgG (M8645; Sigma). Finally, the protein bands were analyzed with enhanced chemiluminescence method.
Immunohistochemical staining
Immunohistochemical staining for E2F3 was performed as previously described. 20 A primary rabbit anti-E2F3 antibody (ab50917; Abcam) and a secondary antibody goat antirabbit IgG H&L (HRP) antibody (ab205718; Abcam) were used for staining.
RNA transfection and vector construction
The siRNA specifically targeting SNHG20 (si-SNHG20, 5′-GCCACUCACAAGAGUGUAUTT-3′) and the negative control (si-NC, 5′-GGATACGGAGTACTATAGC-3′), siRNA against E2F3 (si-E2F3, 5′-TTGCGTTACTTTAAGTACTAA-3′) and the control (si-NC, 5′-TTCTCCGAACG TGTCACGT-3′) were obtained from Santa Cruz Biotechnology. miR-2467-3p mimic (5′-AGCAGAGGCAGAGAGGCUCAGG-3′), and its negative control (miR-NC, 5′-AGGAUGUAUUACCAGUGAUCGG-3′), overexpression plasmid of E2F3 or SNHG20 and their negative control vector were purchased from GenePharma (Shanghai, China). The oligonucleotide and vector were transfected into A549 and H1299 cells utilizing Lipofectamine 3000 (InvivoGen).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazol-3-ium bromide assay
The transfected NSCLC cells at the density 4 × 103 cells/well were placed onto culture plates. After NSCLC cells incubation at 37°C with 5% CO2 for specific time, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazol-3-ium bromide (MTT) (Sigma) was used and reacted to cells for another 4 h. Subsequently, the dimethyl sulfoxide was applied to dissolve formazan crystals for 10 min. The optical density was detected at 490 nm for assessment cell viability of NSCLC cell clines on microplate reader (Applied Biosystems, Foster City, CA).
Apoptotic assay
The treated A549 and H1299 cells were collected and fixed in cold 75% ethanol to measure cell apoptosis. Apoptotic cells were double stained with (25 μg/mL Annexin V-FITC and 40 μg/mL propidium iodide (Invitrogen, Carlsbad, CA) away from light. The apoptotic cells were analyzed on a FACSorter flow cytometer (Applied Biosystems).
Transwell assay
The migration experiment was conducted using 8 μm pore size migration chambers (Merck Millipore, Eschborn, Germany). The treated A549 and H1299 cells were sowed into upper chamber with 200 μL serum-free medium; however, 600 μL of complete medium that contained 10% FBS was dropped into the lower chamber. After culturing, the cells on bottom surface of membranes were fixed and stained with 0.1% crystal violet. Finally, migration assay was evaluated by counting migration cell numbers in random three fields per membrane with microscopy (Applied Biosystems). Moreover, matrigel matrix (BD Science, Sparks, MD) was additional in membranes coated to perform invasion assay.
Dual-luciferase reporter assay
Starbase prediction software was employed to predictive binding site between miR-2467-3p and SNHG20 or E2F3. The fragments of SNHG20 or E2F3 3′UTR containing miR-2467-3p binding site as well as its mutant were cloned into pGL3-basic vectors (Realgene, Nanjing, China), named as WT-SNHG20, MUT-SNHG20, WT-E2F3, and MUT-E2F3. The constructed luciferase reporter and miR-2467-3p mimic/miR-NC were cotransfected NSCLC cells with Lipofectamine 3000 (InvivoGen). After 48 h, cells were harvested and lysed for luciferase activity assay at post-transfection.
Subcellular localization
The localization of SNHG20 was detected by Cytoplasmic & Nuclear RNA Purification Kit (Norgen Biotek Corp., Belmont, MA). Cells were lysed and then centrifuged. Then, the nuclear RNA and cytoplasmic RNA were incubated with anhydrous ethanol and Buffer SK, respectively. Subsequently, the nuclear RNA and cytoplasmic RNA were eluted using the spin column. Finally, the expression of SNHG20 in cytoplasmic and nucleus fractions was detected by qRT-PCR.
Statistical analysis
Differences of mRNA and protein expression levels were dissected with Student's t-test or one-way analysis of variance at right time. The significant difference was considered when p-value <0.05 with SPSS 21.0 software (IBM, Somers, NY). The data in this study were displayed as mean ± standard deviation. Pearson's correlation analysis was employed to assess correlations.
Results
SNHG20 and E2F3 were overexpressed in NSCLC tissues and cells
Considering SNHG20 has been reported as an oncogene in NSCLC, the authors first tested the SNHG20 in NSCLC tissues and normal lung tissues. As presented in Figure 1A, SNHG20 level was increased by nearly two-fold in the cancer tissues in comparison with its paired noncancer tissues. Similar change trend was obtained from Figure 1B, a remarkable enhancement of E2F3 expression was observed in the NSCLC tissues with respect to normal tissues with RT-qPCR assay. The authors further analyzed the relationship between SNHG20 and E2F3, the results implied that SNHG20 positively correlated E2F3 expression in NSCLC tissues (Fig. 1C). In addition, as expected, the relative protein expression level of E2F3 was higher in NSCLC tissues than that in the normal tissues (Fig. 1D). Immunohistochemical analysis results showed that E2F3 was highly expressed in NSCLC tissues (Fig. 1E). Consequently, the data strongly suggested that SNHG20 and E2F3 might be involved in the pathogenesis of NSCLC.
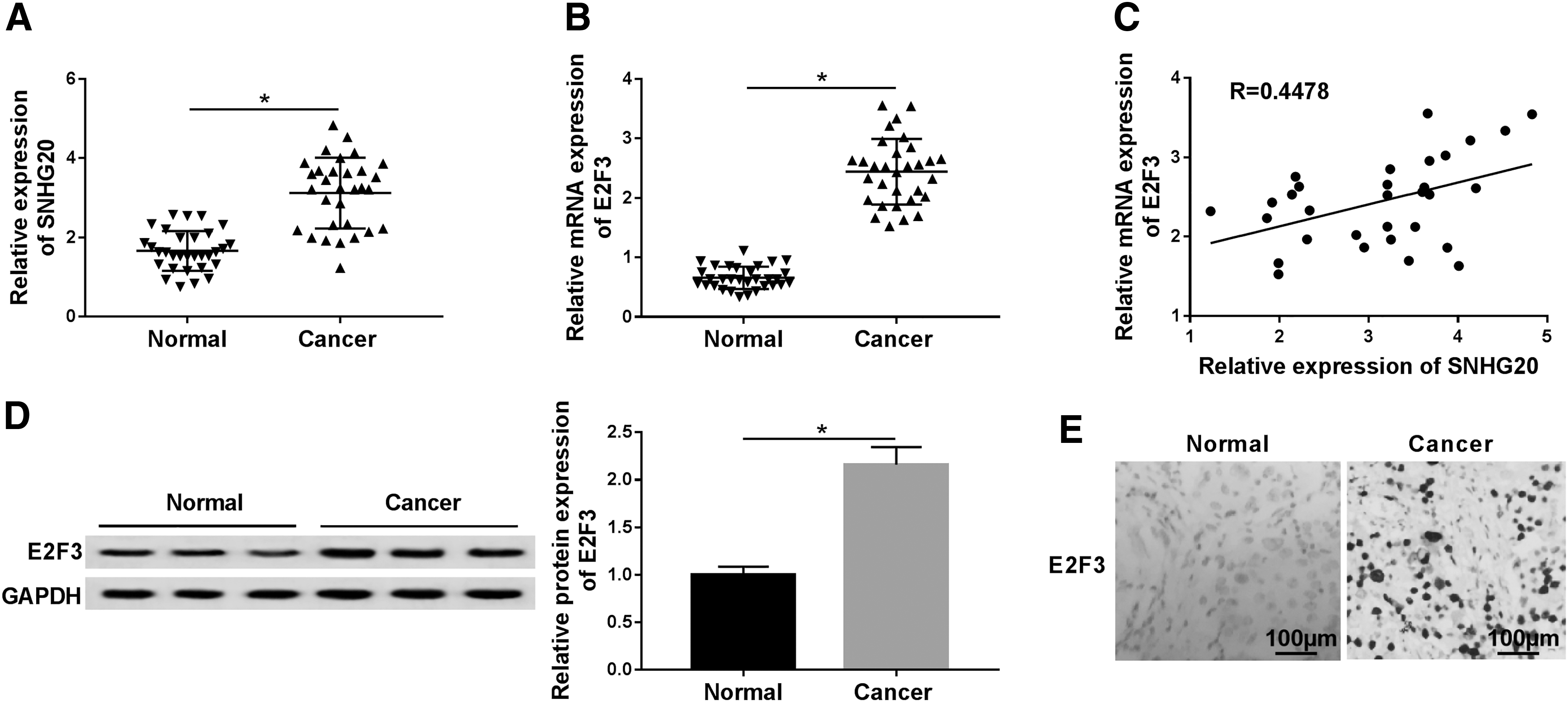
The expression levels of SNHG20 and E2F3 in NSCLC.
Silencing of SNHG20 repressed growth, mobility, and increased apoptosis of NSCLC cells
The authors first examined the expression level of SNHG20 in 16HBE and NSCLC cells. When compared with 16HBE, SNHG20 was increased in A549 and H1299 cells using RT-qPCR assay (Fig. 2A). To observe the role of SNHG20 silencing in vitro, loss-functional was applied to probe effects of SNHG20 knockdown on NSCLC. As shown in Figure 2B, RT-qPCR assay displayed that knockdown of SNHG20 by transfecting si-SNHG20 led to a notable reduction in SNHG20 expression in NSCLC cells compared with those cells transfected si-NC. Analogously, shortage of SNHG20-NSCLC cells showed a decrease in cell viability with MTT assay (Fig. 2C). Subsequently, flow cytometry was applied to analyze cell apoptosis in A549 and H1299 cells after transfection, and si-SNHG20 could also contribute to apoptosis of NSCLC cells (Fig. 2D). In addition, in comparison with si-NC group, SNHG20 silencing dramatically inhibited the abilities of migration and invasion in NSCLC cells (Fig. 2E, F). These data strongly suggested that downregulation of SNHG20 impeded NSCLC process by suppressing proliferation, mobility, and enhancing apoptosis.
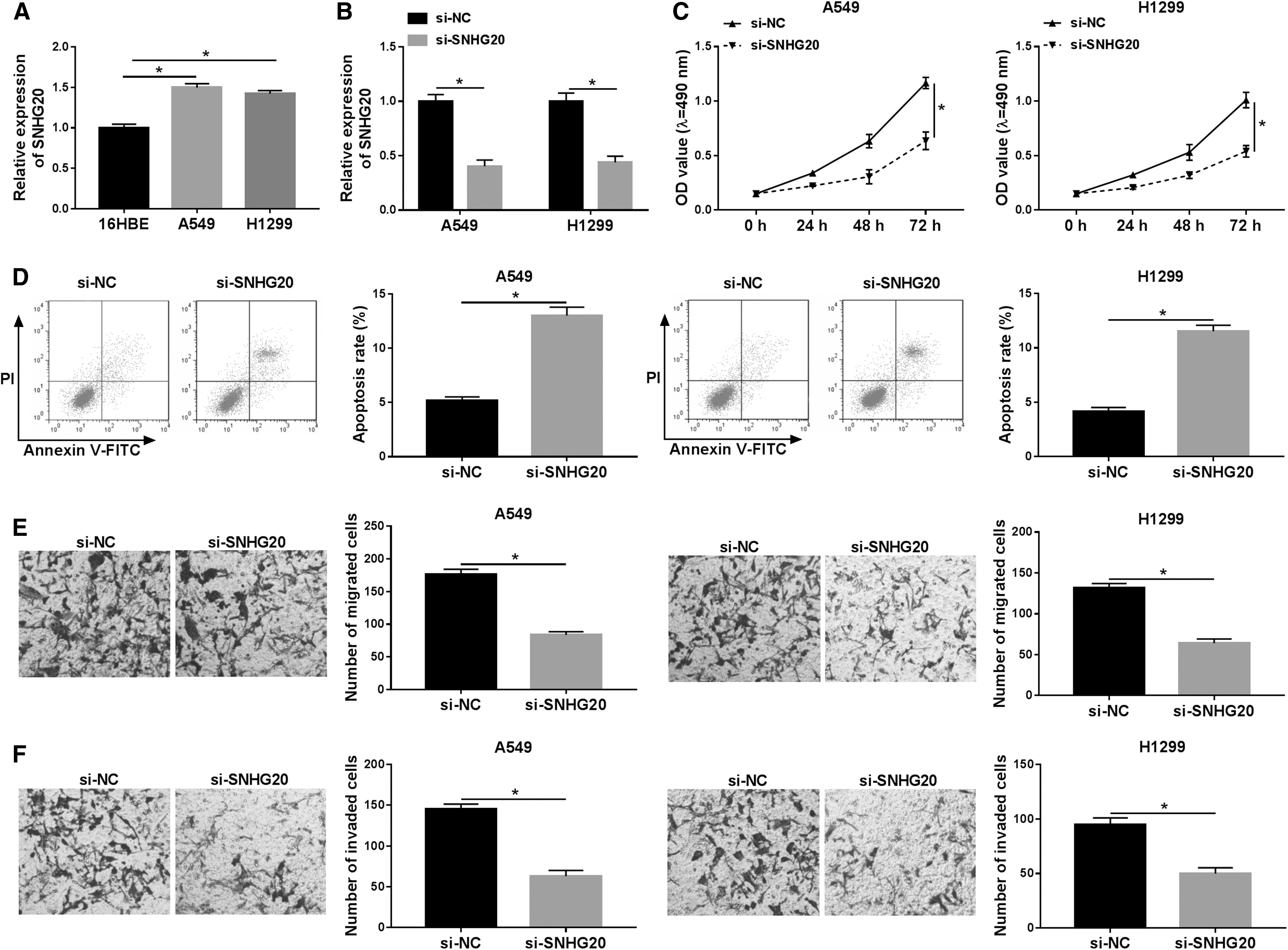
Influence of SNHG20 silencing on NSCLC cells growth, apoptosis, migration, and invasion.
E2F3 silencing suppressed tumorigenesis of NSCLC in vitro
E2F3 was overexpressed in A549 and H1299 cells than 16HBE cells (Fig. 3A, B). In addition, as presented in Figure 3C and D, the transfection of E2F3 siRNA prominently reduced the expression level of E2F3 in A549 and H1299 cells, no matter mRNA or protein. Furthermore, at 72 h post-transfection, cell viability was greatly impeded in NSCLC cells infected with si-E2F3 than that in negative group (Fig. 3E). To probe whether downregulation of E2F3 attributed to cell apoptosis, flow cytometry assay was used and indicated that A549 and H1299 cells transfected with si-E2F3 resulted in an obvious increase in cell apoptosis when in contrast to si-NC group (Fig. 3F). Finally, NSCLC cells migration and invasion assays were conducted with transwell assay. The results indicated that decreased expression of E2F3 remarkably impeded NSCLC cell mobility by reducing the number of migration and invasion (Fig. 3G, H). Collectively, the expression level of E2F3 was notably associated with capacity of proliferation, mobility, and apoptosis in NSCLC cell.

Impact of E2F3 silencing on NSCLC cells growth, apoptosis, migration, and invasion.
SNHG20 silencing impeded growth, mobility, and induced apoptosis in NSCLC cells by regulating E2F3
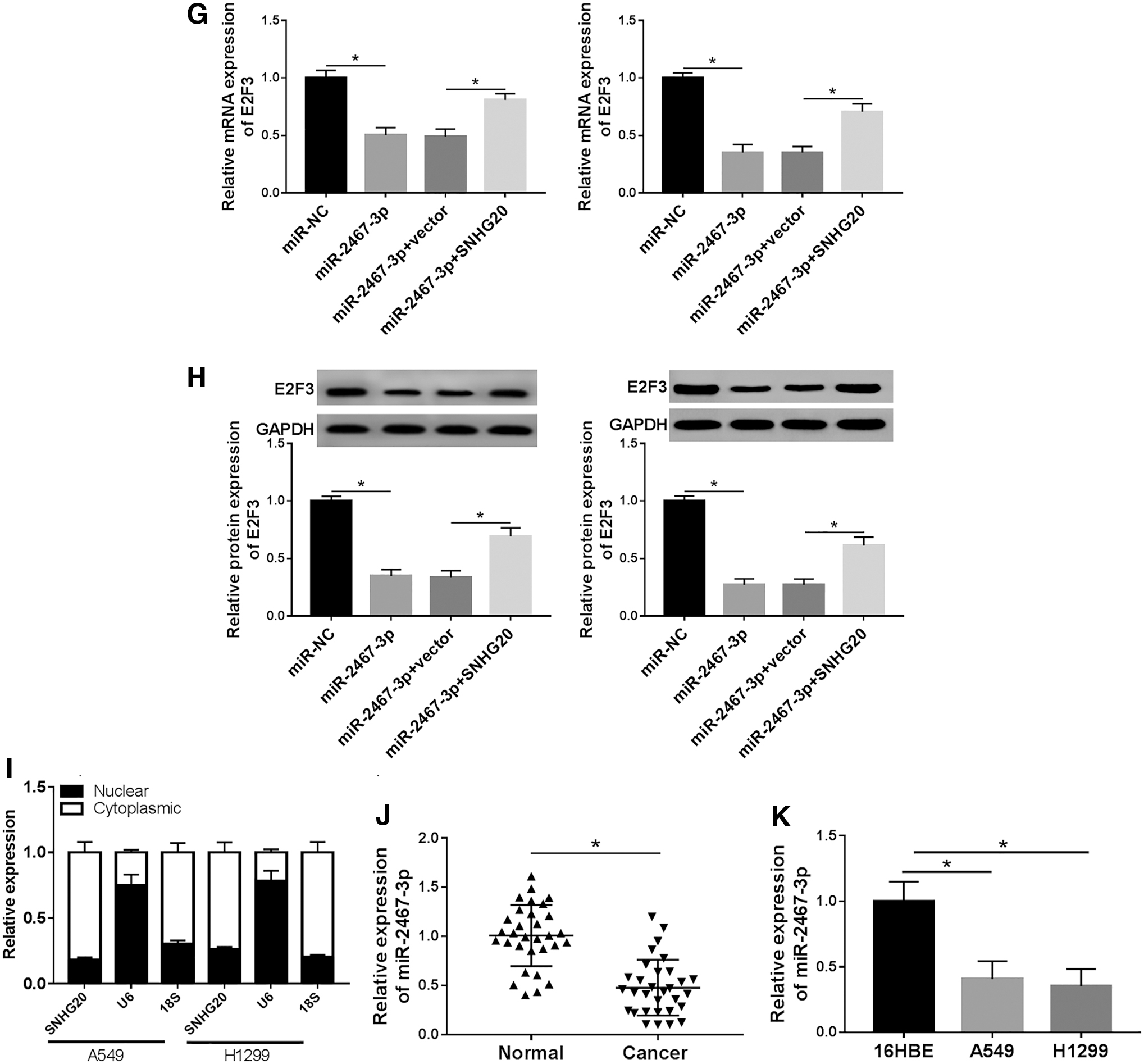
As shown in Figure 4A and B, overexpressed plasmid of E2F3 treatment prominently reinforced the E2F3 level more than twofold in A549 and H1299 cells with RT-qPCR and Western blot assays. In addition, as demonstrated by MTT assay, gain expression of E2F3 effectively mitigated inhibition of cell viability caused by silencing SNHG20 in NSCLC cells (Fig. 4C). Next, the authors also discovered the effect of E2F3 on cells apoptosis in vitro by flow cytometry assay. The results indicated that si-SNHG20 transfection significantly increased the apoptosis rate, which could be apparently restored in A549 and H1299 cells after infection with E2F3 (Fig. 4D). In addition, as shown in Figure 4E and F, overexpression of E2F3 abolished the suppression effect on cell capabilities of migration and invasion in NSCLC cells caused by knockdown SNHG20. Thus, it can be concluded that SNHG20 played a significant role in NSCLC development by regulating E2F3 expression.

Overexpression of E2F3 lightened the inhibition effects on proliferation, motility, and enhancement effects on apoptosis caused by knockdown SNHG20 in NSCLC cells.
SNHG20 and E2F3 were targets of miR-2467-3p
Starbase software was applied to predict the association between miR-2467-3p and SNHG20. Mutation sequences in the SNHG20 and normal complementary sites for miR-2467-3p are presented in Figure 5A. Dual-luciferase reporter assay implied that luciferase activity of WT-SNHG20 but not MUT-SNHG20 significantly reduced in both A549 and H1299 cells on miR-2467-3p overexpression (Fig. 5B). In addition, it is also found that E2F3 was target of miR-2467-3p. As presented in Fig. 5C, 3′UTR of E2F3 retained supposed binding sites for miR-2467-3p; moreover, the relative luciferase activity was decreased in A549 and H1299 cells cotransfected with WT-E2F3 luciferase reporter vector and miR-2467-3p, but inhibitory effect was not observed in NSCLC cells cotransfected MUT-E2F3 and miR-2467-3p mimic than control (Fig. 5D). Next, gain-functional experiment results indicated that A549 and H1299 cells infected with miR-2467-3p mimic notably augmented miR-2467-3p level compared with miR-NC group (Fig. 5E); analogously, a remarkable enhancement of SNHG20 expression was observed in NSCLC cells transfected with overexpression plasmid of SNHG20 (Fig. 5F). Eventually, the results also evidenced that elevated expression of SNHG20 could effectively repress low expression of E2F3 in A549 and H1299 cells caused by miR-2467-3p mimic (Fig. 5G, H). The data showed that SNHG20 was highly expressed in the cytoplasm of A549 cells in comparison with nuclear, indicating that SNHG20 mainly exerted its function in the cytoplasm of cells (Fig. 5I). Moreover, RT-qPCR results suggested that the expression of miR-2467-3p was lowly expressed in NSCLC tissues and cells (Fig. 5J, K). Together with the data, SNHG20 regulated NSCLC through targeting miR-2467-3p/E2F3 axis.

Prediction of miR-2467-3p target gene.
SNHG20 contributed to NSCLC through regulating PI3K/AKT/mTOR signaling pathway
To confirm the SNHG20 accelerated development of NSCLC through PI3K/AKT/mTOR pathway, the related protein levels were assessed in NSCLC cells. The data of Western blot assay suggested that p-PI3K, p-AKT, and p-mTOR was lower in SNHG20 silencing group when compared with si-NC, while overexpression of E2F3 counteracted the effects of SNHG20 silencing in A549 cells (Fig. 6A). The same results were confirmed in H1299 cells (Fig. 6B). Therefore, these results indicated that SNHG20 regulated PI3K/AKT/mTOR signaling pathway in NSCLC by increasing p-PI3K, p-AKT, and p-mTOR through affecting E2F3 expression.
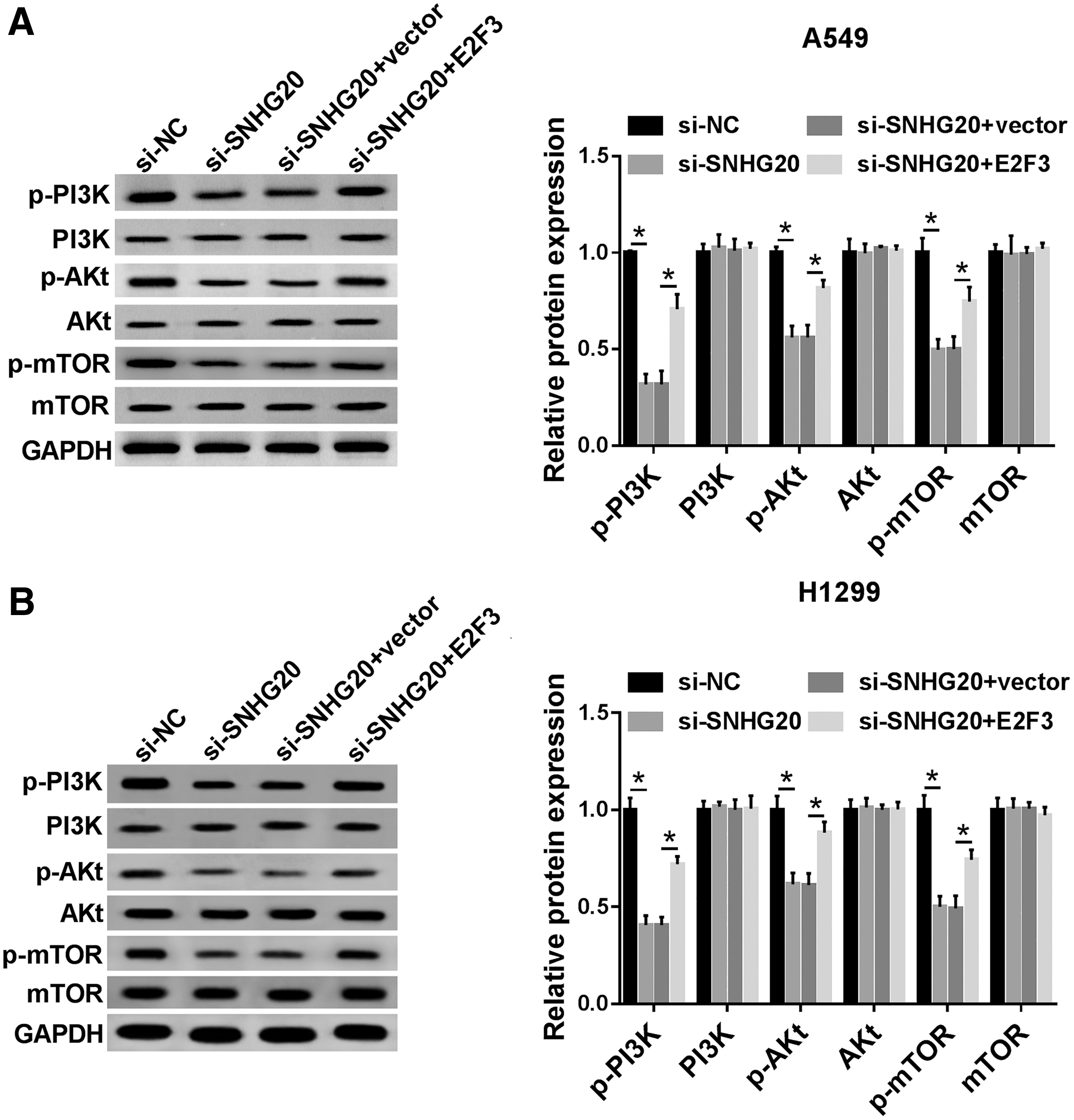
SNHG20 mediated PI3K/AKT/mTOR signaling pathway through regulating E2F3.
Discussion
In this context, it is established that SNHG20 was increased in NSCLC tissues and cells when contrasted to match negative controls. Besides, according to the Lingling et al. study, 21 mechanistic analysis implied that SNHG20 silencing impeded proliferation and motility, as well as augmented apoptosis in NSCLC cells.
Recently, mounting evidence implied that lncRNA functioned as the competitive endogenous RNA to sequester target mRNA expression. A miRNA array conducted by Xiao et al. and confirmed that hsa-miR-2467-3p was upregulated in metastatic colorectal cancer tissues with respect to nonmetastatic colorectal cancers, 22 but the role of miR-2467-3p had not been reported in NSCLC. The bioinformatics-predicted analysis and dual-luciferase reporter analysis implied that miR-2467-3p was a downstream gene of SNHG20 in NSCLC cells. Besides, rescue experiments revealed that enhancement of E2F3 significantly weakened antioncogenic function of SNHG20 silencing. Consequently, E2F3 was target of miR-2467-3p, which was verified by dual-luciferase reporter analysis.
E2F3, a member of the E2F family, was oncogenic in tumorigenesis. 23 Gao et al. pointed out E2F3 was obviously overexpressed in NSCLC tissues in comparison with paired noncancer tissues, 24 besides, E2F3 acted as new biomarkers for the prognosis of human lung carcinoma. 25,26 Previous studies have shown that miR-217 repressed growth and invasion of pancreatic cancer cell by regulating E2F3 27 ; in addition, miR-203a targeted E2F3 in gastric cancer to inhibit proliferation and colony formation. 28 Zhang et al. findings indicated E2F3 contributed to proliferation through regulating cell cycle in NSCLC. 29 This result implied that E2F3 was upregulation in NSCLC tissues and cells than controls. In contrast, E2F3 silencing resulted in a remarkable reduction of cell proliferation, motility, and improvement apoptosis.
Furthermore, a study has also uncovered that PI3K/AKT/mTOR signaling pathway was associated with NSCLC development, 30 which was confirmed in these results. The loss-functional experiment confirmed that SNHG20 may activate PI3K/AKT/mTOR by increasing phosphorylation protein levels (p-PI3K, p-AKT, and p-mTOR) in NSCLC cells. Similar conclusions were obtained from Gao et al. findings, which suggested that the PI3K/AKT/mTOR pathway was mediated by SNHG20, thereby promoting tumorigenesis of glioblastoma. 16
Conclusions
In summary, it is confirmed that SNHG20 was aberrantly expressed and functioned as oncogenic in NSCLC by enhancing E2F3 expression and activating PI3K/AKT/mTOR signaling pathway. Notably, the data first evidenced SNHG20–miR-2467-3p regulatory network in NSCLC, which was a promising and novel approach for the diagnosis of NSCLC.
Footnotes
Authors' Contributions
H.C. and Y.D. conceived the study. X.T. drafted the article. H.C. critically revised the article for important intellectual content. Y.D. facilitated all project-related tasks. All authors have read and approved the article and agreed to be accountable for all aspects of the study in ensuring that questions related to the accuracy or integrity of any part of the study are appropriately investigated and resolved.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.