
Abstract
Objective:
LncRNA IUR has been recently identified as a key regulator of Bcr-Abl-induced tumorigenesis, while its role in gastric carcinoma (GC) is unknown. This study investigated the involvement of IUR in GC.
Materials and Methods:
Gene expression levels were measured by performing quantitative polymerase chain reaction. Interactions between IUR and ROCK1 were analyzed by transfection experiments. Cell invasion and migration were analyzed by Transwell assay.
Results:
In this study, the authors showed that IUR was downregulated in GC. A follow-up study showed that low IUR expression levels predicted poor survival. In GC tissues, ROCK1 was upregulated in GC tissues and inversely correlated with IUR. In GC cells, IUR overexpression mediated the downregulation of ROCK1. ROCK1 overexpression resulted in increased GC cell invasion and migration, while IUR overexpression played an opposite role.
Conclusion:
IUR is downregulated in GC and inhibits GC cell invasion and migration by downregulating ROCK1.
Introduction
Although the incidence of gastric carcinoma (GC) ranks fourth among all types of cancers, it is the 2nd major cause of cancer-related death, owing to its rapid progression and aggressive nature. 1 The prevalence of GC varies widely around the world. GC mainly affects people in South Africa, Eastern Europe, and East Asia. 2 Every year, more than 380,000 Chinese are affected by GC and more than half of the patients will die of this disease within 5 years after diagnosis. 3 Chronic bacterium Helicobacter pylori and Epstein/Barr virus infections are closely correlated with the occurrence of GC. 4,5 However, these infections are not sufficient for the occurrence of GC, which highlights the involvement of genetic factors. 6
Rho-associated, coiled-coil-containing protein kinase 1, or ROCK1, is a type of protein serine/threonine kinase that plays pivotal roles in regulating contractile force generation. 7 In cancer biology, ROCK1 mainly affects cancer cell motility and angiogenesis. 7,8 In effect, inactivation of ROCK1 is a potential approach for cancer therapy. 9 It is known that ROCK1 expression in cancer can be regulated by certain noncoding RNAs (ncRNAs), such as long ncRNAs (lncRNAs, >200 nt). 10,11 IUR is a recently identified tumor suppressive lncRNA and can inhibit Bcr-Abl-induced tumorigenesis in Bcr-Abl-positive acute lymphoblastic leukemia. 12 It is known that both IUR and ROCK1 have interactions with STAT signaling. 12,13 Therefore, IUR and ROCK1 may have interactions. They aim to investigate the interaction between IUR and ROCK1 in GC.
Materials and Methods
GC patients
This study selected 60 GC patients (gender: 34 males and 26 females; age range: 29–68 years; mean age: 49.3 ± 6.4 years) from the 122 cases of GC patients admitted to Huanggang Central Hospital between March 2010 and January 2014. Inclusion criteria were: (1) all GC patients were newly diagnosed; (2) no initiated therapies were observed; and (3) patients willing to complete follow-up. Exclusion criteria were: (1) patients suffered from multiple clinical disorders; (2) patients with recurrent GC; and (3) patients with initiated therapies. Clinical staging was performed according to the standard established by AJCC. There were 12 (stage II), 22 (stage III), and 26 (stage IV) cases, respectively. Before the admission of patients, the aforementioned hospital's Ethics Committee approved this study. All these 60 GC patients signed informed consent.
GC tissues and cells
Biopsy was performed on all 60 GC patients before the initiation of therapies to collect nontumor gastric tissues and GC tissues. Histopathological examinations were performed to test all tissue samples, and all tissues samples were correct: (1) nontumor tissues contained no or less than 1% cancer cells; (2) cancer tissues contained more than 90% cancer cells.
Two human GC cell lines, SNU-16 and N87 (ATCC), were used. A mixture of 10% FBS and 90% RPMI-1640 medium was used for cell culture. Cell culture conditions were 5% CO2, 37°C, and 95% humidity.
Follow-up
All 60 GC patients were monitored for 5 years. All patients were visited on a monthly manner through telephone. Patients who died of other diseases and the ones who failed to complete the follow-up were excluded.
Transient transfections
The vectors expressing IUR and ROCK1 were constructed by RIBOBIO (Guangzhou, China) using pcDNA3.1 vector. Transient transfections were performed using Lipofectamine 2000 reagent (Invitrogen) to transfect 10 nM vectors (empty vector as negative control, NC) into 106 cells. Control (C) cells were untransfected. The following experiments were performed at 24 h post-transfections.
Quantitative polymerase chain reaction
Total RNAs in tissues and SNU-16 and N87 cells were isolated using TRIzol (Invitrogen). All RNA samples were quantified using NanoDrop™ 2000 Spectrophotometer (Thermo Fisher Scientific). Following DNase I digestion, RNA samples were used as template to synthesize cDNAs through reverse transcription using the TruScript Reverse Transcriptase Kit (NORGEN). To detect the expression of IUR and ROCK1 mRNA, SYBR Green Master Mix (Bio-Rad) was used to perform quantitative polymerase chain reaction (qPCR) with GAPDH as endogenous control. Three replicate qPCRs were set for each experiment. All qPCR data were normalized using 2−ΔΔCT method.
Methylation-specific PCR
SNU-16 and N87 cells were subjected to DNA extraction using the Genomic DNA Extraction Kit (ab156900) from Abcam. DNA samples were converted using the DNA Methylation-GoldTM Kit (ZYMO RESEARCH). The Taq DNA Polymerase Kit (NEB) was then used to perform methylation-specific PCR (MSP) to analyze the methylation of ROCK1 gene.
Transwell assay
SNU-16 and N87 cells were harvested at 24 h post-transfections. After cell counting, RPMI-1640 medium was used to resuspend cells with a ratio of 1 mL per 4 × 104 cells. A 96-well Transwell plate was used to cultivate cells (0.1 mL cell suspension per well in the upper chamber) under conditions of 5% CO2, 37°C, and 95% humidity. The lower chamber contained RPMI-1640 medium (20% FBS). Membranes were coated with Matrigel (356234; Millipore) for 6 h at 37°C before invasion assay. Cell culture was performed for 12 h, following membrane staining for 15 min at room temperature using 1% Crystal Violet (Sigma-Aldrich). Cells were observed and counted under a light microscope.
Western-blot
RIPA solution (Sangon) was used to isolate total proteins. After protein denaturation in boiling water for 5 min, protein samples were separated through electrophoresis using SDS-PAGE gel (10%). The proteins were transferred onto PVDF membranes and blocked in FBS containing 5% nonfat milk for 1 h. PVDF membranes were first blotted with ROCK1 (ab156284, 1:1100; Abcam) and GAPDH (ab9485, 1: 1100; Abcam) primary rabbit antibodies at 4°C overnight, followed by blotting with goat anti-Rabbit IgG-HRP secondary antibody(1:1100, MBS435036; MyBioSource). Signals were developed by dropping ECL (Sigma-Aldrich) onto PVDF membranes. Gray values were normalized using ImageJ v.146 software.
Statistical analysis
Paired t test was used to compare GC and nontumor tissues. One-way ANOVA and Tukey's test were used to compare multiple groups. Linear regression was used to analyze correlations. Patients were divided into high and low IUR level groups according to the median value of the expression levels of IUR in GC tissues. Survival curves were plotted and compared by log-rank test. p < 0.05 was statistically significant.
Results
IUR was downregulated in GC and low expression of IUR predicted poor survival
Expression levels of IUR in both GC and nontumor tissues of the 60 GC patients were measured by performing qPCR. It was observed that levels of IUR expression were significantly lower in GC tissues compared with nontumor tissues (Fig. 1A, p < 0.05). Survival curves were plotted for both high and low IUR level groups and compared. It can be observed that the 5-year overall survival rate of patients in low IUR group was significantly lower compared with patients in the high IUR group (Fig. 1B).
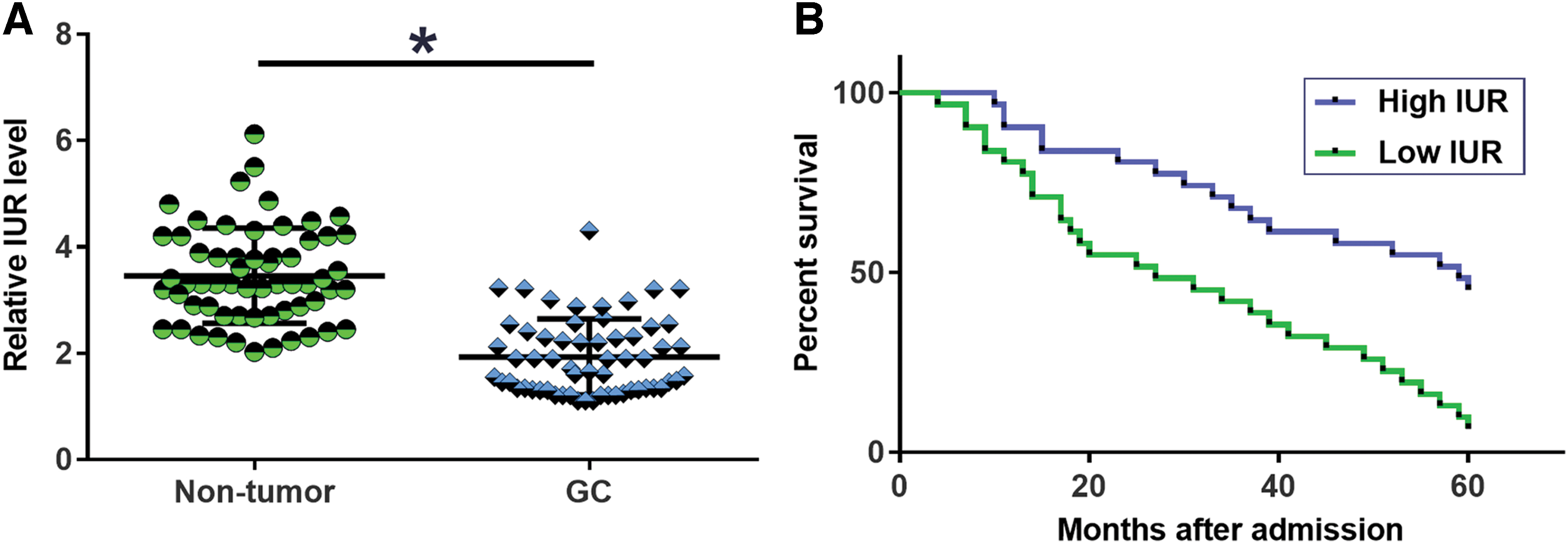
Downregulation of IUR in GC predicted poor survival. Expression levels of IUR in both GC and nontumor tissues of the 60 GC patients were measured by performing qPCR and data were compared by paired t test
ROCK1 was inversely correlated with IUR in GC
Expression levels of ROCK1 mRNA in two types of tissues were also measured by performing qPCR. Levels of ROCK1 mRNA expression were significantly higher in GC tissues in comparison to nontumor tissues (Fig. 2A, p < 0.05). Correlation analysis showed that ROCK1 mRNA and IUR were significantly, inversely correlated across GC tissues (Fig. 2B). However, the correlation between ROCK1 mRNA and IUR was not significant in nontumor tissues (Fig. 2C).
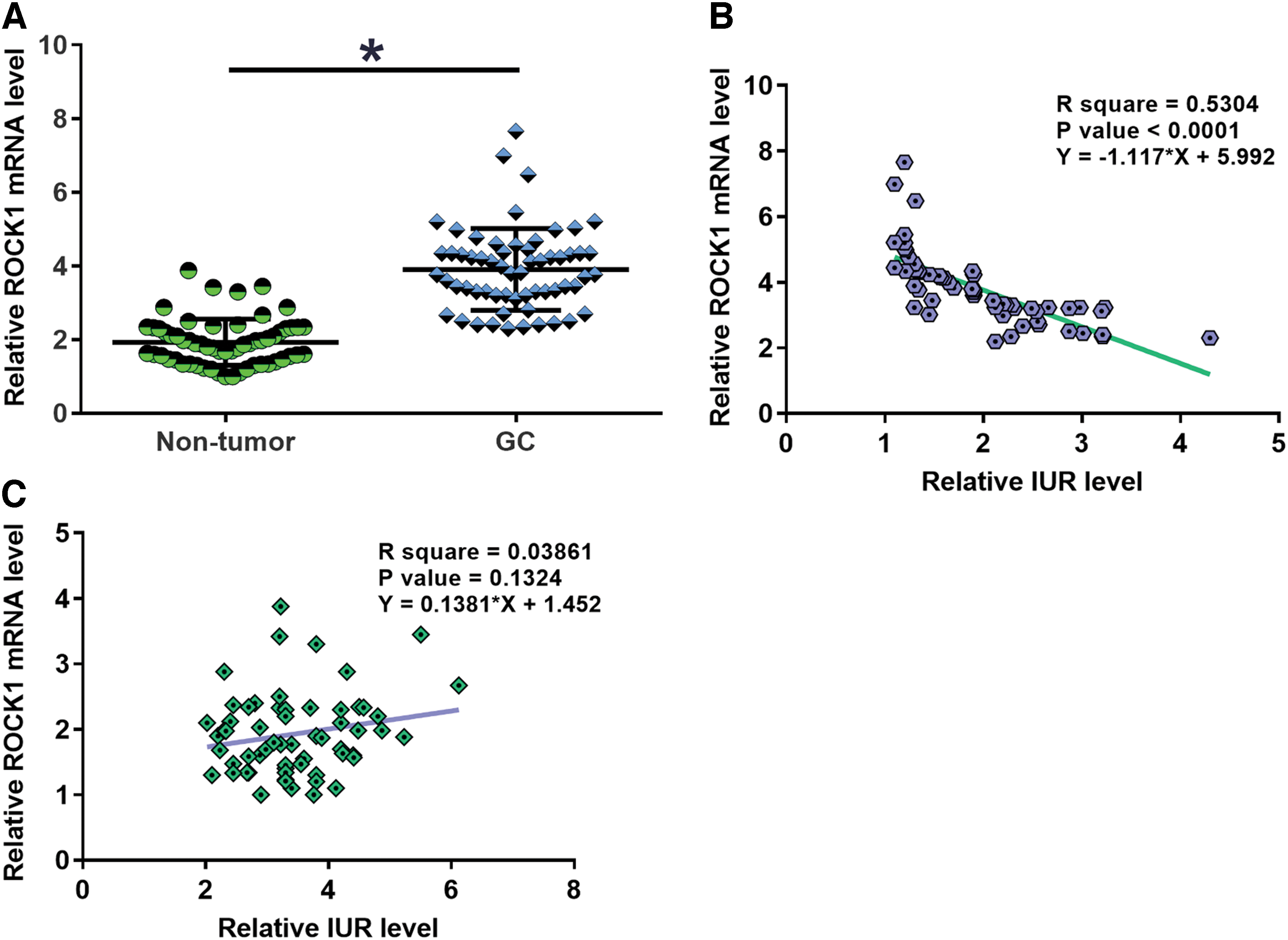
ROCK1 was inversely correlated with IUR in GC. Levels of ROCK1 mRNA expression were also measured by performing qPCR and compared by paired t test between GC and nontumor tissues
IUR overexpression downregulated ROCK1 in GC cells
SNU-16 and N87 cells were transfected with pcDNA3.1 vectors expressing IUR and ROCK1. Overexpression of IUR and ROCK1 was confirmed at 24 h post-transfection (Fig. 3A, p < 0.05). Moreover, compared with two controls, IUR overexpression was followed by ROCK1 downregulation (Fig. 3B, p < 0.05), while no alterations in IUR expression were observed in cells with ROCK1 overexpression (Fig. 3C).
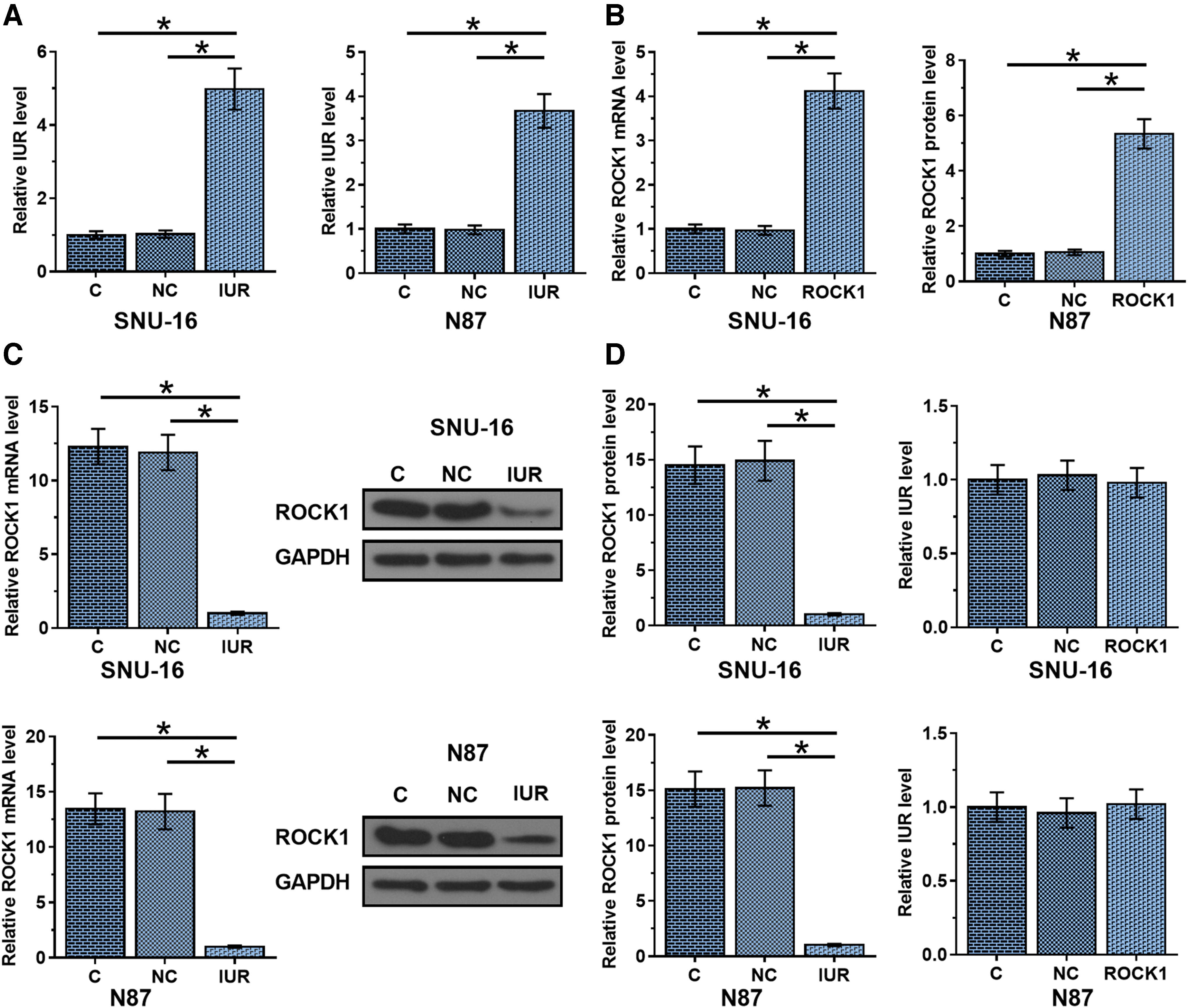
IUR overexpression was accompanied by ROCK1 downregulation in GC cells, SNU-16 and N87 cells were transfected with pcDNA3.1 vectors expressing IUR and ROCK1. Overexpression of IUR and ROCK1 was confirmed at 24 h post-transfection
IUR promoted the invasion and migration of GC cells
Transwell assay analysis showed that ROCK1 overexpression resulted in increased GC cell invasion (Fig. 4A, p < 0.05) and migration (Fig. 4B, p < 0.05) rates, while IUR overexpression resulted in decreased rates of GC cell invasion (Fig. 4A, p < 0.05) and migration (Fig. 4B, p < 0.05). Moreover, ROCK1 overexpression inhibited the role of IUR overexpression (p < 0.05).
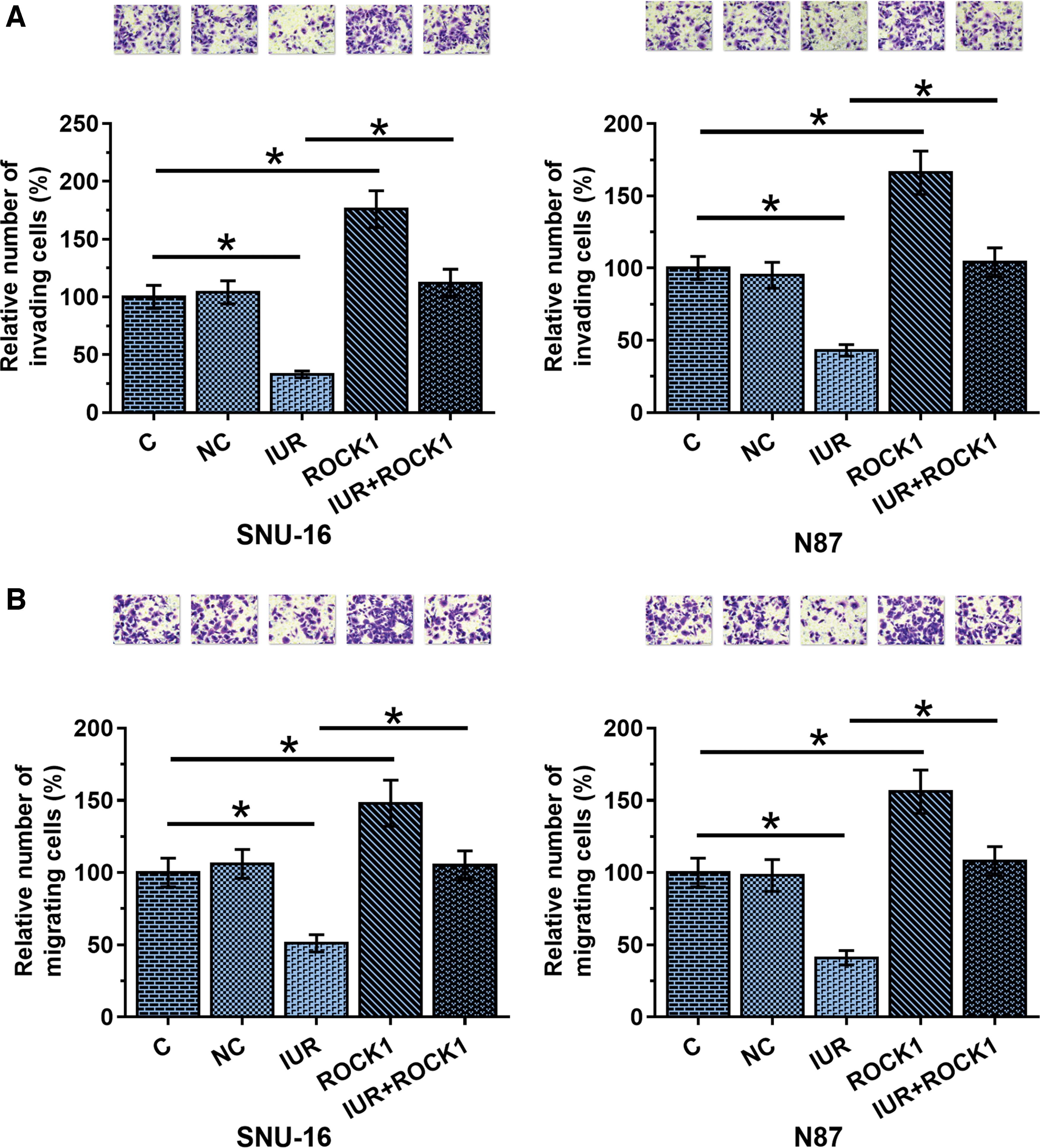
IUR promoted the invasion and migration of GC cells. The effects of IUR and ROCK1 overexpression on cell invasion and migration abilities were analyzed by performing Transwell invasion and migration assay. Experiments were repeated three times and mean values were presented, *p < 0.05. Color images are available online.
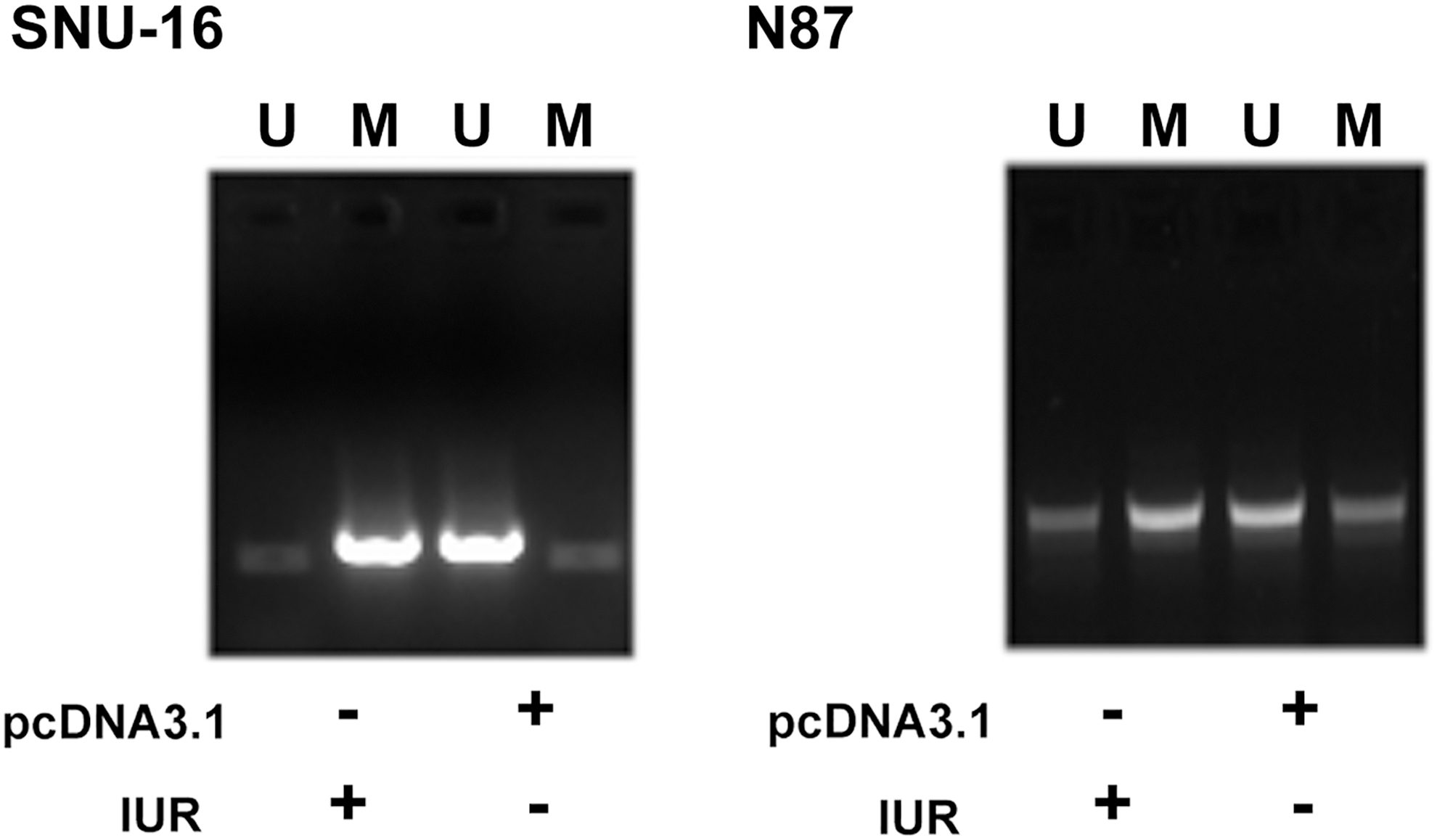
IUR overexpression increased the methylation of ROCK1 in GC cells. MSP was performed to analyze the effects of IUR overexpression on the methylation of ROCK1 gene. Experiments were repeated three times and representative images were presented. U, unmethylation; M, methylation.
IUR overexpression increased the methylation of ROCK1 in GC cells
MSP was performed to analyze the effects of IUR overexpression on the methylation of ROCK1 gene. Compared with cells transfected with empty pcDNA3.1 vector, cells transfected with IUR expression vector showed increased methylation of ROCK1.
Discussion
The functions of IUR in GC have been investigated in this study. The authors found that IUR was downregulated in GC. In addition, IUR may downregulate ROCK1 in GC to inhibit the invasion and migration of GC cells.
Although treatment outcomes of GC have been significantly improved during the past several decades, overall survival of this disease is still poor. 14 The poor survival of GC patients is mainly because of the low early diagnostic rate. 15 However, early diagnosis of GC is technically challengeable, and effective and sensitive biomarkers are lacking. 15 As an alternative approach, studies have been carried out to develop prognostic markers for GC, and the accurate prognosis assignment can help the determination of therapeutic approaches and care programs. 16,17 However, the accuracy of most prognostic markers for GC identified so far remains low. In a recent study, Wang el al. reported IUR as a tumor-suppressive lncRNA in Bcr-Abl-positive acute lymphoblastic leukemia. 12 This study is the first to report downregulation of IUR in GC. Moreover, low IUR levels in GC tissues are closely correlated with the poor survival of GC patients. However, the accuracy of IUR in the prognosis of GC remains to be further analyzed.
ROCK1 plays a critical role in the metastasis of GC tumor. 18 Consistently, they observed the upregulation of ROCK1 in GC and the increased invasion and migration rate of GC cells after ROCK1 overexpression. Their data further confirmed the oncogenic role of ROCK1 in GC. In this study, they also showed that IUR could downregulate ROCK1 at both mRNA and protein levels in GC cells, thereby inhibiting GC cell migration and invasion. They speculate the interaction between IUR and ROCK1 is indirect due to lack of significant correlation between ROCK1 and IUR in nontumor tissues. Both IUR and ROCK1 have interactions with STAT signaling, 12,13 and STAT signaling is activated in GC. Therefore, the activation of STAT signaling may mediate the interaction between IUR and ROCK1 in GC. In addition, their data also showed that IUR overexpression in GC cells resulted in the increased methylation of ROCK1 gene. Therefore, IUR may also downregulate ROCK1 through methylation.
In conclusion, IUR is downregulated in GC and may downregulate ROCK1 to suppress invasion and migration of GC cells.
Footnotes
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
No funding was received for this article.