
Abstract
Background:
Hepatocellular carcinoma (HCC) is one of the most common malignancies with high invasion and metastasis capacities. Circular RNAs (circRNAs) were evidenced to take part in the progression of multifarious cancers, including HCC. However, the role of circ_0091579 in HCC progression has not been fully described. This study aimed to explore the function of circ_0091579 and its potential regulatory mechanism in the progression of HCC.
Materials and Methods:
The expression of circ_0091579, microRNA-490-5p (miR-490-5p), and cancer susceptibility candidate 3 (CASC3) in HCC tissues and cells was detected by quantitative real-time polymerase chain reaction. The circular characteristic and stability of circ_0091579 were verified by RNase R digestion and actinomycin D reaction assays. Cell proliferation, migration, and invasion were determined by methyl thiazolyl tetrazolium assay and Transwell assay, respectively. The level of glycolysis was evaluated by glucose consumption and lactate production. The levels of proteins were examined by Western blot. The interaction between miR-490-5p and circ_0091579 or CASC3 was certified by Dual-luciferase reporter assay.
Results:
circ_0091579 and CASC3 were upregulated, while miR-490-5p was downregulated in HCC tissues and cells. Silencing of either circ_0091579 or CASC3 suppressed cell proliferation, migration, invasion, and glycolysis in HCC cells. Moreover, miR-490-5p was verified to directly bind to circ_0091579 and CASC3. Circ_0091579 upregulated CASC3 by sponging miR-490-5p in HCC cells to promote cell proliferation, invasion, and migration.
Conclusion:
circ_0091579 promoted cell proliferation, migration, invasion, and glycolysis partially through miR-490-5p/CASC3 axis in HCC cells.
Introduction
Liver cancer is a leading high mortality cancer (in men and women) because of the problems of early detection, as well as the limited therapeutic options in later pathogenesis. 1 Hepatocellular carcinoma (HCC) is a common malignancy, which belongs to liver cancer. 2 Although the surgical, medical, and locoregional therapies have achieved advancement, HCC still has a poor prognosis. 3,4 However, the specific molecular mechanism of the occurrence and development of HCC has not been fully elucidated. Therefore, it is particularly critical to explore novel biomarkers for HCC treatment.
Circular RNAs (circRNAs) are a kind of noncoding RNAs with covalently closed continuous loops without structure of 5′ or 3′ polarities, which are more stable than linear RNAs. 5 –7 Usually, circRNAs regulated transcription and translation by acting as microRNA (miRNA) and RNA binding protein sponges or directly binding to the mRNA translation start site. 8,9 Mounting evidence demonstrated that circRNAs performed crucial parts in the progression of many diseases, including cancers, 10,11 such as colorectal cancer 12 and colon cancer. 13 Similarly, previous studies demonstrated that some circRNAs were abnormally expressed in HCC, and the imbalance of these circRNAs was relevant to the tumorigenesis of HCC. 14,15
Glypican-3 (GPC3) was a gene that was regulated by circ_0091579 in circBase, which is a database for circRNAs. Studies demonstrated that GPC3 was obviously elevated and participated in cell invasion in HCC. 16,17 Meanwhile, a previous study found that the level of circ_0091579 was significantly raised in HCC tissues. 18 However, the biological function and specific mechanisms of circ_0091579 in HCC progression are still blurry.
miRNAs are small noncoding RNA molecules with 21–25 nucleotides, and they could reduce the stability of mRNAs and mediate the tumorigenesis. 19,20 Accumulating evidence revealed that some miRNAs were abnormally expressed in HCC tissues and cells and affected the development of HCC, thus acting as biomarkers for HCC diagnosis and prognosis. 21,22
In recent years, microRNA-490-5p (miR-490-5p) has been testified to be abnormally expressed and could function as antioncogene in multifarious cancers, such as bladder cancer, 23 pharyngolaryngeal cancer, 24 and glioma. 25 Moreover, miR-490-5p was downregulated in HCC and could inhibit cell proliferation, migration, and invasion by targeting specific genes. 26 –28 Hence, it was speculated that miR-490-5p served as an antitumor factor in HCC progression. However, the role of miR-490-3p in HCC was controversial. Zhang et al. reported that miR-490-3p was upregulated in HCC and promoted the development of HCC. 29 Another study suggested that miR-490-3p functioned as a tumor suppressor in HCC. 30
Cancer susceptibility candidate 3 (CASC3), also called Metastatic lymph node 51 (MLN51), is a component of exon junction complex. 31 CASC3 has been testified to take part in some diseases, including breast cancer 32 and rheumatoid arthritis. 33 Furthermore, knockdown of CASC3 could suppress the proliferation of HCC cells. 34 Therefore, it is very meaningful to explore the effects of CASC3 on HCC.
In this study, the levels of circ_0091579 in HCC tissues and cells were first determined. Moreover, gain- and loss-of-function experiments were used to verify the role of circ_0091579 in the progression of HCC and underlying regulatory mechanisms among circ_0091579, miR-490-5p, and CASC3.
Materials and Methods
Clinical samples
HCC serum samples (n = 51), as well as 51 pairs of HCC tissues and corresponding adjacent normal tissues from HCC patients, and healthy serum samples (n = 47) from healthy volunteers were collected at Luhe Hospital Affiliated to Capital Medical University. The patients involved in this research had not received anticancer therapies preoperatively. All the patients and healthy volunteers signed written informed consent. Meanwhile, this research had gained an approval from the Ethics Committee of Luhe Hospital Affiliated to Capital Medical University. The clinical features of the HCC patients are provided in Table 1.
The Correlation of circ_0091579 Expression in the Serum of Hepatocellular Carcinoma Patients with Clinical Features of Hepatocellular Carcinoma Patients
Cell culture
The normal human liver epithelial cell line THLE3 and HCC cell line Hep3B were obtained from American Type Culture Collection (ATCC, Manassas, VA) and cultivated in Minimum Essential Medium (Gibco, Grand Island, NY). HCC cell lines (HCCLM3, MHCC97L, and Huh-7) were acquired from Procell Life Science and Technology Co., Ltd. (Wuhan, China). HCCLM3 and Huh-7 cells were cultivated in Dulbecco's modified Eagle's medium (high glucose) (DMEM; Gibco). MHCC97L cells were grown in RPMI-1640 medium (Invitrogen, Carlsbad, CA). Above medium with 10% fetal bovine serum (FBS; Gibco) was utilized to culture the indicated cells at 37°C in an incubator with 5% CO2.
Quantitative real-time polymerase chain reaction
TRIzol reagent (Invitrogen) was used to extract total RNA from tissues and cells. High Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Waltham, MA) was used to synthesize first strand of complementary DNA (cDNA) for circ_0091579 and CASC3 from extracted RNA, while miRNA First Strand cDNA Synthesis Kit (Sangon Biotech Co., Ltd., Shanghai, China) was applied for cDNA synthesis of miR-490-5p. Quantitative real-time polymerase chain reaction (qRT-PCR) was proceeded using 2 × SYBR Green Abstract PCR Mix (Sangon Biotech Co., Ltd.) on a 7300 PCR System (ABI, Carlsbad, CA) based on the user guide. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) or U6 acted as the internal controls for normalization of circ_0091579 and CASC3 levels or miR-490-5p level, respectively. Relative transcriptional folds were calculated using 2−ΔΔCt method.
The primers were acquired from Sangon Biotech Co., Ltd. and listed as follows: circ_0091579: 5′-TGAGCCAGTGGTCAGTCAAA (forward) and 5′-GTGGAGTCAGGCTTGGGTAG (reverse); miR-490-5p: 5′-GCTGGTTTGGAAAGTTCATTG-3′ (forward) and R: 5′-GAAGTGTCCGCTGCTTGT-3′ (reverse); CASC3: 5′-CCACCTGAGGTTGTAAGCAGG (forward) and 5′-GTTCTCACCTCTGCTGCTTTTTA (reverse); GAPDH: 5′-AGAAGGCTGGGGCTCATTTG-3′ (forward) and 5′-AGGGGCCATCCACAGTCTTC-3′ (reverse); U6: 5′-AGAGCCTGTG GTGTCCG-3′ (forward) and 5′-CATCTTCAAAGCACTTCCCT-3′.
Cell transfection
Small interfering RNA (siRNA) against circ_0091579 (si-circ_0091579), siRNA against CASC3 (si-CASC3) and the matched negative control (si-NC), the overexpression plasmids pcDNA-circ_0091579 (circ_0091579), and the negative control (vector) were acquired from Guangzhou FulenGen Co., Ltd. (Guangzhou, China). MiR-490-5p mimics (miR-490-5p) and the matched negative control (miR-NC), miR-490-5p inhibitor (anti-miR-490-5p), and the negative control (anti-NC) were acquired from RiboBio Co., Ltd. (Guangzhou, China). HCCLM3 and Huh-7 cells were transfected with the above plasmids or oligonucleotides using Lipofectamine 2000 reagent (Invitrogen) referring to user guide. After transfection, the cells were processed for the subsequent experiments.
RNase R digestion and actinomycin D reaction assays
The circular characteristic of circ_0091579 was testified using RNase R (Applied Biological Materials, Inc., Vancouver, Canada). In brief, total RNA of circ_0091579 extracted from HCCLM3 cells and its linear counterpart mGPC3 was treated with RNase R (100 μg/mL) at 37°C for 20 min. Then, the relative RNA levels were examined by qRT-PCR analysis. To detect the stability of circ_0091579, extracted RNA was treated with transcription inhibitor actinomycin D (2 mg/mL; Sigma-Aldrich, St. Louis, MO) for 6, 12, 18, and 24 h, followed by qRT-PCR analysis.
Subcellular fractionation location
Cytoplasmic and nuclear RNA fractions of HCCLM3 cells were extracted using Cytoplasmic and Nuclear RNA Purification Kit (Norgen Biotek, Thorold, ON, Canada) following the manufacturer's protocol. Afterward, the expression proportions of cytoplasmic circ_0091579 to nucleus fractions were evaluated by qRT-PCR. GAPDH and U6 served as the cytoplasm control and nucleus control, respectively.
Methyl thiazolyl tetrazolium assay
The methyl thiazolyl tetrazolium (MTT) assay was carried out to determine cell proliferation of HCCLM3 and Huh-7 cells. The cells of different groups were seeded into 96-well plates and then transfected with assigned plasmids. Subsequently, transfected cells were further cultured for 0, 24, 48, and 72 h, severally. Next, each well was added with 15 μL MTT (Sangon Biotech Co., Ltd.), and cells were cultivated for another 4 h at 37°C to produce formazan product. Then, 150 μL dimethyl sulfoxide (Solarbio, Beijing, China) was added to dissolve the formazan product. Finally, the proliferation was determined using a microplate reader (Bio-Rad, Hercules, CA) by detecting OD values at 490 nm wavelength.
Migration and invasion assays
The migration and invasion capacities of HCCLM3 and Huh-7 cells were determined by Transwell chamber (8 μm pore; BD Biosciences, San Jose, CA). For migration assay, transfected HCCLM3 and Huh-7 cells were planted into the upper chamber, with the lower chamber adding with DMEM supplemented with 10% FBS. After incubation for 36 h, the migrated cells were dealt with 4% paraformaldehyde and then dyed with 0.5% crystal violet (Beyotime Biotechnology, Shanghai, China), followed by counted under an inverted microscope. For invasion assay, the processes were similar except for the upper chamber precoated with Matrigel (BD Biosciences).
Evaluation of glycolysis
The glucose consumption and lactate production were determined to evaluate the levels of glycolysis. After transfection for 48 h, the supernatants of cell medium were collected, and the glucose and lactic acid levels in the supernatants were detected using a Glucose Detection Kit (Zhongsheng Biotech Co. Ltd., Beijing, China) and a Lactic Acid Detection Kit (Zhongsheng Biotech Co. Ltd.) following the user guide.
Western blot assay
Total protein was isolated from HCC tissues and cells using RIPA Lysis Buffer (Thermo Fisher Scientific). The protein was quantified by a BCA Protein Assay Kit (Sangon Biotech Co., Ltd.). Then, protein was isolated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and later transferred to polyvinylidene fluoride membrane (Sangon Biotech Co. Ltd.). Next, the membrane was blocked in 5% nonfat milk powder for 1 h at room temperature. After complete blocking, the membrane was rinsed twice with Tris-buffered saline and Tween 20 (TBST) and cultured with the rabbit primary antibodies in a shaker at 4°C for 24 h.
Subsequently, the membrane was incubated with horseradish peroxidase-conjugated anti-rabbit secondary antibody (ab6721, 1:5000; Abcam, Cambridge, MA) for 2 h at room temperature after washing thrice with TBST. Finally, protein bands were detected by ECL Detection Kit (Beyotime Biotechnology), and the signal intensity of bands was determined by ImageJ software normalized to GAPDH. The primary antibodies were listed as follows: Ki-67 antibody (orb378955) (1:1000; Biorbyt, Cambridge, United Kingdom), matrix metalloproteinases-9 (MMP-9) antibody (orb214260, 1:1000; Biorbyt), hexokinase 2 (HK2) antibody (orb48965, 1:1000; Biorbyt), GAPDH antibody (orb38655, 1:1000; Biorbyt), and CASC3 antibody (orb37689, 1:1000; Biorbyt).
Dual-luciferase reporter assay
The binding sequences between miR-490-5p and circ_0091579 or CASC3 were predicted by Circular RNA Interactome database (
Statistical analysis
Data were acquired from more than three replicates and exhibited as mean ± standard deviation. Statistical analysis was conducted with the Student's t-test and one-way analysis of variance using SPSS 21.0. p < 0.05 was designated to be statistically significant.
Results
circ_0091579 was upregulated in HCC serums, tissues, and cell lines
To explore the role of circ_0091579 in HCC, the expression level of circ_0091579 in HCC serum and tissues was first detected using qRT-PCR. As a result, the expression of circ_0091579 in HCC serum samples (n = 51) was visibly raised relative to healthy serum samples (n = 47) (Fig. 1A). According to the mean of the circ_0091579 expression in HCC serum samples, 51 patients were divided into high circ_0091579 expression group and low circ_0091579 expression group, and the data showed that high circ_0091579 expression was associated with poor prognosis (Table 1). Receiver operating characteristic (ROC) curve of sensitivity and specificity was generated for circ_0091579 level to evaluate its potential diagnostic value. The result showed that the area under the ROC curve was 0.771 (Fig. 1B).
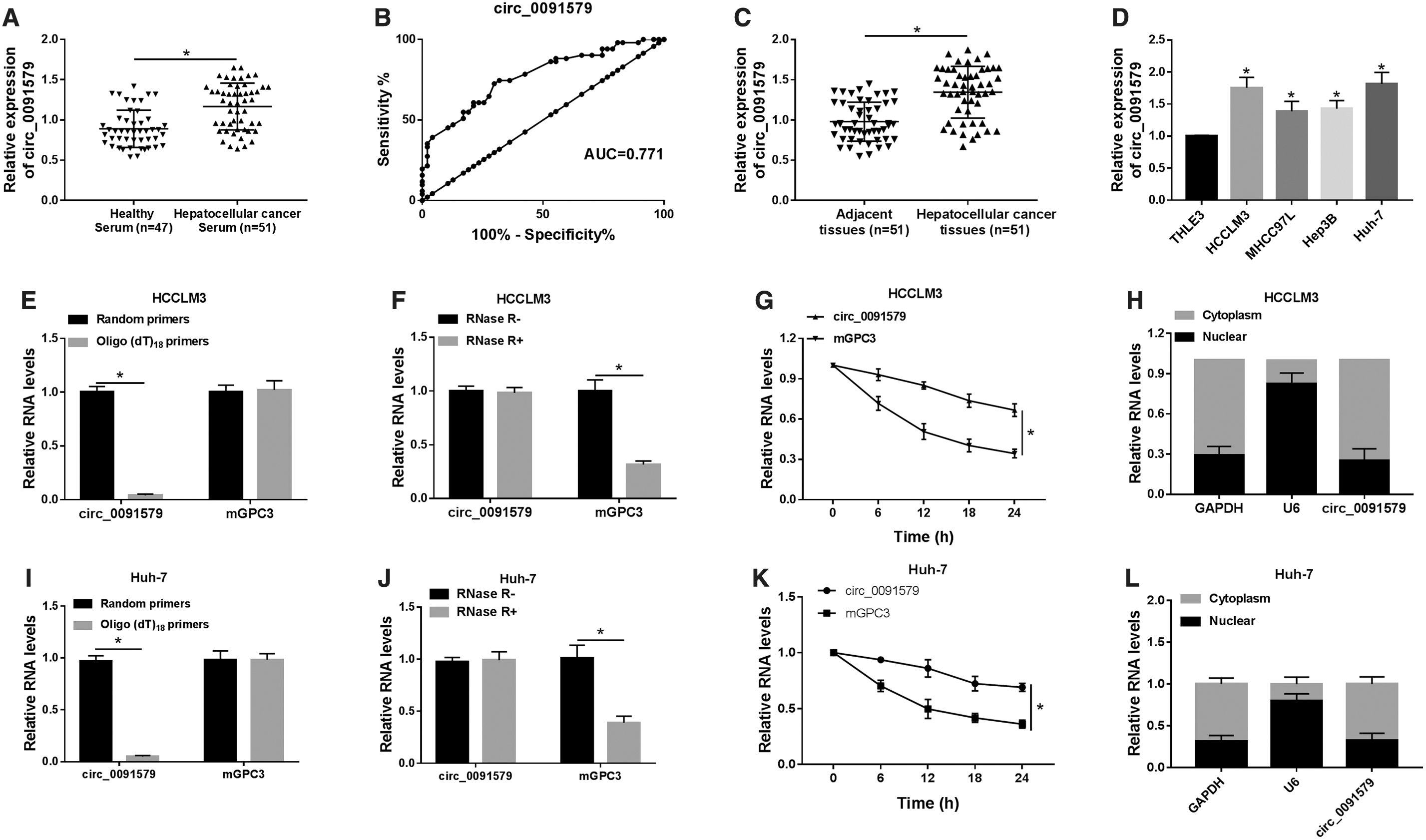
The expression of circ_0091579 was upregulated in HCC serums, tissues, and cell lines.
To further validate the diagnostic value of circ_0091579, 51 pairs of HCC tissues and corresponding adjacent normal tissues from the patients were used. The expression of circ_0091579 was found to be strikingly increased in HCC tissues compared with that in corresponding adjacent normal nontumor tissues (Fig. 1C). Meanwhile, the expression level of circ_0091579 in normal human liver epithelial cell line (THLE3) and HCC cell lines (HCCLM3, MHCC97L, Hep3B, and Huh-7) was further measured. Similarly, the results discovered that the expression level of circ_0091579 was strongly elevated in all detected HCC cells compared with that in THLE3 cells (Fig. 1D).
Besides, the expression level of circ_0091579 in HCCLM3 and Huh-7 cells was higher than any other cells detected here; thus, the two cells were subjected to succedent tests. Then, HCC-derived HCCLM3 cells were reversely transcribed with random hexamer primers or oligo (dT)18 primers to confirm the circular characteristics of circ_0091579. The relative expression of circ_0091579 was obviously downregulated in oligo (dT)18 primer group, while mGPC3 was not changed, implying that circ_0091579 had no poly-A tail (Fig. 1E, I). Furthermore, circ_0091579 was resistant to RNase R, indicating that circ_0091579 is circular (Fig. 1F, J).
Moreover, actinomycin D was used to inhibit transcription followed by the measure for the half-life of circ_0091579 and mGPC3 in HCCLM3 cells. The results showed that circ_0091579 was more stable than mGPC3 (Fig. 1G, K). In addition, the aggregation of circ_0091579 at subcellular level was detected by qRT-PCR, and it revealed that circ_0091579 was predominantly distributed in cytoplasm rather than nuclear in HCCLM3 cells (Fig. 1H, L). Collectively, all these data demonstrated that circ_0091579 was significantly upregulated in HCC serums, tissues, and cell lines with a circular and stable structure.
Knockdown of circ_0091579 inhibited cell proliferation, migration, invasion, and glycolysis in HCC cells
Next, HCCLM3 and Huh-7 cells were transfected with si-circ_0091579 or si-NC to explore the role of circ_0091579 in the progression of HCC. qRT-PCR results showed that the expression of circ_0091579 in HCCLM3 and Huh-7 cells transfected with si-circ_0091579 was significantly downregulated, which validated the transfection efficiency (Fig. 2A). MTT assay revealed that si-circ_00915791-mediated circ_0091579 silencing triggered a notable reduction of cell proliferative ability in HCCLM3 and Huh-7 cells (Fig. 2B). Consistently, the migration (Fig. 2C) and invasion (Fig. 2D) capacities of HCCLM3 and Huh-7 cells transfected with si-circ_0091579 were sharply decreased compared with si-NC-transfected cells.
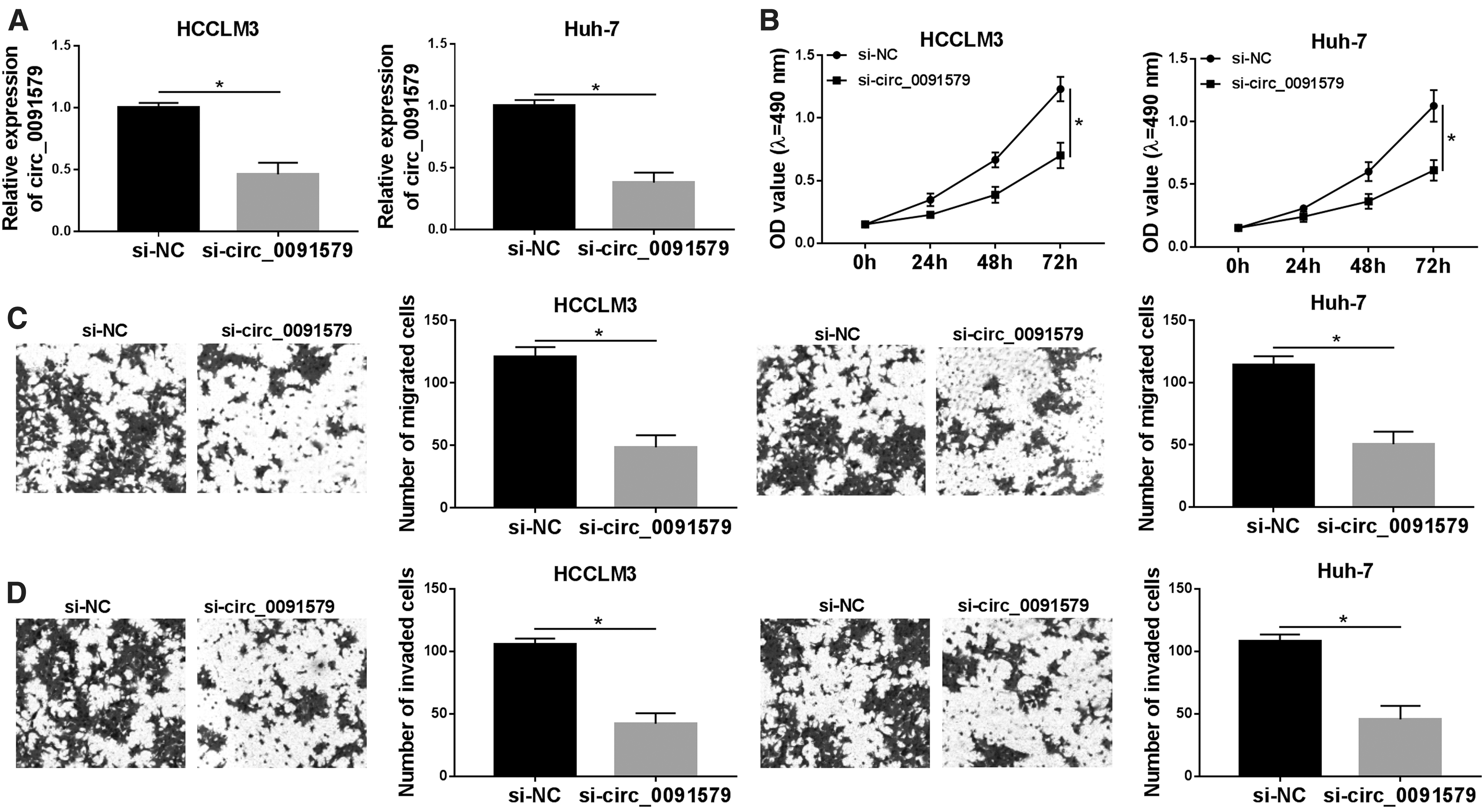
circ_0091579 knockdown inhibited cell proliferation, migration, invasion, and glycolysis in HCC cells.
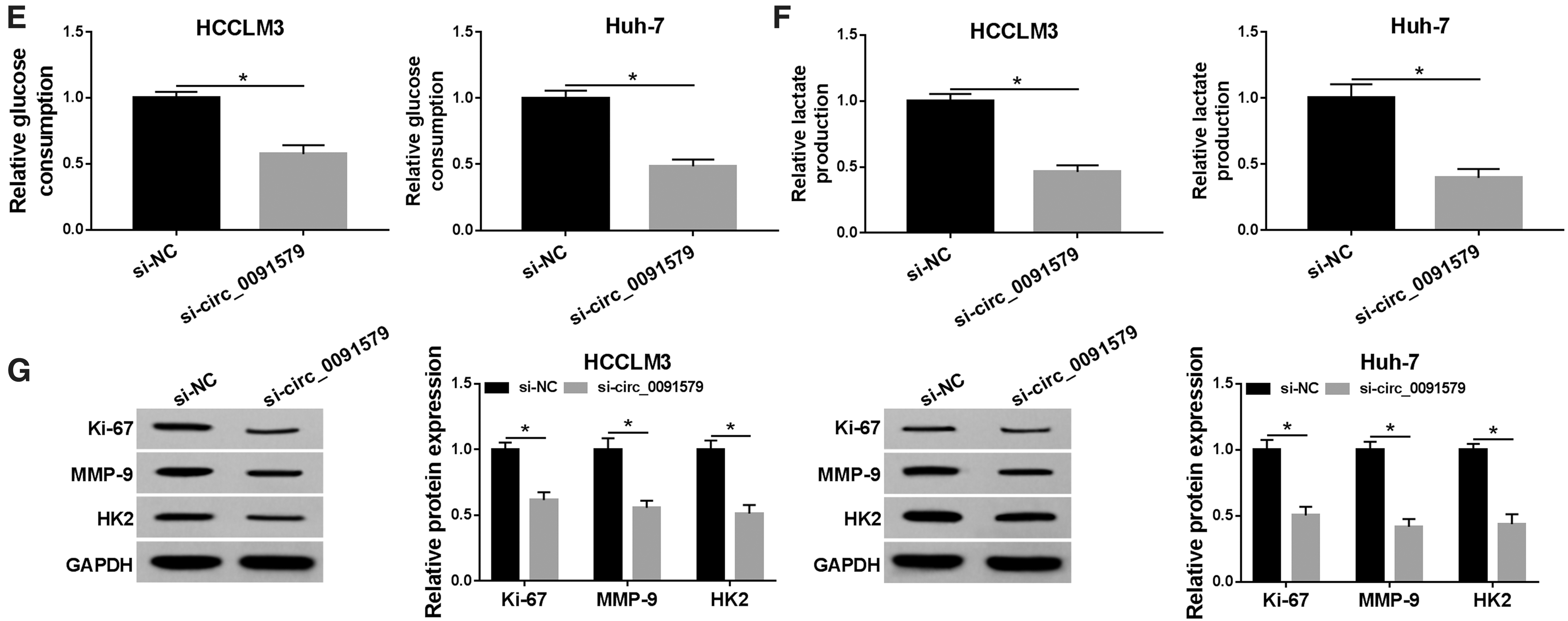
Subsequently, the level of glycolysis was evaluated by measuring the glucose consumption and lactate production. The results suggested that deficiency of circ_0091579 attenuated glucose consumption (Fig. 2E) and lactate production (Fig. 2F), clarifying that si-circ_0091579 diminished glycolysis in HCC cells. In addition, the protein levels of proliferation marker Ki-67, metastasis-related marker MMP-9, and glycolysis-related protein HK2 were notably reduced by si-circ_0091579 in HCCLM3 and Huh-7 cells (Fig. 2G). Overall, these findings implied that deficiency of circ_0091579 suppressed cell proliferation, migration, invasion, and glycolysis in HCC cells.
circ_0091579 targeted miR-490-5p and negatively regulated miR-490-5p expression in HCC cells
To explore the association between circ_0091579 and miR-490-5p, the binding sites between them were predicted by Circular RNA Interactome database (Fig. 3A). Then, the luciferase reporter vectors (WT-circ_009157 or MUT-circ_009157) containing the wild-type or mutated miR-490-5p-binding sequence in circ_009157 were constructed and cotransfected with miR-490-5p or miR-NC into HCCLM3 and Huh-7 cells. The results indicated that the luciferase activity of WT-circ_009157 was distinctly reduced in HCCLM3 and Huh-7 cells cotransfected with miR-490-5p, whereas the luciferase activity of MUT-circ_009157 was insusceptible (Fig. 3B). Meanwhile, the expression level of miR-490-5p in HCC tissues, as well as in HCCLM3 and Huh-7 cells, was significantly downregulated and tested by qRT-PCR (Fig. 3C, D).
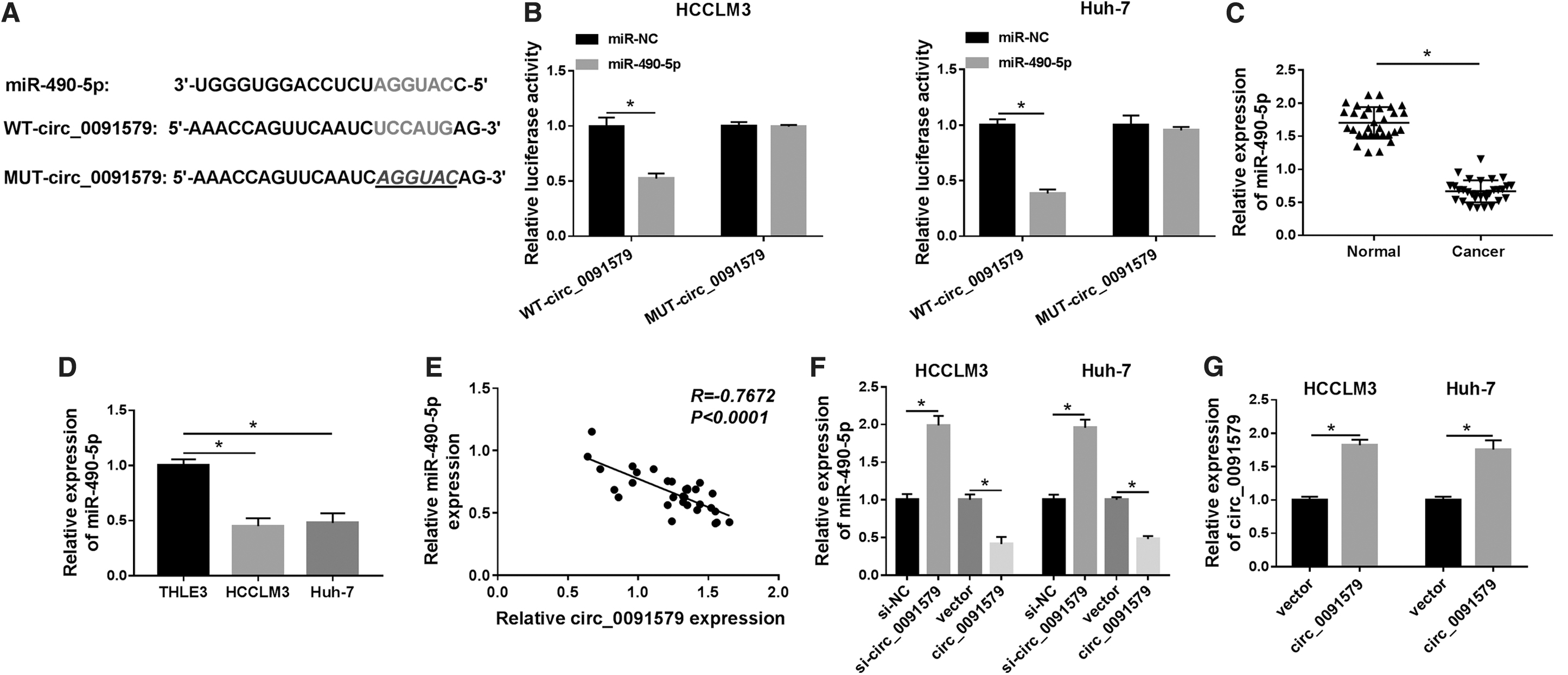
circ_0091579 targeted miR-490-5p and inhibited miR-490-5p expression.
Besides, the expression of circ_0091579 was negatively correlated with miR-490-5p expression in HCC tissues (Fig. 3E). As expected, the expression of circ_009157 was obviously elevated in HCCLM3 and Huh-7 cells after overexpression of circ_009157 (Fig. 3F). Gain- and loss-of-function assays disclosed that the expression of miR-490-5p in both HCCLM3 and Huh-7 cells transfected with si-circ_009157 was evidently upregulated relative to that of si-NC group, whereas the expression of miR-490-5p in both HCCLM3 and Huh-7 cells transfected with circ_009157 was obviously downregulated compared with that of vector-transfected cells (Fig. 3G). Therefore, it can be concluded that miR-490-5p was a target of circ_009157 and negatively regulated by circ_009157 in HCC cells.
circ_009157 regulated cell proliferation, migration, invasion, and glycolysis through miR-490-5p in HCC cells
In view of the role of circ_009157 in HCC cells and the association between circ_009157 and miR-490-5p, whether the effects of circ_009157 on cell proliferation, migration, invasion, and glycolysis were correlative with miR-490-5p was further explored. The upregulated expression of miR-490-5p in HCCLM3 and Huh-7 cells transfected with miR-490-5p indicated a successful transfection (Fig. 4A). The proliferation ability of HCCLM3 and Huh-7 cells transfected with miR-490-5p was obviously declined compared with that in miR-NC group, while the effect was partly overturned by circ_009157 overexpression (Fig. 4B). Moreover, the miR-490-5p-induced decrease in the migration (Fig. 4C) and invasion (Fig. 4D) abilities of HCCLM3 and Huh-7 cells was also partially reversed by circ_009157 overexpression.

circ_0091579 enhanced cell proliferation, migration, invasion, and glycolysis through miR-490-5p in HCC cells.
In addition, overexpression of miR-490-5p weakened glucose consumption (Fig. 4E) and lactate production (Fig. 4F), while these effects were partly eliminated by circ_0091579. Meanwhile, the decreased expression of Ki-67, MMP-9, and HK2 was regained through upregulation of circ_0091579 in HCCLM3 and Huh-7 cells (Fig. 4G). These data suggested that circ_009157 might regulate cell proliferation, migration, invasion, and glycolysis by targeting miR-490-5p in HCC cells.
CASC3 was a target of miR-490-5p in HCC cells
To further explore the role of miR-490-5p in HCC cells, TargetScan online database was used to predict the targeted gene of miR-490-5p. The results demonstrated that there was a binding site between miR-490-5p and CASC3 (Fig. 5A). The reporter plasmids WT-CASC3 containing the predicted miR-490-5p binding sites and mutant MUT-CASC3 were constructed. As shown in Figure 5B, the luciferase activity in HCCLM3 and Huh-7 cells was greatly limited when cotransfected with miR-490-5p and WT-CASC3. Then, the mRNA and protein expression levels of CASC3 in HCC tissues and cells were examined by qRT-PCR and Western blot. The results indicated that the expression of CASC3 in HCC tissues, as well as in HCCLM3 and Huh-7 cells, was significantly increased compared with normal tissues and THLE3 cells, respectively (Fig. 5C, D).
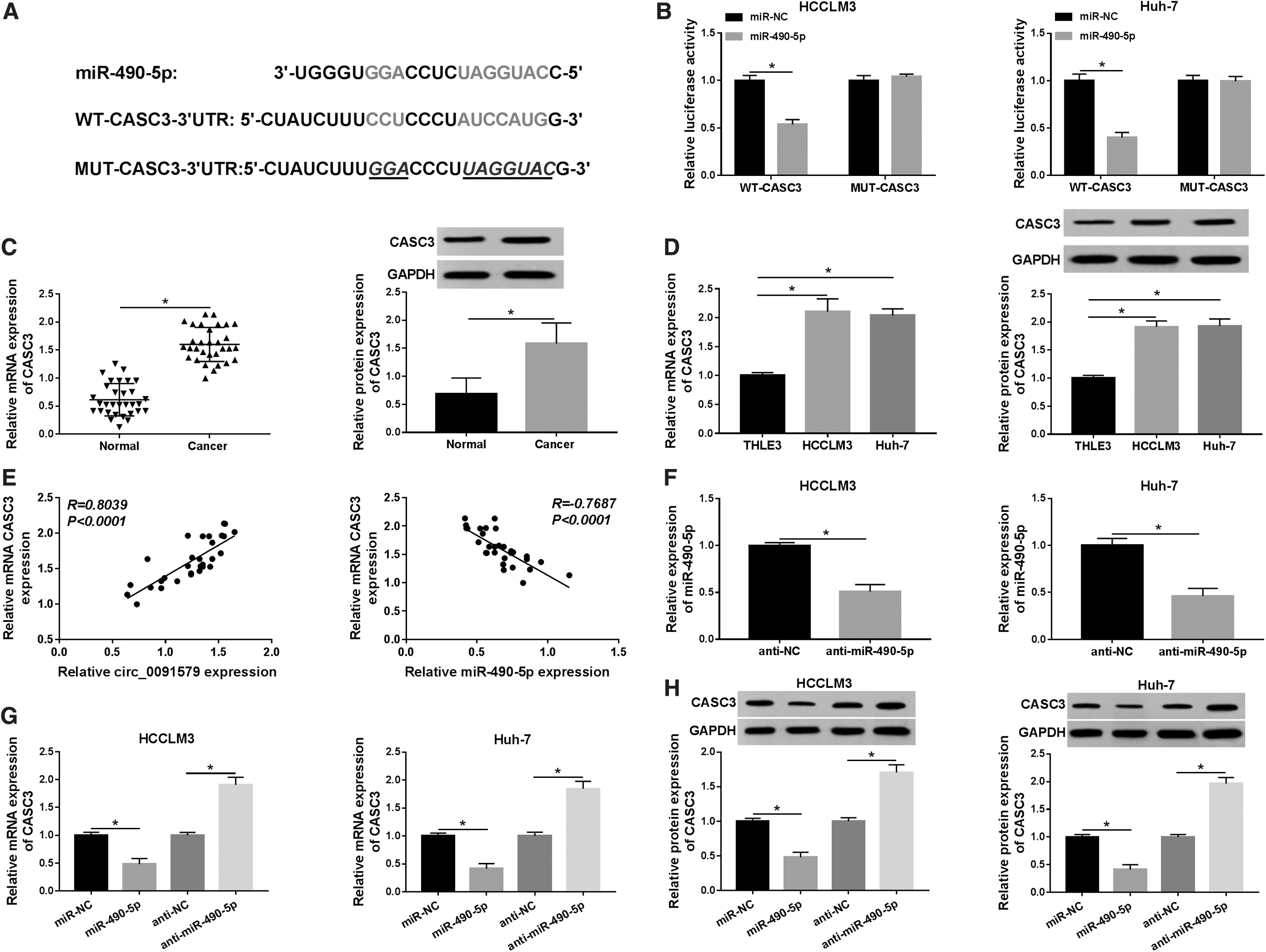
CASC3 was a target of miR-490-5p in HCC cells.
In addition, the level of CASC3 in HCC tissues was positively correlated with circ_009157 level and negatively correlated with miR-490-5p level (Fig. 5E). The effect of miR-490-5p knockdown (anti-miR-490-5p) was determined by qRT-PCR (Fig. 5F). The results showed that miR-490-5p resulted in an obvious inhibition in CASC3 mRNA and protein expression in HCCLM3 and Huh-7 cells, whereas the effects were opposite by transfection of anti-miR-490-5p (Fig. 5G, H). All these evidences showed that miR-490-5p directly targeted CASC3 and regulated the expression of CASC3 in HCC cells.
Knockdown of miR-490-5p reversed the effects of CASC3 silencing on cell proliferation, migration, invasion, and glycolysis in HCC cells
Based on the above results, knockdown experiments were proceeded to further verify the correlation between miR-490-5p and CASC3 in HCC cells. The mRNA and protein expression levels of CASC3 in HCCLM3 and Huh-7 cells transfected with si-CASC3 were apparently downregulated relative to si-NC-transfected cells, implying a successful transfection (Fig. 6A, B). Cell proliferation (Fig. 6C), migration (Fig. 6D), and invasion (Fig. 6D) capacities of HCCLM3 and Huh-7 cells transfected with si-CASC3 were overtly reduced in comparison with those of si-NC group, while the reduction was partly abrogated when transfected with si-CASC3 and anti-miR-490-5p.

MiR-490-5p suppressed cell proliferation, apoptosis, migration, and invasion through CASC3 in HCC cells.

Furthermore, the glucose consumption (Fig. 6F) and lactate production (Fig. 6G) were both distinctly declined in HCCLM3 and Huh-7 cells transfected with si-CASC3, whereas the effects were dramatically abated by anti-miR-490-5p. Consistently, si-CASC3-mediated reduced protein levels of Ki-67, MMP-9, and HK2 were all restored by cotransfection with anti-miR-490-5p (Fig. 6H). From the above, the findings indicated that miR-490-5p inhibited cell proliferation, migration, invasion, and glycolysis by regulating CASC3 in HCC cells.
circ_009157 regulated the expression of CASC3 by sponging miR-490-5p in HCC cells
To further explore the relationship among circ_009157, miR-490-5p, and CASC3, the mRNA and protein expression levels of CASC3 in HCCLM3 and Huh-7 cells transfected with miR-NC, miR-490-5p, miR-490-5p + vector, or miR-490-5p + circ_009157 were measured by qRT-PCR and Western blot, respectively. The results demonstrated that the expression of CASC3 in HCCLM3 and Huh-7 cells transfected with miR-490-5p or miR-490-5p + vector was significantly downregulated, while the decrease was effectively alleviated in miR-490-5p + circ_009157 group (Fig. 7A, B). Thus, it was concluded that circ_009157 could regulate CASC3 expression through miR-490-5p in HCC cells.

circ_0091579 upregulated the expression of CASC3 through miR-490-5p in HCC cells.
Discussion
HCC is one of the most prevalent and aggressive cancers worldwide, which severely threatens the human health. The progression of HCC is a multigene and multistage process. 35 –37 However, the therapeutic methods of HCC are limited until now. Thus, it is critical to explore the underlying mechanisms of HCC progression and search for early diagnostic or prognostic biomarkers and therapeutic targets for HCC.
An increasing number of circRNAs were verified to function as miRNA sponges to repress their function and served as novel biomarkers for early diagnosis and prognosis of diseases. 8,38,39 For example, circRNA_001569 regulated cell proliferation and invasion of colorectal cancer by targeting miR-145. 12 circRNA PIP5K1A affected the progression of colon cancer by inhibiting miR-1273a expression. 13 Moreover, circ_0137606/miR-1231 pathway was involved in the tumorigenesis of bladder cancer. 40
Recent studies suggested that a few circRNAs were abnormally expressed and played essential roles in HCC by regulating cell proliferation and metastasis by targeting different miRNAs. 15 Nevertheless, circ_0091579 was rarely reported in cancers, hence the function of it needed to be further validated. Zhang et al. reported that circ_0091579 was overexpressed and might work as a diagnostic marker in HCC. 18 Consistent with these reports, this study certified that the level of circ_0091579 in HCC tissues and cell lines was notably elevated relative to that of corresponding normal tissues or normal cell lines, respectively. Moreover, cell proliferation, glycolysis, migration, and invasion abilities were apparently repressed in HCC cells after circ_0091579 silencing. These findings indicated that circ_0091579 acted as a cancer-promoting gene in HCC progression.
Increasing data disclosed that multiple miRNAs were abnormally expressed in HCC and served as tumor suppressors or oncogenes to participate in tumor progression. 41,42 miR-490-5p was a pivotal regulatory factor in various kinds of cancers. 43,44 It has been reported that miR-490-5p has many different targets in cancer, such as c-FOS and EGFR in bladder cancer, PIK3CA in esophageal squamous cell carcinoma and renal carcinoma, and so on. 23,43 –45 Recent studies indicated that miR-490-5p was strikingly downregulated in HCC tissues compared with that in normal liver tissues and could inhibit HCC cell proliferation and metastasis by regulating different downstream target genes. 27,46 For example, hsa_circ_0103809 could accelerate HCC progression through miR-490-5p/SOX2 axis. 28
In line with these findings, their research found that the expression of miR-490-5p was reduced in HCC tissues and cells. Simultaneously, it was verified that miR-490-5p was a target of circ_0091579 and negatively regulated by circ_0091579. Furthermore, this study also confirmed that the blocked influences of miR-490-5p on cell proliferation, migration, invasion, and glycolysis were partly reversed by circ_0091579 overexpression. From these data, it was testified that circ_0091579 could regulate cell proliferation, migration, invasion, and glycolysis through targeting miR-490-5p in HCC cells.
CASC3 is a nucleocytoplasmic shuttling protein and it has been demonstrated to be overexpressed in breast cancer. 32 However, the role of CASC3 in HCC remains elusive. Xu et al. suggested that CASC3 silencing inhibited cell proliferation in HCC cells by serving as a target of miR-124-1, implying that CASC3 might function as a cancer-promoting gene. 34
Similarly, this study suggested that the level of CASC3 was enhanced in HCC tissues and cells, and deficiency of CASC3 impeded cell proliferation, migration, invasion, and glycolysis in HCC cells. Besides, this study evidenced that CASC3 was targeted by miR-490-5p, and miR-490-5p knockdown partly restored the effects of CASC3 silencing on HCC progression. In addition, it was proved that overexpression of circ_0091579 largely overturned the blocked influences of miR-490-5p overexpression on the mRNA and protein levels of CASC3. Taken together, all these findings illuminated that circ_0091579 facilitated cancer malignant behaviors, at least partly, by regulating miR-490-5p/CASC3 axis in HCC.
However, there are some limitations in this study. For instance, the in vivo data were lacking in the experiments, and the commercial cells could not replicate the in vivo environment. Hence, to further investigate the roles of circ_0091579 in HCC development, the animal experiments should be performed in the future work. In addition, the results of RNA pull-down and RNA immunoprecipitation assays still need to be supplemented to verify the targeting correlation between miR-490-5p and circ_0091579 or CASC in future research.
Conclusion
This study identified the participation of circ_0091579, miR-490-5p, and CASC3 in HCC tissues and cells. circ_0091579 could promote cell proliferation, migration, invasion, and glycolysis by sponging miR-490-5p to upregulate CASC3 expression. Therefore, this study contributed to elucidate the molecular mechanisms of circ_0091579 in HCC progression and might provide a potential therapeutic target for HCC treatment.
Availability of Data and Materials
The analyzed data sets generated during the present study are available from the corresponding author on reasonable request.
Ethics Approval and Consent to Participate
The present study was approved by the Ethics Review Committee of Luhe Hospital Affiliated to Capital Medical University. Written informed consent was obtained from all enrolled patients.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.