
Abstract
Background:
Long noncoding RNAs (lncRNAs) X inactivate-specific transcripts (XIST) have been found to be dysregulated in breast cancer (BC). Nevertheless, the influence and mechanism of XIST on BC progression remain largely undefined.
Methods:
The expression levels of XIST, miR-362-5p, and ubiquitin-associated protein 1 (UBAP1) mRNA were detected by quantitative real-time polymerase chain reaction. Cell proliferation, apoptosis, migration, and invasion abilities were determined using MTT assay, flow cytometry, and transwell assay, respectively. Targeted relationship between miR-362-5p and XIST or UBAP1 was validated by the dual-luciferase reporter assay. Western blot was performed to evaluate UBAP1 protein level. Xenograft mice model was established for the investigation of XIST in tumor growth.
Results:
The authors' data indicated that XIST and UBAP1 were downregulated in BC tissues and cells. XIST overexpression weakened BC cell proliferation, migration, invasion, and facilitated the apoptosis, and XIST silencing exerted opposite effect. Mechanistically, XIST directly interacted with miR-362-5p and miR-362-5p mediated the regulatory effects of XIST overexpression on BC cell malignant behaviors. UBAP1 was a direct target of miR-362-5p. MiR-362-5p exerted its regulatory effects on BC cell behaviors by UBAP1. Moreover, XIST modulated UBAP1 expression through acting a competing endogenous RNA of miR-362-5p. XIST overexpression mediated antiproliferation, antimigration, anti-invasion, and proapoptosis effects were abated by the restored expression of UBAP1 in BC cells. Furthermore, the upregulation of XIST hindered tumor growth in vivo.
Conclusion:
The current study suggested that XIST overexpression hampered BC cell progression in vitro and in vivo at least partially by targeting the miR-362-5p/UBAP1 axis, illuminating XIST as a promising therapeutic agent for BC management.
Introduction
Breast cancer (BC) remains the most prevalent malignancy and the leading cause of cancer-related death in women worldwide. 1 Due to the westernized lifestyle, obesity, and physical inactivity, the incidence of BC is rapidly rising in China. 2 Conventional treatments, such as mastectomy, radiotherapy, and adjuvant chemotherapy, have recently improved the prognosis of primary BC patients. 3 Nevertheless, the survival rate of patients with advanced BC remains unsatisfactory. Hence, a better understanding for the pathogenesis of BC progression is very imperative for BC management.
In recent years, noncoding RNAs (ncRNAs) have attracted increasing attention for their regulatory functions in cellular biological processes from normal development to tumorigenesis. 4 The development of sequencing technologies has resulted in the discovery of a large number of ncRNAs species, such as long noncoding RNAs (lncRNAs) and microRNAs (miRNAs). 5,6 LncRNAs are endogenous >200 nucleotide RNAs that harbor a broad range of molecular and cellular functions. 7 MiRNAs are small, highly conversed ∼22 nucleotide RNA molecules that play crucial gene-regulatory roles by pairing to the 3′-untranslated regions (3′-UTR) of target mRNAs. 8 The competing endogenous RNAs (ceRNAs) hypothesis proposes that lncRNAs can function as miRNAs sponges to protect the target mRNAs from repression in cancer biology. 9
X inactivate-specific transcript (XIST), a 19 kb lncRNA from the inactive X-chromosome, is the vital regulator of X chromosome inactivation, whose alteration occurs in BC cells. 10 XIST has been found to be prominently downregulated in BC. 11,12 Moreover, Xing et al. uncovered that the loss of XIST accelerated brain metastasis in BC partially through the increase of exosomal miR-503. 11 Zheng et al. underscored that the elevated expression of XIST weakened BC cell growth and metastasis, highlighting XIST as a therapeutic agent for BC treatment. 12 Nevertheless, the underlying mechanism of XIST on BC progression remain mostly undiscovered.
Previous researches had reported that miR-362-5p was highly expressed in BC, and its knockdown hindered MCF7 cell malignant behaviors. 13,14 XIST was manifested to suppress BC cell progression in vitro by sponging miR-155 and regulating caudal-type homeobox 1 (CDX1) expression. 12 Nevertheless, whether miR-362-5p is a mediator of XIST on BC cell behaviors is still undefined.
In this research, the authors first determined the effects of XIST on BC cell proliferation, migration, invasion, and apoptosis. Consequently, they further investigated the molecular mechanism of XIST on BC cell progression.
Materials and Methods
Clinical specimens and cell culture
In the current project, clinical samples were collected from 30 BC patients confirmed at Beijing Anzhen Hospital, Capital Medical University between October 2016 and December 2017, with written informed consent. The clinicopathological features of these patients are presented in Table 1. The 30 corresponding normal breast tissues were located at least 5 cm away from the primary tumor. These fresh specimens were cryopreserved at −80°C until use. This research was approved by the Ethics Committee of Beijing Anzhen Hospital, Capital Medical University, and all patients gave written informed consent.
Clinicopathological Features of These Patients (n = 30)
ER, estrogen receptor; PR, progesterone receptor; HER2, human epidermal growth factor receptor type 2.
Human normal breast cell line (CCD-1095SK), two BC cell line (MCF7 and MDA-MB-231), and 293T cells (ATCC®CRL-11268) were obtained from ATCC (Manassas, VA). CCD-1095SK, MCF7, and 293T cells were maintained in EMEM (ATCC), and MDA-MB-231 cells were cultured in Leibovitz's L-15 Medium (ATCC) containing 10% FBS (Bovogen Biologicals, Essendon, Australia), 1% penicillin/streptomycin (Biochrom, Cambourne, United Kingdom) at 37°C in 5% CO2.
Plasmid and oligonucleotide transfection
For the upregulation of XIST, 10 ng of corresponding overexpression plasmid (pcDNA-XIST) was introduced into cells, and the nontarget plasmid (pcDNA-Control) was used as a negative control. XIST or ubiquitin associated protein 1 (UBAP1) silencing was achieved using 20 nM of siRNA targeting XIST (si-XIST) or UBAP1 (si-UBAP1), with nontarget siRNA (si-NC) as the negative control. For the alteration of miR-362-5p expression, the cells were transfected with 20 nM of the synthetic miR-362-5p mimic, miR-362-5p inhibitor, or a corresponding nontarget sequence (miR-NC mimic or inhibitor NC). The DharmaFECT 4 transfection reagent (Dharmacon, Lafayette, CO) was used for each transfection. All plasmids and oligonucleotides were obtained from GenePharma (Shanghai, China).
Quantitative real-time polymerase chain reaction
An iPrep™ TRIzol™ Plus RNA Kit (Invitrogen, Breda, the Netherlands) was used for the extraction of total RNA, following the manufacturer's recommendations. For miR-362-5p quantification, cDNA synthesis was implemented using the TaqMan MicroRNA RT Kit (Applied Biosystems, Rotkreuz, Switzerland); quantitative real-time polymerase chain reaction (RT-qPCR) was carried out with a mirVana™ miRNA Detection Kit (Ambion, Darmstadt, Germany); and U6 was used for normalization.
For the determination of XIST and UBAP1 mRNA, the Maxima H Minus First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Passau, Germany) was used for cDNA synthesis; RT-qPCR was conducted using the FastStart SYBR Green I Kit (Roche, Charente, France); GAPDH was used for normalization. The primers for amplification (5′-3′) were listed: miR-362-5p: GTCACGAAATCCTTGGAACCTAG (sense) and TATGGTTGTTCTCGTCTCCTTCTC (antisense); U6: TCGCTTCGGCAGCACATATAC (sense) and CGCTTCACGAATTTGCGTG (antisense); XIST: TCAGCCCATCAGTCCAAGATC (sense) and CCTAGTTCAGGCCTGCTTTTCAT (antisense); UBAP1 mRNA: CAGGTTCTAAATGGCTTC (sense) and ATGTGCTATGGAAAGTGC (antisense); GAPDH: TCTCTGCTCCTCCCTGTTC (sense) and ACACCGACCTTCACCATCT (antisense).
Cell proliferation assay
Cells (5.0 × 104) in 96-well microplates were transfected with the indicated plasmids and/or oligonucleotides. After transfection at 0, 24, 48, and 72 h, the MTT solution (Solarbio, Beijing, China) was used at a final concentration of 0.5 mg/mL per well before adding the dimethyl sulfoxide (DMSO, 150 μL) for 2 h. Cell proliferation was detected by measuring formazan concentration, which was reflected at an optical density of 490 nm using a microplate reader (Thermo Fisher Scientific).
Flow cytometry for cell apoptosis
Transfected cells (1.0 × 106) were washed and harvested following trypsinization (0.05% trypsin/EDTA; Invitrogen). The Apoptosis Detection Kit (Thermo Fisher Scientific) was used to assess cell apoptosis, following the producer's guidance.
Transwell migration and invasion assay
The modified 24-Transwell plates (8-μm pores, Corning Life Sciences, Amsterdam, The Netherlands) were used for the determination of cell migration and invasion abilities. In migration assays, transfected cells (1.0 × 104) were suspended in media with 1% FBS and placed into the top chamber. In invasion assays, transfected cells (1.0 × 105) were added into the upper compartment with Matrigel-precoated insert (Corning Life Sciences). In both assays, 600 μL of 10% FBS medium was placed into the bottom chamber as chemoattractant. Twenty-four hours later, the migrated or invaded cells were stained with 0.2% crystal violet (Solarbio). Images were photographed under a microscope (Nikon, Shinagawa, Tokyo, Japan) and the penetrated cells were counted at five random fields at × 100 magnification.
Bioinformatics and dual-luciferase reporter assay
The online starBase v.3.0 software was used to predict the miRNAs that potentially bind to XIST and miR-362-5p molecular targets. XIST luciferase reporter plasmid containing miR-362-5p-binding sequence (WT-XIST) and its mutant in the seed region (MUT-XIST), and UBAP1 3′-UTR reporter construct (WT-UBAP1–3′-UTR) and the site-directed mutagenesis of the target sequence (MUT-UBAP1–3′-UTR) were designed and synthesized by Hanbio Biotechnology (Shanghai, China). Fifty nonograms of WT-XIST, MUT-XIST, WT-UBAP1–3′-UTR, or MUT-UBAP1–3′-UTR was transfected into 293T cells, together with 20 nM of miR-362-5p mimic or miR-NC mimic. Luciferase activity of firefly and Renilla were simultaneously measured 48 h posttransfection using the Dual-luciferase Reporter Assay System Kit (Promega, Madison, WI) and GloMax Luminometer.
Western blot for UBAP1
Equal amount of total protein was separated by a SDS polyacrylamide gel and then electroblotted onto a polyvinylidene fluoride membrane (PVDF, GE Health Care, Little Chalfont, United Kingdom). The membranes were incubated with antibodies against UBAP1 (ab89124; Abcam, Cambridge, United Kingdom) or GAPDH (ab181602; Abcam). The ECL detection system was utilized to determine the protein bands by photographing with the ChemiDoc XRS System (Bio-Rad Laboratories, Aldermaston, United Kingdom).
Lentiviral vector transduction
Lentiviral vectors containing XIST-encoding sequence (lenti-XIST) or a scrambled negative sequence (lenti-control) were constructed by Fulengen (Guangzhou, China). MDA-MB-231 cells were infected by these lentiviral vectors in media containing 8 μg/mL of polybrene (Sigma-Aldrich, Steinheim, Germany). One day later, the cells with positive infection were screened by puromycin (Sigma-Aldrich).
Xenograft model assay
To perform in vivo assays, 6-week-old BALB/c nude mice (Guangdong Research Center of Laboratory Animal, Foshan, China) were used in this study, after obtaining approval from Animal Ethics Committee of Beijing Anzhen Hospital, Capital Medical University. These mice were housed at the animal facility in a specific-pathogen-free environment. Approximately 5.0 × 106 lenti-XIST-transducing MDA-MB-231 cells were subcutaneously implanted into the nude mice, with lenti-NC-transducing cells as the negative control (10 mice per group). In the end, all mice were killed and tumor tissues were excised.
Statistical analysis
The data were analyzed by a Student's t-test or one-way ANOVA. All experiments were conducted by three biological replicates × 2 technical replicates. p values <0.05 meant that the difference was statistically significant.
Results
XIST and UBAP1 were downregulated in BC tissues and cells
First, the authors determined the expression levels of XIST and UBAP1 in BC tissues and corresponding noncancerous breast tissues. As demonstrated by RT-qPCR, XIST (Fig. 1A) and UBAP1 mRNA (Fig. 1B) levels were significantly reduced in BC tissues compared with normal controls. After that, they assessed XIST and UBAP1 expression in BC cells (MCF7 and MDA-MB-231) and human normal breast CCD-1095SK cells. In line with BC tissues, the expression levels of XIST (Fig. 1C) and UBAP1 mRNA (Fig. 1D) were remarkably downregulated in BC cells when comparing with normal controls.
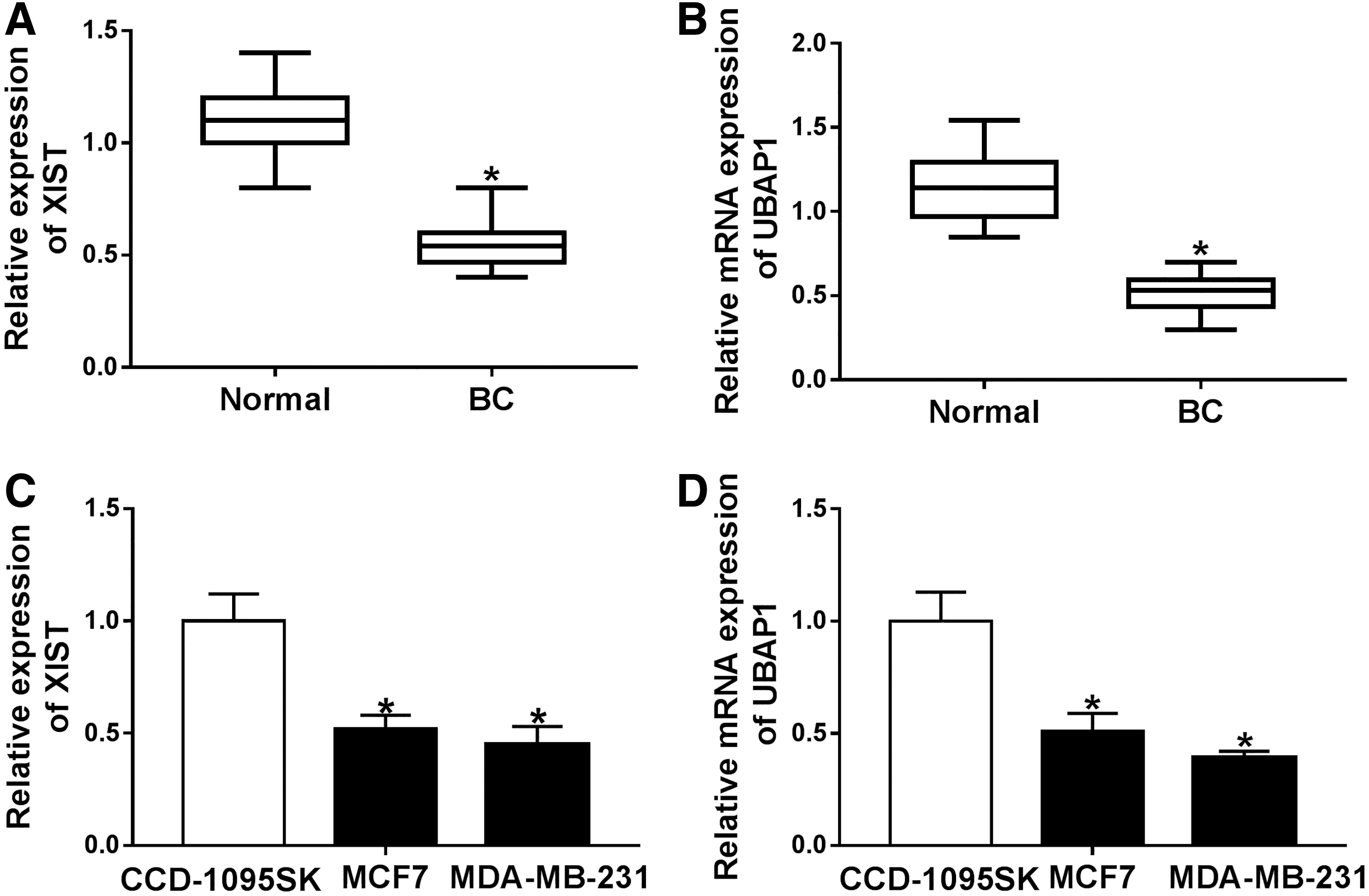
XIST and UBAP1 were downregulated in BC tissues and cells.
XIST regulated BC cell proliferation, migration, and invasion and facilitated the apoptosis
To observe the role of XIST in BC progression, gain-of-function experiments were carried out using XIST overexpression plasmid (pcDNA-XIST). Transient introduction of pcDNA-XIST, but not a negative plasmid, markedly elevated the expression of XIST in both MCF7 and MDA-MB-231 cells (Fig. 2A). MTT assays revealed that cell proliferation ability was dramatically weakened by XIST overexpression in the two BC cells (Fig. 2B, C). Flow cytometry results showed that in contrast to negative control, XIST overexpression triggered a significant promotion in cell apoptosis (Fig. 2D). Moreover, transwell assays demonstrated that the high expression of XIST led to a distinct suppression in cell migration (Fig. 2E) and invasion (Fig. 2F). In addition, the authors knocked down the expression of XIST in MCF7 and MDA-MB-231 cells using si-XIST. RT-qPCR data revealed that XIST expression was prominently reduced by si-XIST compared with the negative group (Supplementary Fig. S1A). Moreover, XIST silencing resulted in a significant enhancement in cell proliferation, a distinct inhibition in cell apoptosis, as well as a clear promotion in cell migration and invasion (Supplementary Fig. S1B–F).
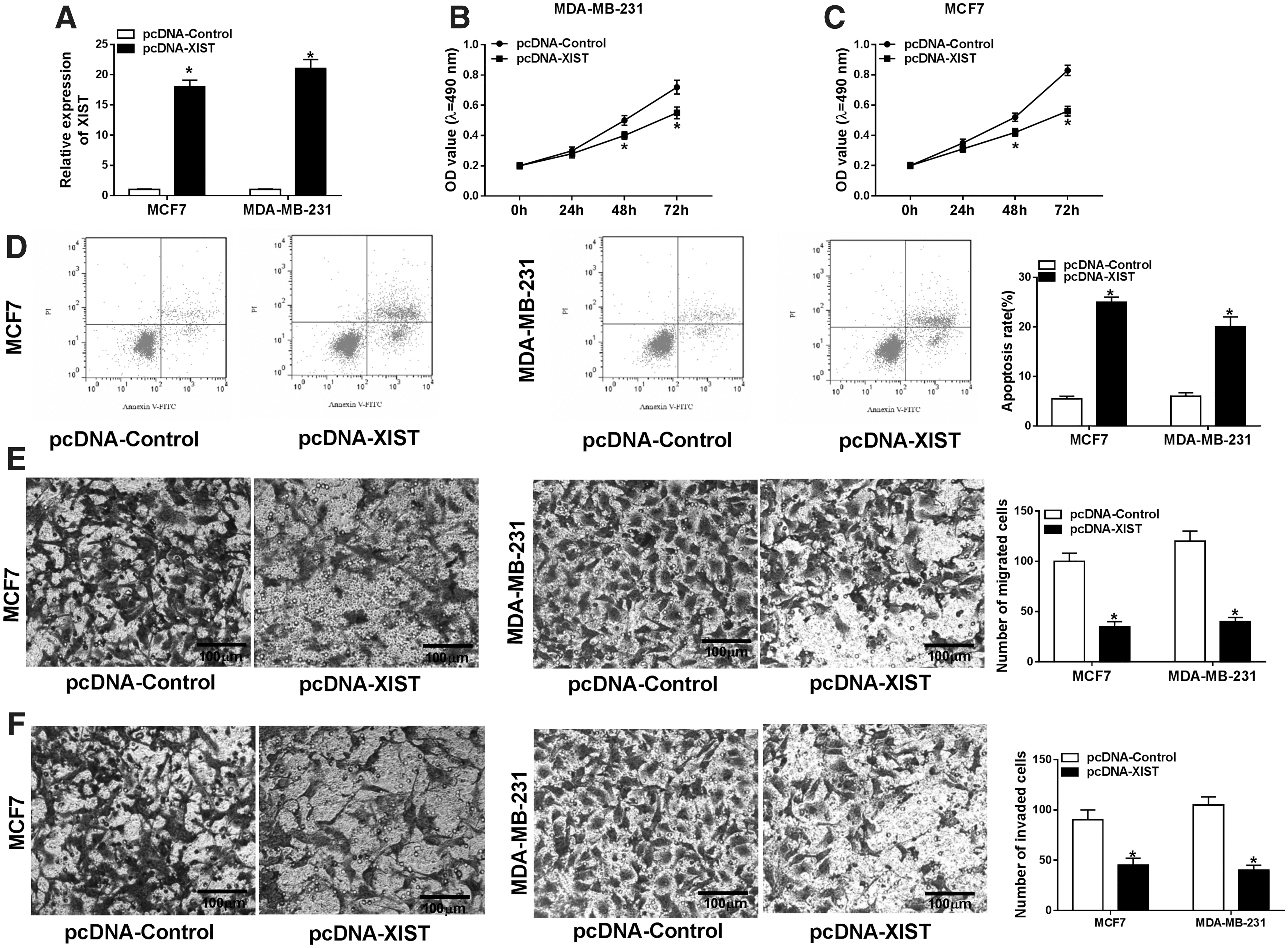
XIST overexpression repressed BC cell proliferation, migration, invasion, and facilitated the apoptosis. MCF7 and MDA-MB-231 cells were transfected with pcDNA-control or pcDNA-XIST for the indicated time point.
MiR-362-5p directly interacted with XIST and mediated the regulation of XIST overexpression on BC cell malignant behaviors
To understand the mechanisms by which XIST influenced BC cell progression in vitro, the authors implemented a detailed analysis for the interacted miRNAs of XIST. Using the online starBase v.3.0 software, the predicted data showed a potential target sequence for miR-362-5p within XIST (Fig. 3A). To validate this, XIST luciferase reporter plasmid or its mutant in the seed region was introduced into 293T cells, together with miR-362-5p mimic or miR-NC mimic. With the wild-type plasmid and miR-362-5p upregulation induced a striking downregulation in luciferase activity (Fig. 3B). Upon the transfection of the site-directed mutant, little change in luciferase was observed in the presence of miR-362-5p mimic (Fig. 3B). Moreover, RT-qPCR data revealed that in comparison to the negative group, miR-362-5p level was prominently decreased by XIST overexpression in the two BC cells (Fig. 3C).
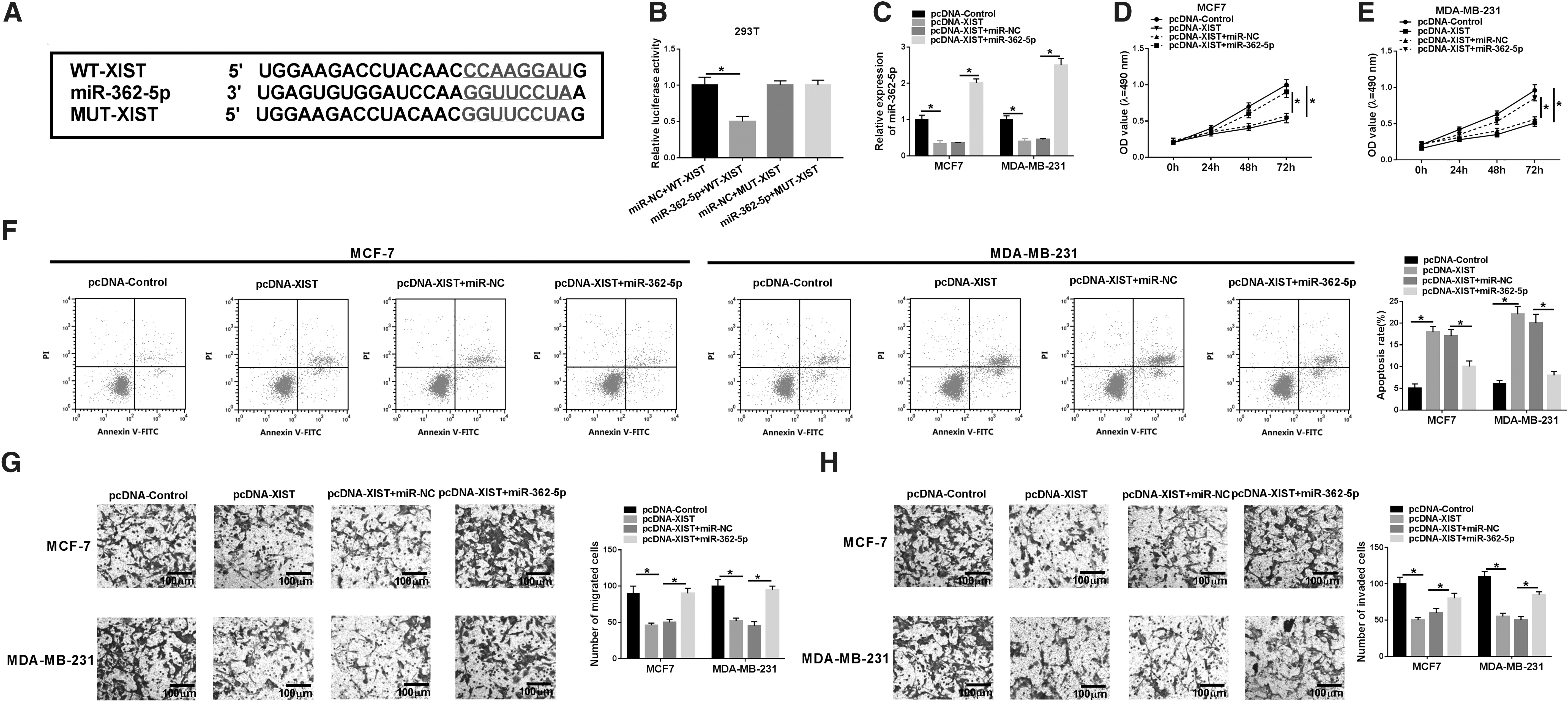
Overexpression of XIST exerted its regulatory effect on BC cell proliferation, migration, invasion, and apoptosis by miR-362-5p.
Then, they examined whether miR-362-5p functioned as a potential molecular mediator of XIST overexpression on BC cell progression. As shown by RT-qPCR, the cotransfection of miR-362-5p, but not a scrambled negative sequence, evidently reversed the reduction of XIST overexpression on miR-362-5p expression (Fig. 3C). Functional experiments data revealed that pcDNA-XIST-mediated antiproliferation (Fig. 3D, E) and proapoptosis (Fig. 3F) effects were highly abated by the restored expression of miR-362-5p. In addition, pcDNA-XIST-mediated repression in cell migration (Fig. 3G), and invasion (Fig. 3H) was significantly abolished by miR-362-5p expression restoration.
UBAP1 was a direct target of miR-362-5p and the regulatory effects of miR-362-5p depletion on BC cell malignant behaviors were mediated by UBAP1
Using the software starBase v.3.0, a predicted miR-362-5p-binding sequence was identified within UBAP1 3′-UTR (Fig. 4A). Cotransfection of UBAP1 3′-UTR luciferase reporter construct and miR-362-5p mimic in 293T cells induced a significant downregulation in luciferase activity (Fig. 4B). To verify whether the miR-362-5p-binding sites were required for this effect, the authors performed the site-directed mutagenesis to generate a UBAP1 3′-UTR mutant-type reporter construct, in which the target sequence was mutated. Notably, this mutant no longer elicited such an effect (Fig. 4B). In addition, their data showed that UBAP1 expression was substantially increased by the introduction of miR-362-5p inhibitor (Fig. 4C–F).
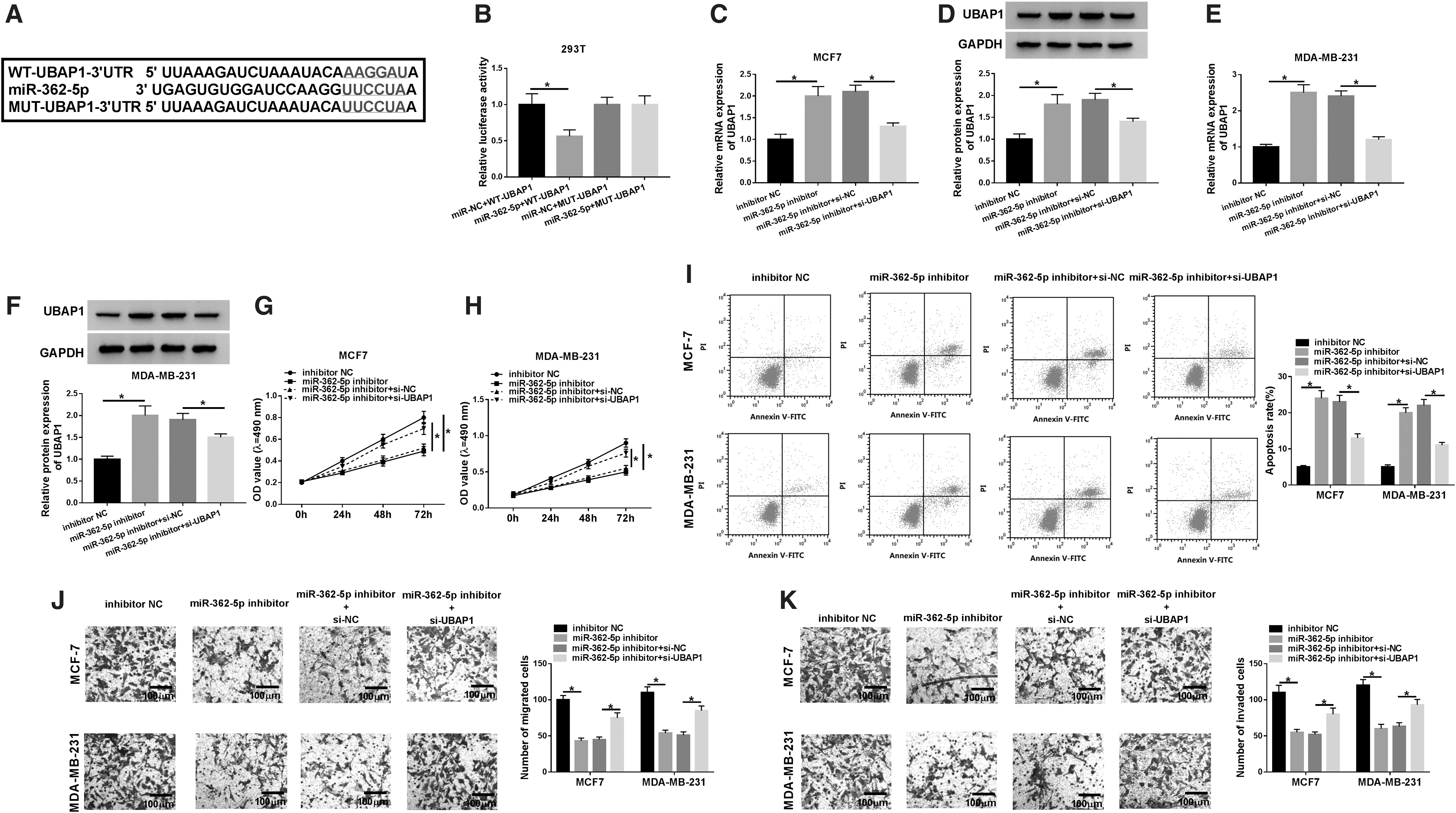
Knockdown of miR-362-5p mitigated BC cell proliferation, migration, invasion, and enhanced cell apoptosis by targeting UBAP1.
Then, MCF7 and MDA-MB-231 cells were cotransfected with miR-362-5p inhibitor and si-UBAP1. As shown in Figure 4C–F, miR-362-5p inhibitor-mediated UBAP1 augment was remarkably abated by the cotransfection of si-UBAP1 in the two cells compared with negative group. Subsequent experiment results revealed that miR-362-5p depletion exerted antiproliferation (Fig. 4G, H), proapoptosis (Fig. 4I), antimigration (Fig. 4J), and anti-invasion (Fig. 4K) effects in the two cells. However, these effects were significantly abrogated by the restored expression of UBAP1 (Fig. 4G–K).
XIST regulated UBAP1 expression via sequestering miR-362-5p
Next, the authors further investigated, if so, how XIST modulated UBAP1 expression in BC cells. As expected, in contrast to the negative control, the mRNA (Fig. 5A) and protein (Fig. 5B, C) levels of UBAP1 were strikingly increased by XIST overexpression in the two cells. However, this effect was significantly abolished by the introduction of miR-362-5p mimic (Fig. 5A–C).
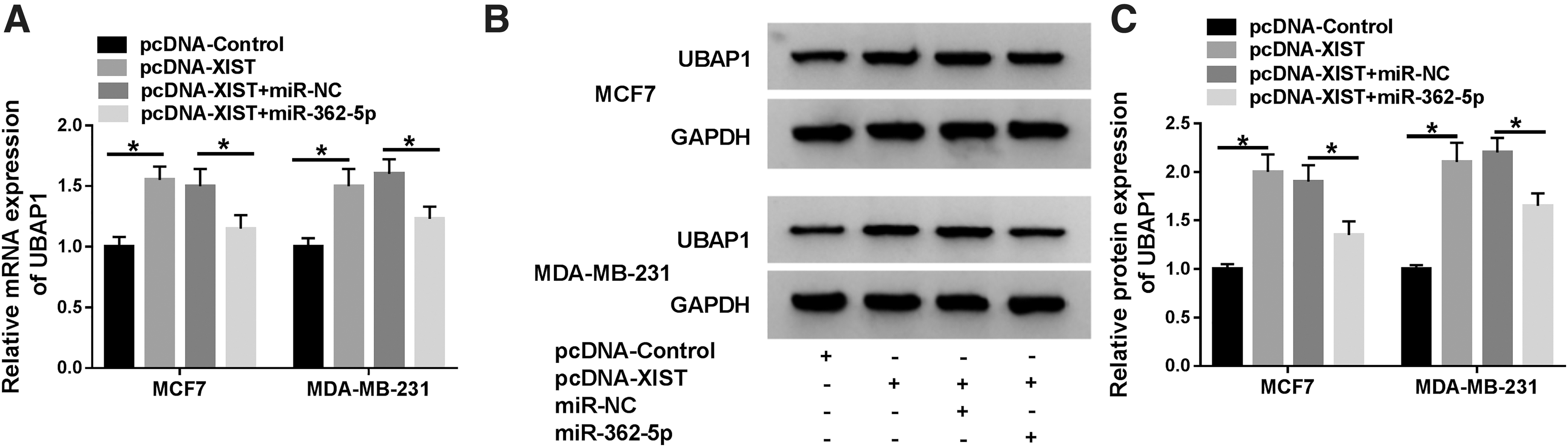
XIST modulated UBAP1 expression by sponging miR-362-5p. MCF7 and MDA-MB-231 cells were transfected with pcDNA-control, pcDNA-XIST, pcDNA-XIST+miR-NC mimic, or pcDNA-XIST+miR-362-5p mimic for 48 h.
XIST overexpression-mediated regulatory effects on BC cell malignant behaviors were reversed by the restored expression of UBAP1
Then, it was further explored whether XIST exerted its regulation on BC cell progression mediated by UBAP1. Si-UBAP1 was transfected into the two BC cells together with pcDNA-XIST. In comparison to the negative group, pcDNA-XIST-mediated UBAP1 increase was remarkably abated by the cotransfection of si-UBAP1 (Fig. 6A–D). Moreover, pcDNA-XIST-mediated antiproliferation (Fig. 6E, F), proapoptosis (Fig. 6G), antimigration (Fig. 6H), and anti-invasion (Fig. 6I) effects were drastically abolished by UBAP1 expression restoration in the two cells.

Upregulation of XIST inhibited BC cell proliferation, migration, invasion, and accelerated cell apoptosis by UBAP1. MCF7 and MDA-MB-231 were transfected with pcDNA-control, pcDNA-XIST, pcDNA-XIST+si-NC, or pcDNA-XIST+si-UBAP1.
Upregulation of XIST retarded tumor growth in vivo
Finally, the authors further observed the impact of XIST in vivo using the xenograft model. In comparison to the negative control, the transduction of lenti-XIST prominently hindered tumor cell growth in vivo (Fig. 7A, B). Moreover, XIST expression was prominently elevated (Fig. 7C) and miR-362-5p expression was remarkably reduced (Fig. 7D) in the tumor tissues derived from lenti-XIST-transducing MDA-MB-231 cells. In addition, the mRNA (Fig. 7E) and protein (Fig. 7F) levels of UBAP1 were strongly elevated by XIST overexpression in the excised tumors.
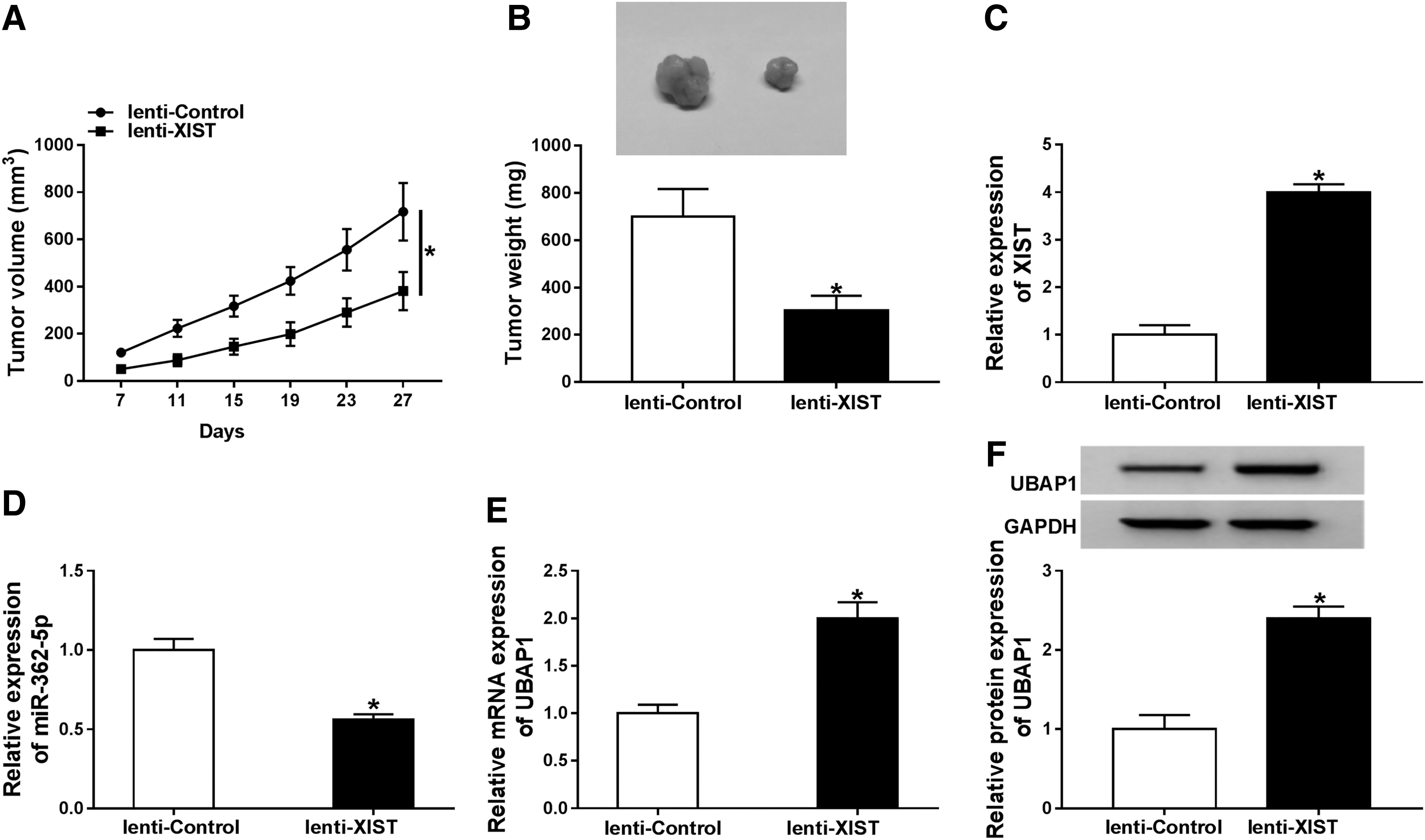
XIST overexpression inhibited tumor growth in vivo. About 5.0 × 106 lenti-XIST-transducing MDA-MB-231 cells were subcutaneously injected into the nude mice, with lenti-NC-transducing cells as the negative control (10 mice per group). At the end, all mice were sacrificed and tumor tissues were excised.
Discussion
LncRNAs are being discovered to have relevance to human cancer, contributing to crucial tumor cell functions, including proliferation, metastasis, and apoptosis. LncRNAs participate in the tumorigenesis and progression of human cancer via both oncogenic and tumor-suppressive pathways. 7 The current work demonstrated that the high level of XIST repressed BC cell progression in vitro and in vivo possibly through acting as a ceRNA of miR-362-5p and modulating UBAP1 expression.
Dysregulation of XIST has been demonstrated in many human cancers. For example, Wei et al. reported that XIST expression was increased in pancreatic cancer (PC), and XIST overexpression enhanced the proliferation of PC cells by sponging miR-133a and modulating EGFR expression. 15 Zhu et al. manifested that XIST silencing inhibited cervical cancer cell malignant behaviors through acting as a molecular sponge of miR-200a and regulated fused in sarcoma (Fus) expression. 16 Fang et al. validated that XIST depletion exerted a potential tumor suppressive effect in nonsmall cell lung cancer. 17 Former evidence had demonstrated that the low high of XIST was involved in BC progression and brain metastasis. 11,18,19 In this research, the results supported that XIST expression was significantly decreased in BC, and XIST overexpression suppressed BC cell proliferation, migration, invasion, and enhanced cell apoptosis, consistent with a recent document. 12 In short, XIST protected against BC malignant progression, in line with earlier reports. 11,12,18
Then, they used the online software starBase v.3.0 to help identify the potentially interacted miRNAs of XIST. MiR-362-5p was fascinating in this research because miR-362-5p upregulation had been found in triple-negative BC tissues. 13 Previous investigations had reported the dysregulation of miR-362-5p in a multitude of human cancers. For instance, Ni et al. found a significant upregulation of miR-362-5p in hepatocellular carcinoma (HCC) tissues, and miR-362-5p depletion hampered HCC cell proliferation and metastasis. 20 Yang et al. manifested that miR-362-5p silencing retarded tumor growth of chronic myelocytic leukemia cells by targeting DNA damage-inducible 45α (GADD45α). 21 Wu et al. demonstrated that the high level of miR-362-5p hindered the malignant behaviors of the metastatic neuroblastoma cells. 22 These findings from the study described above suggested that miR-362-5p functioned as an onco-miRNA or tumor suppressor in some human cancers. In this study, the authors for the first time confirmed that XIST directly interacted with miR-362-5p. They also validated that miR-362-5p knockdown attenuated BC cell progression in vitro, in line with a previous report. 14 More importantly, it was uncovered that miR-362-5p mediated the regulation of XIST overexpression on BC cell malignant behaviors.
MiRNAs are widely known to function as key players in biological processes through transcriptional repression of target genes expression. Among the ∼2940 targets of miR-362-5p, UBAP1 was selected for further research owing to its origin from the tumor-suppressive locus in chromosome. 23 The findings by Qian et al. had also demonstrated that UBAP1 gene exists at human chromosome undergoing loss of heterozygosity in nasopharyngeal carcinoma (NPC). 23 Xiao et al. manifested that UBAP1 was downregulated in NPC tissues, and the low UBAP1 expression might drive NPC malignant transformation. 24 In addition, an earlier document demonstrated that UBAP1 downregulation was discovered in many neoplastic tissues. 25 In the present research, the data revealed a significant downregulation of UBAP1 in BC tissues and cells, eliciting a potential involvement of UBAP1 in BC progression. The authors were first to uncover that UBAP1 was a direct functional target of miR-362-5p in BC cells. Moreover, the regulatory effects of miR-362-5p depletion on BC cell proliferation, migration, invasion, and apoptosis were mediated by UBAP1. Similar to their findings, previous reports validated that miR-362-5p modulated tumor progression via targeting several other genes, such as CYLD lysine 63 deubiquitinase (CYLD) and phosphatidylinositol 3-kinase-C2β. 20,22
It is widely accepted that lncRNA functions as a ceRNA of specific miRNA to regulate gene expression. Therefore, the authors further investigated whether UBAP1 was involved in the regulation of the XIST/miR-362-5p axis on BC progression. As expected, their data illuminated that XIST regulated UBAP1 expression by sponging miR-362-5p, and XIST overexpression inhibited BC cell proliferation, migration, invasion, and accelerated cell apoptosis by UBAP1. Zheng et al. reported that XIST overexpression repressed BC cell malignant behaviors by acting as a ceRNA of miR-155 and modulating CDX1 expression. 12
Finally, the xenograft assays demonstrated that the high level of XIST hampered tumor growth in vivo. Furthermore, the upregulation of XIST resulted in decreased miR-362-5p expression and increased UBAP1 level in the xenograft tumor. These data suggested that XIST overexpression repressed tumor growth in vivo possibly via the miR-362-5p/UBAP1 axis. More researches about the role and molecular mechanism of UBAP1 on BC progression will be carried out in further work.
Conclusion
In conclusion, the authors' research indicated that the high level of XIST suppressed BC cell progression in vitro and in vivo at least partially by acting as a ceRNA of miR-362-5p and modulating UBAP1 expression. Their research provided new evidence for XIST as a promising therapeutic agent for BC management.
Footnotes
Authors' Contributions
Conception and design: Baoyin Liu and Chengyu Luo; Development of methodology: Hua Lin; Acquisition of data: Xiaoxin Ji; Analysis and interpretation of data: Xin Li; Writing, review, and revision of article: Baoyin Liu and Chengyu Luo; All coauthors have reviewed and approved of the article before submission.
Disclosure Statement
The authors declare that they have no financial conflicts of interest.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.