
Abstract
Background:
Osteosarcoma (OS) is a primary malignant tumor in children and adolescents. Long noncoding RNA HNF1A antisense RNA 1 (HNF1A-AS1) is connected with OS development. However, there are few reports on the role and mechanism of HNF1A-AS1 in OS.
Materials and Methods:
Quantitative real-time polymerase chain reaction (qRT-PCR) was employed to assess the expression of HNF1A-AS1, miR-32-5p, and high-mobility group protein B1 (HMGB1). Western blot analysis was performed to detect the protein level of HMGB1. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), colony formation, transwell, or flow cytometer assays were applied to determine the proliferation, migration, invasion, and apoptosis of OS cells. The interaction between HNF1A-AS1 and miR-32-5p or HMGB1 was predicted by the starBase database and confirmed by dual-luciferase reporter assay. Enzyme-linked immunosorbent assay was employed to analyze levels of HMGB1 in the OS cell supernatant.
Results:
HNF1A-AS1 and HMGB1 were upregulated, while miR-32-5p was downregulated, in OS tissues and cells. Functionally, HNF1A-AS1 depletion induced apoptosis and impeded proliferation, migration, and invasion of OS cells. Interestingly, HNF1A-AS1 bound to miR-32-5p to regulate the expression of HMGB1. Furthermore, miR-32-5p knockdown overturned the effects of HNF1A-AS1 knockdown on apoptosis, proliferation, migration, and invasion of OS cells. In addition, the effects of HNF1A-AS1 silencing on the malignant behaviors of OS cells were reserved by HMGB1 overexpression. In addition, HNF1A-AS1 regulated the HMGB1 level in the OS cell supernatant through the miR-32-5p/HMGB1 axis.
Conclusion:
Downregulation of HNF1A-AS1 blocked OS progression through the miR-32-5p/HMGB1 axis, which provides a possible target and prognostic biomarker for treatment of OS.
Introduction
Osteosarcoma (OS) occurs mainly in the vicinity of the shoulder joint and the long metaphysis (humerus, proximal humerus, and distal femur) and is a universal elementary malignancy found in children and adolescents. 1,2 At the present day, the treatment of OS is mainly based on surgery combined with multiagent chemotherapy. 3 Moreover, patients with recurrence and lung metastases account for about 30%–40% of OS patients and their overall 5-year survival rate is about 20%. 4 Patients with advanced or relapsed OS have a poor prognosis. 5 Consequently, there is an urgent need to further explore new mechanisms to introduce new methods for treatment of OS to enhance the prognosis of OS patients.
Long non-coding RNAs (lncRNAs) lack the potential for protein coding and have functionally structured RNA domains, picturesque regulatory mechanisms, cis-regulated activity, and alternative forms of biogenesis. 6 The literature indicates that lncRNAs that have been identified can serve as potential therapeutic targets or biomarkers for multiple cancers. 7 lncRNA HNF1A antisense RNA 1 (HNF1A-AS1) is connected with development of a range of cancers, such as hepatocellular cancer, 8 colon cancer, 9 and esophageal squamous cell cancer. 10 Furthermore, it has been revealed in the literature that HNF1A-AS1 is connected with the occurrence of OS. 11 At present, there are few reports on the role and mechanism of HNF1A-AS1 in OS.
MicroRNAs (miRNAs), a large family of post-transcriptional regulatory genes (∼21 nucleotides), play an elementary role in viral infection, cell metabolism and differentiation, organismal development, and tumorigenesis. 12,13 Moreover, studies have shown that miRNAs can serve as potential therapeutic targets and biomarkers for some diseases. 14,15 It is reported that miRNA-32-5p (miR-32-5p) is associated with the inflammatory response, drug resistance, and tumor proliferation. 16 –19 At the moment, the role of miR-32-5p and its related molecular mechanisms are rarely reported in OS.
High-mobility group protein B1 (HMGB1), a nonhistone that binds primarily to nuclear DNA, can be released extracellularly and is connected with the inflammatory immune response as well as tumor metastasis, invasion, and growth. 20,21 In addition, it has a promoting effect in diverse malignant tumors such as nonsmall cell lung cancer, 22 hepatocellular cancer, 23 and gastric cancer. 24 Studies have shown that HMGB1 can enhance the drug resistance of OS, and high HMGB1 expression indicates that the prognosis of patients with OS is poor. 25,26 However, it is unclear whether HMGB1 is regulated by HNF1A-AS1 and miR-32-5p in OS.
Hence, the expression patterns of HNF1A-AS1 in OS were investigated. Moreover, the effect of HNF1A-AS1 on OS cells was explored. Besides, whether the HNF1A-AS1-miR-32-5p-HMGB1 axis regulates OS progression was studied. This research sheds light on a possible biomarker and therapeutic target for the diagnosis and treatment of OS.
Materials and Methods
OS patient specimens
This study was ratified and supervised by the ethics committee of Jingmen No. 1 People's Hospital. OS tumor tissues and neighboring nontumor tissues were gained from 68 patients diagnosed with OS at Jingmen No. 1 People's Hospital. All OS patients participating in the study did not receive chemotherapy or radiotherapy before undergoing surgery and signed an informed consent.
Quantitative real-time polymerase chain reaction
An RNA extraction kit from Invitrogen (Carlsbad, CA) was employed to extract total RNA for detection of HNF1A-AS1, miR-32-5p, and HMGB1 levels in OS tumor tissues and neighboring nontumor tissues as well as OS cells. The complementary DNA of HNF1A-AS1, miR-32-5p, and HMGB1 was synthesized with the PrimeScript RT reagent kit from TIANGEN (Beijing, China). In addition, the mRNA levels of HNF1A-AS1, miR-32-5p, and HMGB1 were evaluated through the SYBR Premix Ex Taq™ II kit from Takara (Dalian, China). Using the 2−ΔΔCt method, the expression levels of HNF1A-AS1, miR-32-5p, and HMGB1 were normalized to the expression of U6 small nuclear RNA (snRNA) or glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Primers were displayed as follows: GAPDH (F: 5′-GAAGGTGAAGGTCGGAGTC-3′, R: 5′-GAAGATGGTGATGGGATTTC-3′), HNF1A-AS1 (F: 5′-TCAAGAAATGGTGGCTAT-3′, R: 5′-GCTCTGAGACTGGCTGAA-3′), miR-32-5p (F: 5′-CGGTATTGCACATTACTAAGTTGCA-3′, R: 5′-CTCGCTTCGGCAGCACA-3′), HMGB1 (F: 5′-AGCTGCTAGCGCCTAGCGAT-3′, R: 5′-CCCGTCTGATAGCGCATTCGTGT-3′), and U6 snRNA (F: 5′-GCTCGCTTCGGCAGCACA-3′, R: 5′-GAGGTATTCGCACCAGAGGA-3′).
Cell culture and transfection
Five OS cell lines (U2OS, Saos-2, 143B, HOS, and MG63) and the hFOB1.19 cells were received from American Type Culture Collection (Manassas, VA). HOS and MG63 cells were cultivated using Eagle's minimum essential medium (MEM) (Gibco, Invitrogen). Ham's F12 medium (F12) (Gibco) was employed for the maintenance of hFOB1.19 cells. In addition, U2OS, Saos-2, and 143B cells were cultivated by using Dulbecco's modified Eagle's medium (DMEM) (Gibco). The above media contained fetal bovine serum (10%, FBS; Life Technologies, Carlsbad, CA), penicillin (1%; Baomanbio, Shanghai, China), and streptomycin (1%; Baomanbio). All of the above cell lines were cultivated in an incubator with 5% CO2 at 37°C constant temperature.
The pcDNA3.1 vector (Invitrogen) was employed to construct the pcDNA-HNF1A-AS1 overexpression vector (HNF1A-AS1) and pcDNA-HMGB1 overexpression vector (HMGB1). Small hairpin negative control (sh-NC) and HNF1A-AS1 small hairpin (sh-HNF1A-AS1) were obtained from GenePharma (Shanghai, China). miRNA mimic targeting miR-32-5p (miR-32-5p), miRNA inhibitor targeting miR-32-5p (anti-miR-32-5p), scrambled mimic control (miR-NC), and miR-32-5p inhibitor negative control (anti-miR-NC) were obtained from RiboBio (Guangzhou, China). When the cell density reached 70%–90%, Lipofectamine 2000 reagent (Invitrogen) was employed for transfection of U2OS or Saos-2 cells.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was applied to assess cell proliferation. In short, transfected OS cells (5 × 103 cells/well) were cultured in 96-well plates (Corning Costar, Corning, NY) at 37°C constant temperature with 5% CO2 for 24, 48, and 72 h. Following this, MTT (20 μL; Sigma) was added to each well and maintained for 4 h. Next, the supernatant of each well was removed and 150 μL of dimethyl sulfoxide was replenished for the dissolution of formazan crystals. The microplate absorbance reader from Thermo Fisher Scientific (Waltham, MA) was applied to evaluate the color reaction at 490 nm.
Cell colony formation assay
Transfected OS cells (5 × 103 cells/well) were seeded into a six-well plate containing complete medium (3 mL) and the medium was replaced every 2 days. After culture for 10 days, the cells were fixed with cold methanol and then stained with crystal violet (0.1%). Following this, the cells were washed with phosphate-buffered saline, and then the colonies (≥50 cells) were figured and imaged under a microscope (Olympus, Tokyo, Japan).
Flow cytometer assay
The Annexin V-FITC–propidium iodide (PI) kit (BD Biosciences, San Diego, CA) was employed to detect the apoptosis of transfected OS cells. In short, cells transfected for 48 h were harvested and washed with cold phosphate-buffered saline and then resuspended in binding buffer. Following this, Annexin V-FITC and PI were added to the binding solution with suspended cells. The apoptotic rate of transfected OS cells was examined using FACScan® flow cytometry (BD Biosciences).
Transwell assay
The migration and invasion of transfected OS cells were assessed using the transwell chamber (8 μm pore filter; BD Biosciences). For the invasion assay, the Matrigel matrix (BD Biosciences) was applied and placed in the upper chamber. Briefly, FBS (10%) was added as a chemoattractant to DMEM and placed in the lower chamber. Contemporaneously, transfected OS cells (5 × 104) were added to DMEM with 1% FBS in the upper chamber. After 24 h of cultivation, methanol (100%) was employed for fixation of the migrated or invaded cells in the lower surface of the filter membrane. Following this, 0.1% crystal violet was applied to stain migrated or invaded OS cells. In the end, an inverted microscope (Olympus) was used to figure the migrated and invaded OS cells.
Dual-luciferase reporter assay
The starBase2.0 database was employed for prediction of the binding sites between HNF1A-AS1 or HMGB1 and miR-32-5p. Then, wild-type HMGB1 (HMGB1 WT 3′UTR), mutant HMGB1 (HMGB1 MUT 3′UTR), wild-type HNF1A-AS1 (HNF1A-AS1 WT), and mutant HNF1A-AS1 (HNF1A-AS1 MUT) sequences that contain putative miR-32-5p binding sites were synthesized. These sequences were then separately inserted into the pGL3-control luciferase reporter vector (Promega) to construct the luciferase reporter vector. Following this, the luciferase reporter vector or control vector was cotransfected with miR-32-5p or miR-NC for U2OS and Saos-2 cells, respectively. At last, the luciferase activities of luciferase reporter vectors were assessed through the luciferase reporter assay kit (Promega).
Western blot analysis
The whole protein of OS tissues and adjacent normal bone as well as OS cells was extracted through RIPA lysis buffer (Beyotime, Shanghai, China). Then, the whole protein was assessed with the BCA assay kit from Pierce (Rockford, IL), followed by separation on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and transferred to polyvinylidene difluoride (PVDF) membranes (Millipore, Billerica, MA). Subsequently, the PVDF membranes were blocked through Tris-buffered saline-Tween buffer with 5% skim milk for 1 h. Then, PVDF membranes were incubated with primary antibodies overnight at 4°C.
Following this, the membranes were then washed and incubated with goat anti-mouse IgG (Abcam, Cambridge, MA). The EasySee Western blot kit from Transgen Biotech (Beijing, China) was utilized to detect protein bands. Quantity One software from Bio-Rad Laboratories (Hercules, CA) was used for assessment of protein bands. Primary antibodies in this study were purchased from Abcam, including anti-HMGB1 (1:1000) and anti-β-actin (1:1000), and β-actin was regarded as a loading control.
Enzyme-linked immunosorbent assay
The levels of HMGB1 in the supernatant of OS cells were determined with the enzyme-linked immunosorbent assay (IBL, Toronto, Canada) based on the manufacturer's instructions.
Statistical analysis
Differences with p < 0.05 were considered statistically significant. Student's t-test or one-way ANOVA was performed to compare differences between groups. Data are displayed as mean ± standard deviation. The Kaplan–Meier method was employed to construct the survival curves of OS patients with high or low HNF1A-AS1 expression. The correlation between HNF1A-AS1, miR-32-5p, and HMGB1 was analyzed using Pearson's correlation coefficient. SPSS 18.0 software (SPSS, Chicago, IL) was employed to perform the statistical analysis.
Results
HNF1A-AS1 was upregulated in OS tissues and cell lines
To explore the role of HNF1A-AS1 in OS, the authors detected the expression pattern of HNF1A-AS1 in 68 patients with OS. Results of quantitative real-time polymerase chain reaction (qRT-PCR) exhibited an evident augmentation of HNF1A-AS1 in OS tissues compared with neighboring nontumor tissues (Fig. 1A). Besides, the authors further assessed the correlation of the HNF1-AS1 expression level with the clinical stage and lung metastases. Results revealed that HNF1A-AS1 was strikingly higher in clinical II and III stages than in the clinical I stage (Fig. 1B). Moreover, an enhanced expression level of HNF1A-AS1 was discovered in lung metastases compared with nonlung metastasis OS tissues (Fig. 1C).
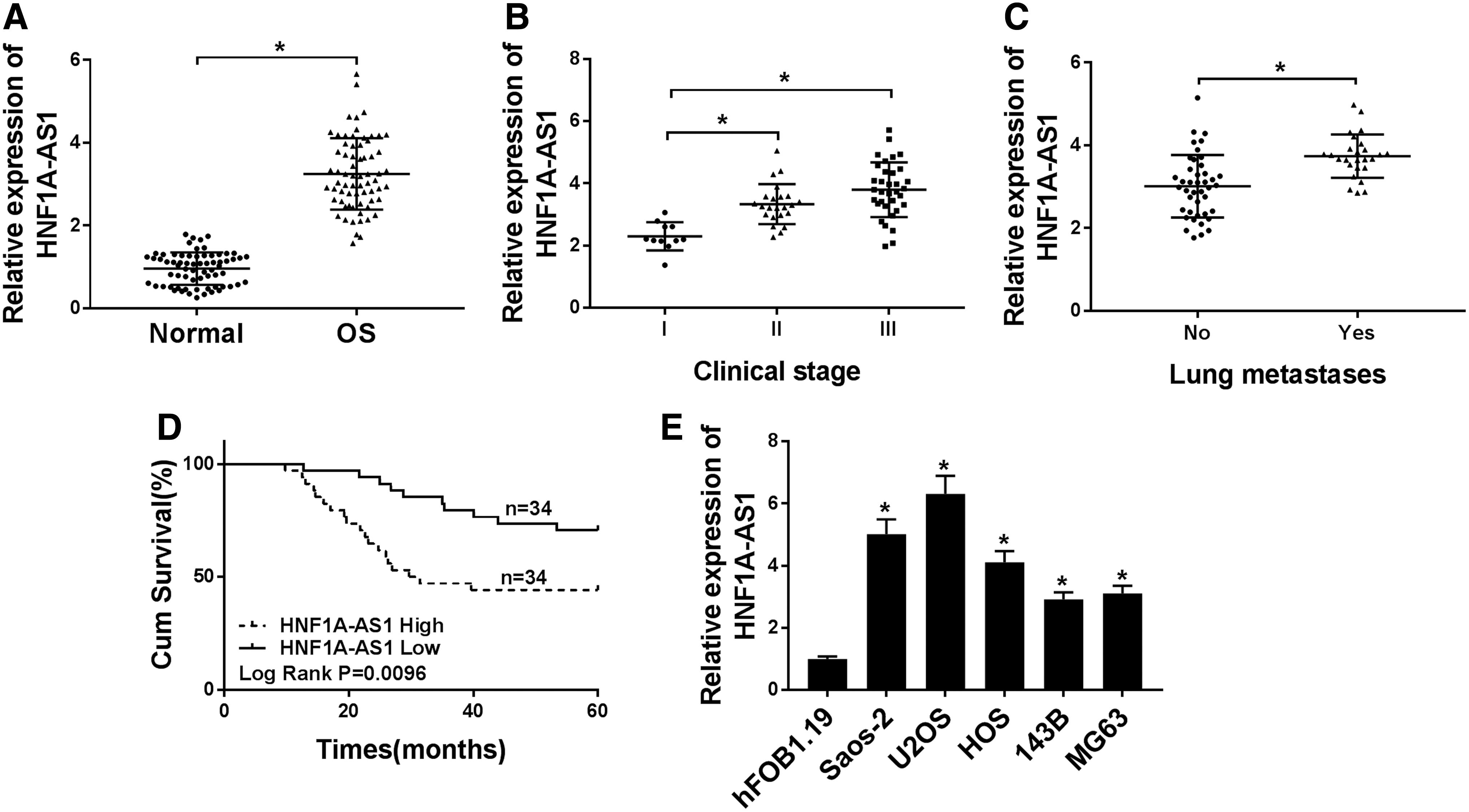
HNF1A-AS1 was upregulated in OS tissues and cell lines.
Furthermore, OS patients were divided into two groups based on HNF1A-AS1 median expression in OS tissues: high HNF1A-AS1 expression group (>median, N = 34) and low HNF1A-AS1 expression group (<median, N = 34). The Kaplan–Meier analysis was performed and results revealed that patients with high HNF1A-AS1 expression (n = 34) in OS have a worse survival rate compared with that of OS patients with low HNF1A-AS1 expression (n = 34) (Fig. 1D).
In addition, the expression level of HNF1A-AS1 in OS cell lines (Saos-2, U2OS, HOS, 143B, and MG63) and hFOB1.19 cells was measured using qRT-PCR. As exhibited in Figure 1E, a remarkably higher HNF1A-AS1 expression was observed in OS cells versus hFOB1.19 cells. Therefore, HNF1A-AS1 was upregulated in OS tissues and cell lines, and high HNF1A-AS1 was associated with the clinical stage, lung metastases, and overall survival.
Silencing of HNF1A-AS1 expedited apoptosis and restrained proliferation, migration, and invasion of OS cells
Based on the above results, the authors selected Saos-2 and U2OS cells for further exploration of the role of HNF1A-AS1 in OS. In the first place, sh-HNF1A-AS1 or sh-NC was transfected into U2OS and Saos-2 cells to silence the expression of HNF1A-AS1. As displayed in Figure 2A, the expression of HNF1A-AS1 was conspicuously decreased in both U2OS and Saos-2 cells transfected with sh-HNF1A-AS1 compared with the control group, indicating that U2OS and Saos-2 cells have good transfection efficiency. Furthermore, the effects of HNF1A-AS1 silencing on cell proliferation, apoptosis, migration, and invasion were further investigated in OS cells. MTT assay was employed to assess the effect of HNF1A-AS1 knockdown on OS cell proliferation. The results revealed that proliferation of U2OS and Saos-2 cells was markedly impeded by HNF1A-AS1 knockdown compared with the control group (Fig. 2B, C).
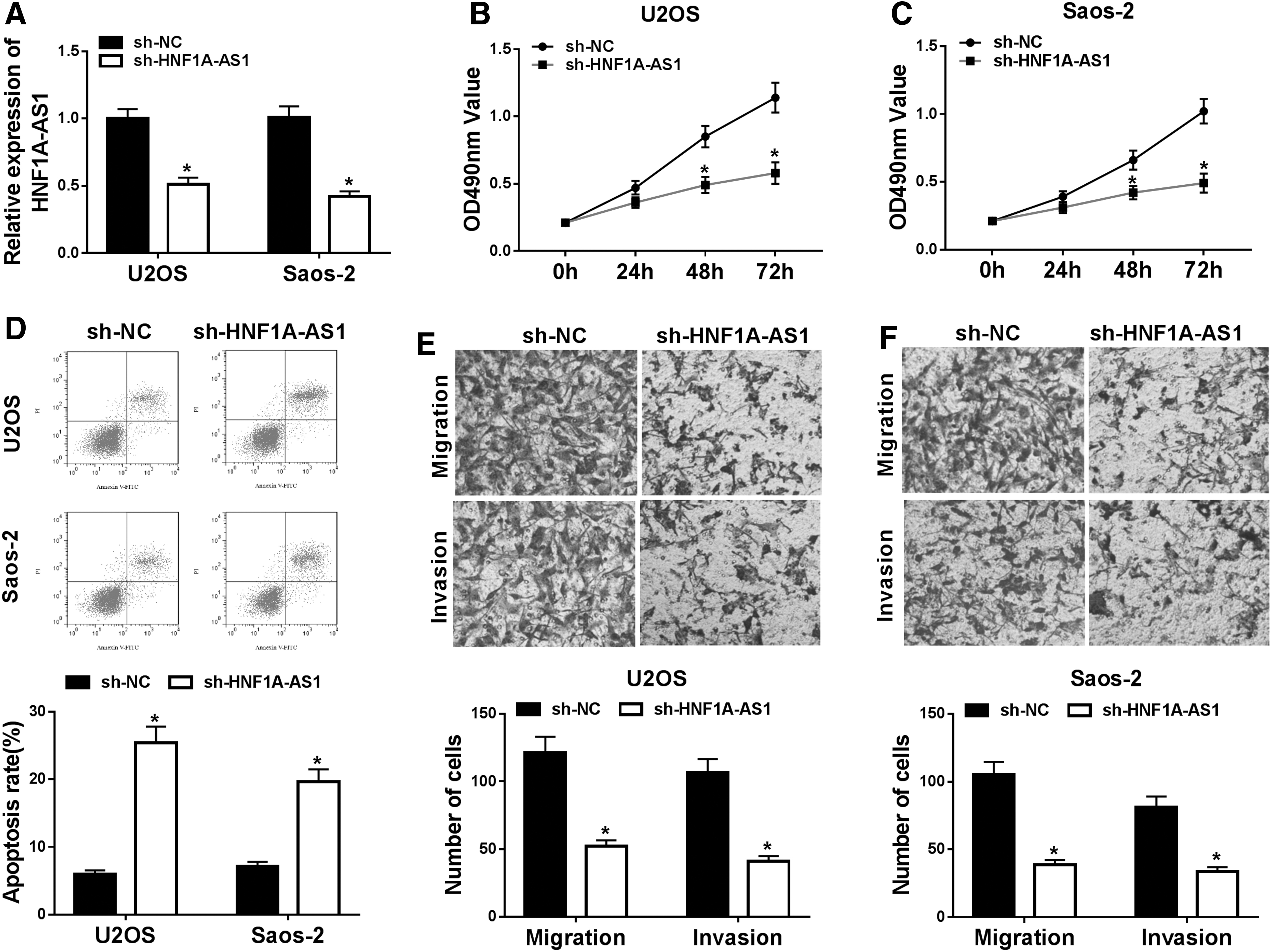
HNF1A-AS1 knockdown restrained proliferation, migration, and invasion and expedited apoptosis of OS cells. Sh-HNF1A-AS1 or sh-NC was transfected into U2OS and Saos-2 cells.
Cell colony formation assay showed that reduced HNF1A-AS1 expression constrained the colony formation ability of U2OS and Saos-2 cells (Supplementary Fig. S1). Flow cytometry assay indicated that reduction of HNF1A-AS1 drastically elevated the apoptosis rate of U2OS and Saos-2 cells than that of the control group (Fig. 2D). In addition, the transwell assay was performed and results revealed that conspicuous migration and invasion inhibition of U2OS and Saos-2 cells was caused by decreased HNF1A-AS1 versus the control group (Fig. 2E, F).
HNF1A-AS1 bound to miR-32-5p and negatively regulated its expression
In view of the above results, the underlying molecular mechanisms of HNF1A-AS1 in OS were further explored. Originally, the starBase database (
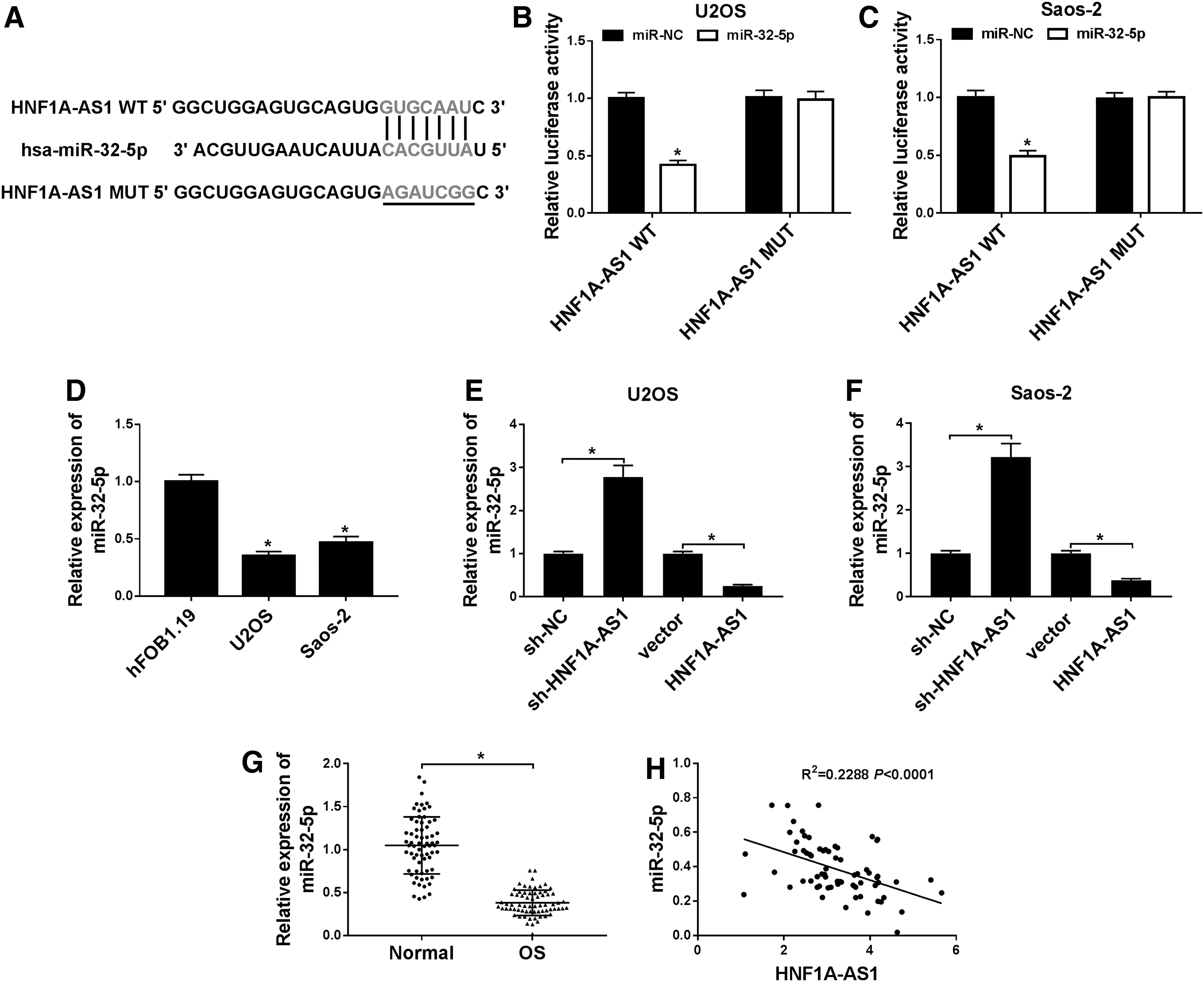
HNF1A-AS1 negatively regulated miR-32-5p expression.
Subsequently, the expression of miR-32-5p was detected with qRT-PCR in OS cells. The results showed that miR-32-5p was apparently decreased in U2OS and Saos-2 cells compared with hFOB1.19 cells (Fig. 3D). Moreover, the effect of HNF1A-AS1 on expression of miR-32-5p was analyzed by qRT-PCR. The results displayed that HNF1A-AS1 overexpression inhibited expression of miR-32-5p in U2OS and Saos-2 cells, while the decrease in HNF1A-AS1 enhanced expression of miR-32-5p (Fig. 3E, F). In addition, miR-32-5p was obviously reduced in OS tissues compared with the neighboring nontumor tissues (Fig. 3G). Furthermore, Pearson's correlation analysis revealed that HNF1A-AS1 was negatively correlated with miR-32-5p expression in OS tissues (Fig. 3H). Together, these results indicated that HNF1A-AS1 could bind to miR-32-5p and negatively regulate its expression.
MiR-32-5p reduction reversed the effect of HNF1A-AS1 knockdown on proliferation, apoptosis, migration, and invasion of OS cells
To verify whether HNF1A-AS1 is involved in the progression of OS through miR-32-5p, the authors transfected miR-NC, miR-32-5p, sh-HNF1A-AS1+anti-NC, or sh-HNF1A-AS1+anti-miR-32-5p into U2OS and Saos-2 cells to detect the expression level of miR-32-5p. Results of qRT-PCR revealed that expression of miR-32-5p was drastically increased in U2OS and Saos-2 cells transfected with miR-32-5p, and enhanced miR-32-5p expression by HNF1A-AS1 knockdown was abolished by miR-32-5p reduction (Fig. 4A, B). Following this, the authors further verified whether HNF1A-AS1 manages proliferation, apoptosis, migration, and invasion of OS cells by affecting miR-32-5p expression with MTT, transwell, and flow cytometry assays.
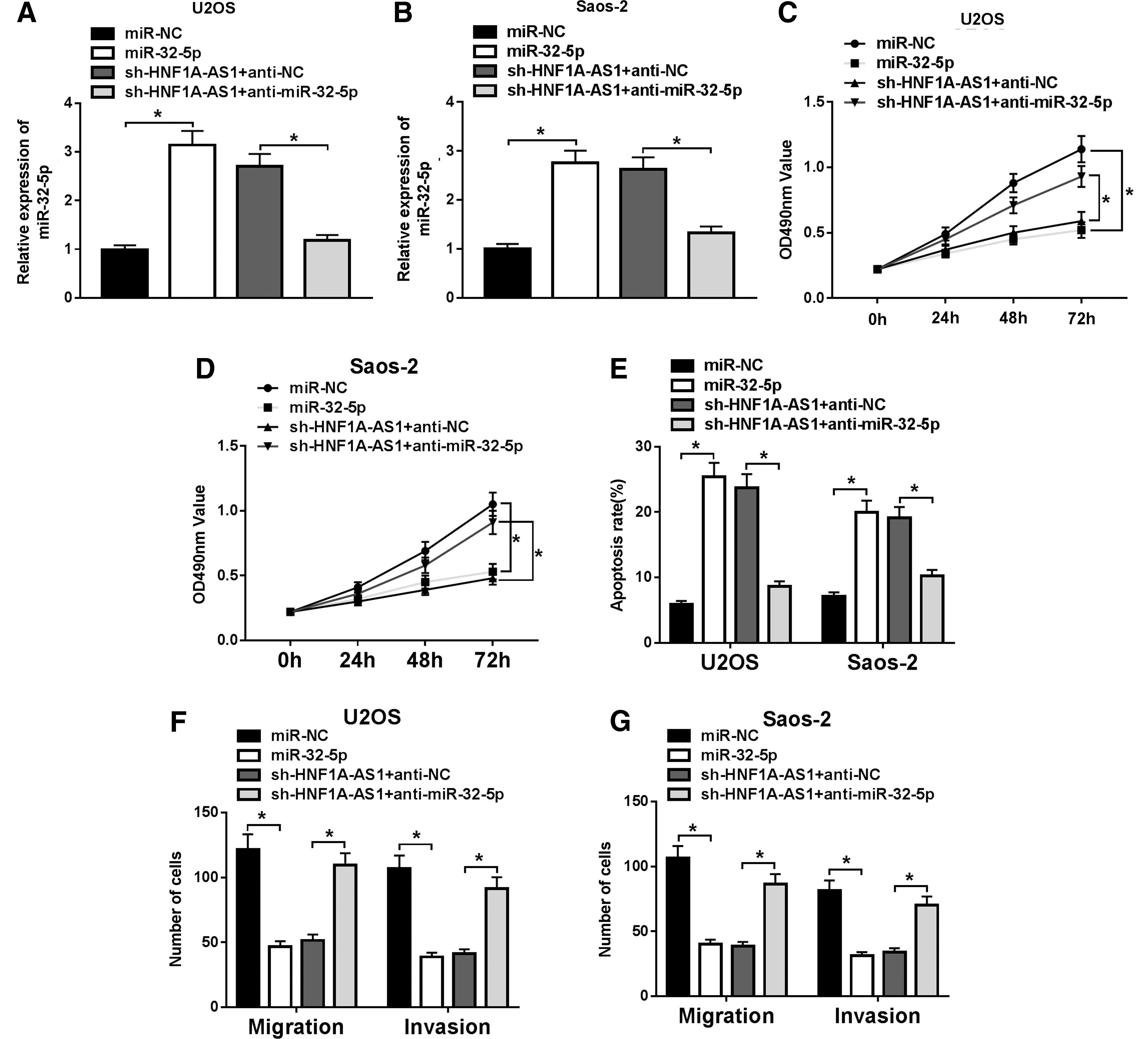
MiR-32-5p reduction undermined the effect of HNF1A-AS1 knockdown on proliferation, apoptosis, migration, and invasion of OS cells. miR-NC, miR-32-5p, sh-HNF1A-AS1+anti-NC, or sh-HNF1A-AS1+anti-miR-32-5p was transfected into U2OS and Saos-2 cells.
Results of the MTT assay showed that overexpression of miR-32-5p suppressed proliferation of U2OS and Saos-2 cells, while miR-32-5p reduction reversed the inhibitory effect of HNF1A-AS1 knockdown on proliferation of U2OS and Saos-2 cells (Fig. 4C, D). Moreover, the flow cytometry assay revealed a marked elevation of cell apoptosis induced by miR-32-5p enhancement in U2OS and Saos-2 cells, while downregulation of miR-32-5p partially abrogated the effect of knockdown of HNF1A-AS1 on apoptosis of U2OS and Saos-2 cells (Fig. 4E).
In addition, the transwell assay displayed that augmentation of miR-32-5p repressed migration and invasion of U2OS and Saos-2 cells, while reduction of miR-32-5p remarkably attenuated the inhibitory effect of HNF1A-AS1 knockdown on OS cell migration and invasion (Fig. 4F, G). These data indicated that reduction of miR-32-5p overturned the effect of HNF1A-AS1 knockdown on proliferation, apoptosis, migration, and invasion of OS cells.
HNF1A-AS1 regulated HMGB1 expression by binding to miR-32-5p
Considering that lncRNA can be used as a competing endogenous RNA (ceRNA), the authors further speculated that HNF1A-AS1 regulates the miR-32-5p targeting gene by binding to miR-32-5p and participates in progression of OS. To verify this speculation, the starBase database was employed to predict the target gene for miR-32-5p (Fig. 5A). The results presented that HMGB1 was a potential target gene of miR-32-5p. Then, the luciferase plasmids of HMGB1 WT 3′UTR and HMGB1 MUT 3′UTR were constructed and cotransfected with miR-32-5p or miR-NC into U2OS and Saos-2 cells, respectively. Luciferase reporter assay was then performed and results exhibited that the luciferase vitality of HMGB1 WT 3′UTR was apparently reduced by miR-32-5p, whereas HMGB1 MUT 3′UTR clogged this effect (Fig. 5B, C).
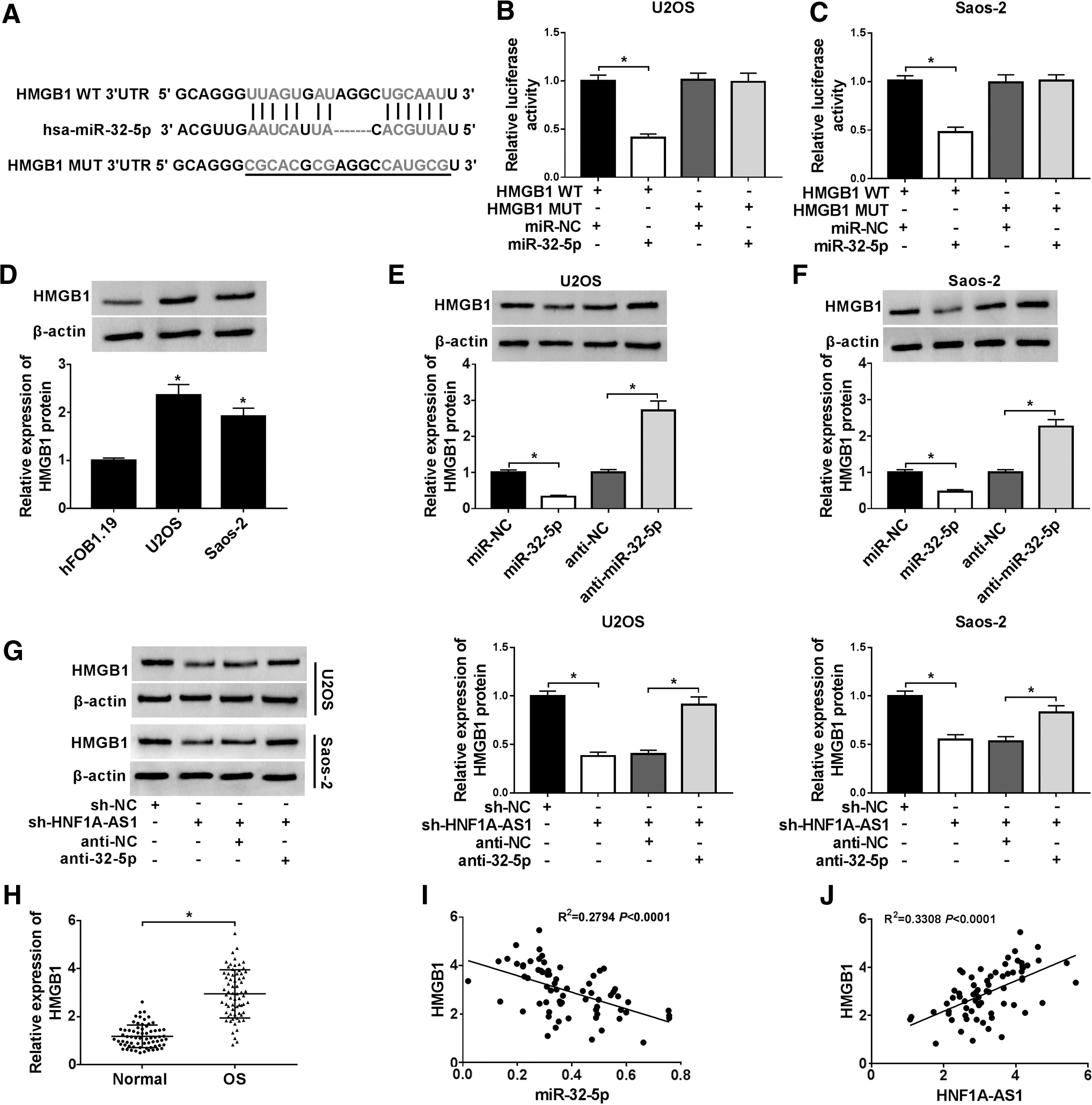
HNF1A-AS1 bound to miR-32-5p to regulate HMGB1 expression.
Subsequently, Western blot analysis was utilized to measure the protein level of HMGB1 in OS and hFOB1.19 cells. The results showed that HMGB1 protein was prominently upregulated in U2OS and Saos-2 cells compared with hFOB1.19 cells (Fig. 5D). Furthermore, the effect of miR-32-5p on expression of HMGB1 protein was assessed using Western blot analysis. The results displayed that increased miR-32-5p retarded the expression of U2OS and Saos-2 cells, while reduced miR-32-5p elevated HMGB1 protein expression (Fig. 5E, F). Besides, Western blot analysis was applied to analyze whether HNF1A-AS1 regulates HMGB1 expression through miR-32-5p. The results showed that knockdown of HNF1A-AS1 apparently downregulated HMGB1 protein expression, while this reduction was abolished by miR-32-5p silencing (Fig. 5G). It was consistent with the trend of HMGB1 protein expression in U2OS and Saos-2 cells, and expression of HMGB1 mRNA was also robustly improved in OS tissues (Fig. 5H).
In addition, the Pearson correlation analysis indicated that HMGB1 expression was negatively correlated with miR-32-5p expression and positively correlated with HNF1A-AS1 in OS tissues (Fig. 5I, J). Together, these data revealed that HNF1A-AS1 could bind to miR-32-5p to regulate HMGB1 expression in OS cells.
HMGB1 overexpression attenuated the effect of HNF1A-AS1 reduction on proliferation, apoptosis, migration, and invasion of OS cells
To explore whether HMGB1 was involved in the progression of OS regulated by HNF1A-AS1, we first examined the levels of HMGB1 protein in OS cells transfected with sh-HNF1AAS1, sh-NC, pcDNA, or pcDNA-HMGB1. Results of the Western blot analysis exhibited that the protein level of HMGB1 was remarkably increased in U2OS and Saos-2 cells transfected with HMGB1 compared with the control group. Western blot analysis was then employed to measure the level of HMGB1 protein in response to HNF1A-AS1 silencing and the results displayed that a decrease of HNF1A-AS1 reduced the level of HMGB1, indicating that HMGB1 was connected with the role exerted by HNF1A-AS1 (Fig. 6A).
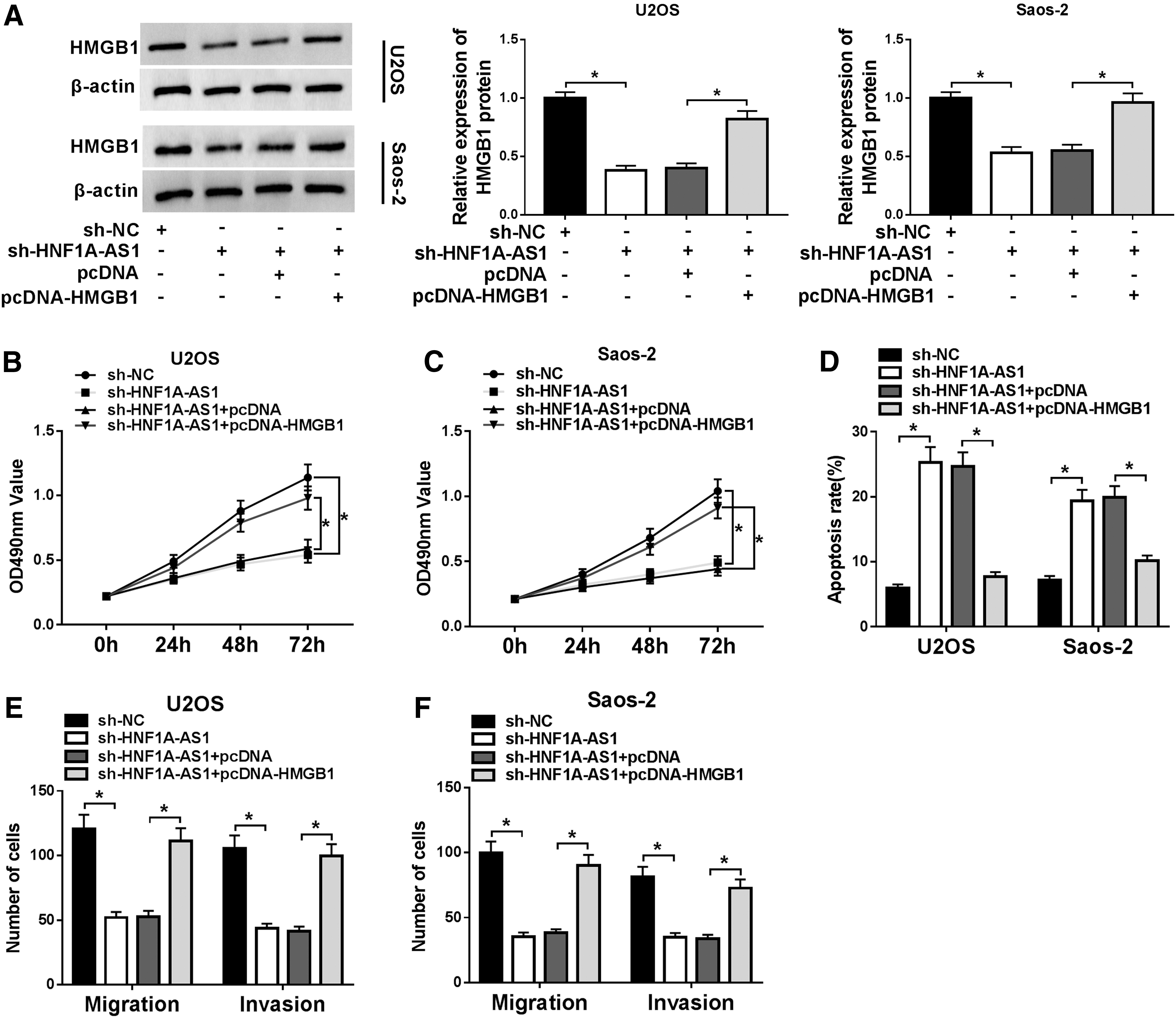
HMGB1 was related to the effect of HNF1A-AS1 on proliferation, apoptosis, migration, and invasion of OS cells.
Subsequently, the HMGB1 expression vector or empty vector was transfected into U2OS and Saos-2 cells transfected with sh-HNF1A-AS1 to perform the rescue experiments. The MTT assay presented that the impeded proliferation induced by HNF1A-AS1 downregulation was partially overturned by introduction of HMGB1 (Fig. 6B, C). Furthermore, the flow cytometry assay proved that the enhanced apoptosis rate caused by downregulation of HNF1A-AS1 was, in part, recovered by overexpression of HMGB1 (Fig. 6D). Additionally, results of the transwell assay showed that repressed migration and invasion ability induced by reduced HNF1A-AS1 were regained by HMGB1 augmentation (Fig. 6E, F). Besides, the authors detected the level of HMGB1 in the cell supernatant by enzyme-linked immunosorbent assay. Results showed that levels of HMGB1 in the supernatant of U2OS and Saos-2 cells were elevated compared with hFOB1.19 cells (Supplementary Fig. S2A).
In addition, levels of HMGB1 in the supernatant of U2OS and Saos-2 cells were negatively regulated by miR-32-5p (Supplementary Fig. S2B, C). Moreover, HNF1A-AS1 knockdown downregulated HMGB1 levels in the supernatant of U2OS and Saos-2 cells, while this influence was overturned by miR-32-5p inhibition (Supplementary Fig. S2D, E). In addition, the reduction of levels of HMGB1 in the supernatant of U2OS and Saos-2 cells caused by HNF1A-AS1 silencing was recovered by HMGB1 overexpression (Supplementary Fig. S2D, E). Collectively, these results suggested that HNF1A-AS1 reduction regulated proliferation, apoptosis, migration, and invasion of OS cells through the miR-32-5p/HMGB1 axis.
Discussion
OS has a high lung metastasis and local recurrence rate, which threatens the health of children and adolescents. In recent years, a substantial amount of evidence exhibits that lncRNAs are emerging regulators of cell fate and gene expression and exert vital roles in tumorigenesis. Hence, the authors aimed to investigate the role of HNF1A-AS1 and its molecular mechanism in OS in this study.
According to previous literature, HNF1A-AS1 exerted a carcinogenic role in series of tumors. For instance, enhanced HNF1A-AS1 expedited metastasis of colon cancer by impeding the feedback loop of miR-34a-STRT1-p53. 9 One report revealed that elevation of HNF1A-AS1 accelerated the cell cycle, proliferation, migration, and invasion in nonsmall cell lung cancer. 27 In addition, Wang et al. reported that high HNF1A-AS1 expression boosted metastasis and growth of esophageal squamous cell carcinoma through binding to miR-214 to regulate SOX4 expression. 10
This study also proved a marked enhancement in HNF1A-AS1 in OS tissues and cells, and enhanced HNF1A-AS1 was associated with the clinical stage, lung metastases, and overall survival. Furthermore, silencing of HNF1A-AS1 facilitated cell apoptosis and hindered proliferation, migration, and invasion in OS cells. The previous study revealed that HNF1A-AS1 was augmented in OS tissues and cells, and reduction of HNF1A-AS1 repressed cell G1/S transition and proliferation, invasion, and migration. 28 Moreover, Zhao et al. also revealed that HNF1A-AS1 elevation promoted cell metastasis and proliferation in OS by activating the Wnt/β-catenin pathway. 29 This study was consistent with the above studies, indicating that HNF1A-AS1 exerted a carcinogenic role in tumors.
A great deal of evidence has identified that HNF1A-AS1 can expedite diverse cancers by interacting with miRNAs and repressing their expression. 30,31 In the present study, the authors utilized the starBase database prediction and confirmed by the luciferase reporter assay that miR-32-5p was a target for HNF1A-AS1. It was reported that miR-32-5p was an antitumor gene in cancer. Liu et al. showed that in cervical cancer tissues and cells, miR-32-5p was reduced and the enhancement of miR-32-5p repressed migration, clone formation, proliferation, and invasion of HeLa cells. 17 Another study revealed that in clear cell renal cell cancer, the elevation of miR-32-5p repressed metastasis of clear cell renal cell cancer cells by shifting the signaling pathway of miR-32-5p/TR4/HGF/Met. 32
In this study, miR-32-5p was decreased in OS tissues and cells. Moreover, miR-32-5p elevation induced cell apoptosis and impeded proliferation, migration, and invasion in OS cells. Besides, previous literature indicated that miR-32 was reduced in OS tissues and the augmentation of miR-32 restrained proliferation of OS cells by downregulating Sox9. 33 From all the above, the authors deduced that miR-32-5p was a suppressor gene in OS.
In general, miRNA exerts its effects in different cancers by targeting different genes. In the current study, the authors employed the starBase database and luciferase reporter assay and confirmed that HMGB1 was a target gene of miR-32-5p. Moreover, HMGB1 was elevated in OS tissues and cells, and HNF1A-AS could regulate HMGB1 expression by targeting miR-32-5p. In addition, HMGB1 overexpression weakened the effect of HNF1A-AS1 downregulation on cell apoptosis, proliferation, migration, and invasion in OS cells.
One report presented that HMGB1 was enhanced in OS cells, and HMGB1 inhibition repressed the progression of OS by downregulating the lncRNA MALAT1 gene. 34 Liu et al. indicated that augmentation of HMGB1 partly overturned the inhibitory effect of miR-505 upregulation on migration, proliferation, and proliferation. 35 In addition, HMGB1 could act as a damage-associated molecular pattern molecule and was regulated by the HNF1A-AS1/miR-32-5p/HMGB1 axis. Therefore, the authors concluded that knockdown of HNF1A-AS1 enhanced apoptosis and impeded proliferation, migration, and invasion of OS cells.
Conclusions
In conclusion, silencing of HNF1A-AS1 constrained proliferation, migration, and invasion and expedited apoptosis of OS cells by elevating HMGB1 expression by binding to miR-32-5p. This research showed that HNF1A-AS1 might be a potential biomarker and therapeutic target for the diagnosis and treatment of OS.
Footnotes
Authors' Contributions
P.L. and X.Z. were involved in conception and design; T.D. was involved in development of methodology; P.L. and T.D. were involved in acquisition, analysis, and interpretation of data; P.L. was involved in writing, review, and revision of the article; T.D. was involved in administrative, technical, and material support; and X.Z. was involved in study supervision. All coauthors reviewed and approved of the article before submission.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was approved by the Natural Science Foundation of Hubei Province, China (2017CFC808, 2018CFC859).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.