
Abstract
Background:
Circular RNAs (circRNAs) have been reported to be implicated in the pathogenesis of prostate cancer (PCa). Herein, the authors explore the role and molecular mechanism of circRNA SWI/SNF-related, matrix-associated, actin-dependent regulator of chromatin, subfamily a, member 5 (circSMARCA5) in PCa.
Materials and Methods:
The levels of circSMARCA5, SMARCA5, miR-432, and programmed cell death 10 (PDCD10) were determined by quantitative real-time polymerase chain reaction (qRT-PCR). The circular structure and stability of circSMARCA5 were validated by qRT-PCR using Oligo dT primer, transcriptional inhibitor actinomycin D, or RNase R treatment, respectively. Cell proliferation, migration, invasion, epithelial/mesenchymal transition (EMT), and glycolysis were detected by 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), transwell migration and invasion assays, Western blot assay, and Glucose or Lactate Detection Kit, respectively. The target relationship between miR-432 and circSMARCA5 or PDCD10 was validated by dual-luciferase reporter assay and RNA immunoprecipitation (RIP) assay. Western blot was performed to detect the protein expression of PDCD10 in PCa cells.
Results:
CircSMARCA5 was aberrantly upregulated, and was a circular and stable RNA in PCa cells. CircSMARCA5 accelerated the proliferation, metastasis, and glycolysis of PCa cells. MiR-432 was a direct target of circSMARCA5, and circSMARCA5 accelerated the development of PCa through miR-432 in PCa cells. PDCD10 was a direct target of miR-432, and PDCD10 addition reversed the inhibitory effects of miR-432 accumulation on the proliferation, metastasis, and glycolysis of PCa cells. CircSMARCA5 upregulated the expression of PDCD10 through sponging miR-432 in PCa cells.
Conclusion:
CircSMARCA5 deteriorated PCa through the miR-432/PDCD10 axis. CircSMARCA5/miR-432/PDCD10 axis might be an underlying therapeutic target for PCa treatment.
Introduction
Prostate cancer (PCa) is a commonly diagnosed cancer in males all over the world. 1 The conventional therapeutic strategies for PCa are chemotherapy, radiotherapy, and surgery. The pathogenesis of PCa is complex, and the genetic factor is identified as the leading factor. 2 Therefore, it is imperative to investigate the potential mechanism of PCa occurrence and development.
Circular RNAs (circRNAs) have been implicated in the progression of PCa. For instance, Feng et al. claimed that circ0005276 accelerated the growth and metastasis of PCa cells through FUS and XIAP. 3 Huang et al. demonstrated that circABCC4 promoted the development of PCa through miR-1182/FOXP4 axis. 4 Herein, the authors focused on the function of circRNA SWI/SNF-related, matrix-associated, actin-dependent regulator of chromatin, subfamily a, member 5 (circSMARCA5) in PCa. CircSMARCA5 is generated from the SMARCA5 gene, and has been reported to participate in the development of PCa and gastric cancer. 5,6 Kong et al. demonstrated that the abundance of circSMARCA5 was elevated in PCa, and circSMARCA5 played an oncogenic role in PCa. 6 However, the precise mechanism of circSMARCA5 in PCa remains largely unknown.
MicroRNAs (miRNAs) could post-transcriptionally regulate the expression of target genes through inhibiting translation or promoting the degradation of messenger RNAs (mRNAs). 7,8 Accruing articles have demonstrated that miRNAs are involved in cell growth, apoptosis, and metastasis. 9 –12 MiR-432 was downregulated in various cancers, such as cervical cancer, glioma, osteosarcoma, and PCa. 13 –16 Herein, the authors aimed to explore the role and the signal network behind miR-432 in PCa.
Programmed cell death 10 (PDCD10) encodes an intracellular apoptosis-related protein. 17 Accumulating articles have reported that PDCD10 was implicated in the pathogenesis of pancreatic adenocarcinomas and colorectal cancer. 18,19 Fu et al. claimed that miR-103 suppressed the proliferation and metastasis while promoted the apoptosis of PCa cells through negatively regulating PDCD10. 20 However, the expression pattern and function of PDCD10 in PCa are not fully addressed.
Herein, the authors investigated the role of circSMARCA5 in PCa cells, and they investigated the potential mechanism by which circSMARCA5 accelerated the progression of PCa.
Materials and Methods
Tissue specimens and cells
PCa tissues (n = 30) and matching nontumor tissues (n = 30) used to measure the abundance of circSMARCA5 were obtained from patients at the Fourth Hospital of Hebei Medical University. Human material was obtained with written informed consents from patients and permission of the Ethics Committee of the Fourth Hospital of Hebei Medical University.
Human PCa cell lines (DU145 and 22RV1) and normal human prostate epithelial cell line RWPE-1 were purchased from BeNa Culture Collection (Beijing, China). DU145, 22RV1, and RWPE-1 cells were cultivated in Roswell Park Memorial Institute-1640 (RPMI-1640) medium (Gibco, Carlsbad, CA) added with 10% fetal bovine serum (FBS; Gibco), 100 U/mL penicillin/100 μg/mL streptomycin solution in a humidified incubator at 37°C with 5% CO2.
Quantitative real-time polymerase chain reaction
RNA was extracted from PCa tissues and cells with TRIzol reagent (Invitrogen, Carlsbad, CA). DNase I and RNase R were used to remove DNA, ribosomal RNA, and linear RNA and enrich circRNA. Reverse transcription was conducted through a Reverse Transcription Kit (TaKaRa, Liaoning, China). Quantitative real-time polymerase chain reaction (qRT-PCR) was conducted using the SYBR Premix Ex Taq™ kit (TaKaRa). U6 small nuclear RNA or β-actin served as the control, and the enrichment of circSMARCA5, SMARCA5, miR-432, and PDCD10 was measured through 2−ΔΔCt method. 21 The primers used in this study were shown as follows: circSMARCA5 (Forward, 5′-CTCCAAGATGGGCGAAAG-3′; Reverse, 5′-TGTGTTGCTCCATGTCTAATCA-3′), SMARCA5 (Forward, 5′-CGGCGGGAGCAACAGCAGCA-3′; Reverse, 5′-GTTTGCATTTTTTCTTCATA-3′), miR-432 (Forward, 5′-AACGAGACGACGACAGACT-3′; Reverse, 5′-CTTGGAGTAGGTCATTGGGT-3′), PDCD10 (Forward, 5′-CGACTCACTATAGGGCCAAAAGAATTTACGCCTTAATGTG-3′; Reverse, 5′-GGTCGGTCCCATGGATCCCTTGCAGCATATCGCTTTCAAG-3′), U6 (Forward, 5′-GCTTCGGCAGCACATATACTAAAAT-3′; Reverse, 5′-CGCTTCACGAATTTGCGTGTCAT-3′), and β-actin (Forward, 5′-CCTCTCCCAAGTCCACACAG-3′; Reverse, 5′-GGGCACGAAGGCTCATCATT-3′).
Actinomycin D or RNase R treatment and nuclear/cytoplasmic fractionation
Actinomycin D or RNase R treatment was used to detect the stability of circSMARCA5 and SMARCA5 mRNA, and the nuclear/cytoplasmic fractionation was applied to ascertain the intracellular localization of circSMARCA5. Actinomycin D was mixed with total RNA at 0, 6, 12, 18, or 24 h, and the expression of circSMARCA5 and SMARCA5 mRNA was measured by qRT-PCR. Forty units of RNase R was added into 10 μg RNA for 15 min, and the abundance of circSMARCA5 and SMARCA5 mRNA was measured by qRT-PCR. Nuclear and cytoplasmic RNA fractionation was isolated with the PARIS™ Kit (Invitrogen).
Cell transfection
MiR-432 mimic (miR-432), miRNA negative control (miR-NC), miR-432 inhibitor (anti-miR-432) and its control (anti-miR-NC), small interfering RNA targeting circSMARCA5 (si-circSMARCA5), siRNA-NC (si-NC), small hairpin RNA targeting circSMARCA5 (sh-circSMARCA5), shRNA-NC (sh-NC), PDCD10 overexpression vector (pcDNA-PDCD10), and pcDNA empty vector (pcDNA) were acquired from GenePharma (Shanghai, China). Transfection was performed using Lipofectamine 3000 (Invitrogen).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
The 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was performed to detect the influence of circSMARCA5, miR-432, and PDCD10 on the proliferation of PCa cells. DU145 and 22RV1 cells were seeded into 96-well plates, and the viability of PCa cells was measured by MTT reagent (Invitrogen) following transfection for 0, 24, 48, and 72 h. Dimethyl sulfoxide (DMSO; Sigma, St. Louis, MO) was used to dissolve the formazan. The absorbance was measured at 490 nm to detect the proliferation of PCa cells.
Transwell assay
For migration assay, DU145 and 22RV1 cells suspended in 100 μL serum-free medium were transferred into the upper chambers. Five hundred microliters of medium added with 10% FBS was pipetted to the lower chambers. The cells on the top side were removed after incubation for 48 h, and the migrated cells were fixed and dyed using 4% paraformaldehyde and 0.5% Crystal Violet. The migrated PCa cells were counted using a microscope. The capacity of invasion was measured using the upper chambers precoated with BD Matrigel matrix (BD Biosciences, San Jose, CA).
Western blot assay
After relevant treatment, DU145 and 22RV1 cells were collected, and Radioimmunoprecipitation assay (RIPA) lysis buffer (Beyotime, Shanghai, China) was used to isolate the proteins. The proteins were quantified through the bicinchoninic acid (BCA) method. Then 20 mg proteins were run on 10% gel and transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA). After 1 h blocking in the 5% nonfat milk, the membranes were incubated with primary antibodies targeting E-cadherin (ab1416; Abcam, Cambridge, MA), N-cadherin (ab18203; Abcam), Vimentin (ab92547; Abcam), matrix metallopeptidase 9 (MMP9; ab219372; Abcam), lactate dehydrogenase A (LDHA; ab226016, Abcam), PDCD10 (ab110531; Abcam), and β-actin (ab8226; Abcam) at 4°C overnight. The PVDF membranes were then incubated with secondary antibody (ab205718, Abcam). The protein signal was visualized with the enhanced chemiluminescent (ECL) system (Beyotime).
Glucose consumption and lactate production assays
DU145 and 22RV1 cells were maintained in glucose-free RPMI-1640 medium for 16 h. Subsequently, the PCa cells were cultured in high-glucose RPMI-1640 medium for a further 24 h. The glucose level in the cells and the lactate level in the medium were measured by the Fluorescence-based Glucose Assay Kit (BioVision, Milpitas, CA) and Fluorescence-Based Lactate Assay Kit (BioVision).
Lactate dehydrogenase assay
After transfection, LDHA activity was assessed in DU145 and 22RV1 cells by the LDH Cytotoxicity Assay Detection Kit (Beyotime) according to the manufacturer's instructions.
Dual-luciferase reporter assay
The sequences of circSMARCA5 (mutant or wild-type) and the 3′ untranslated region (3′ UTR) of PDCD10 (mutant or wild-type) were cloned to the pmirGLO vector (Promega, Madison, WI). Luciferase activity was examined through dual-luciferase reporter assay system (Promega) in DU145 and 22RV1 cells cotransfected with miR-NC + circSMARCA5-WT, miR-NC + circSMARCA5-MUT, miR-432 + circSMARCA5-WT, or miR-432 + circSMARCA5-MUT. The validation of the combination between miR-432 and PDCD10 was performed following a similar approach.
RNA immunoprecipitation assay
DU145 and 22RV1 cells were harvested and lysed in RNA immunoprecipitation (RIP) buffer (Millipore), and the RIP assay was performed using the Magna RIP™ RNA-Binding Protein Immunoprecipitation Kit (Millipore, Billerica, MA), anti-Argonaute-2 (Anti-Ago2; Millipore), and anti-Immunoglobulin G (Anti-IgG; Millipore).
Animal treatment
Animal assay protocols were permitted by the Ethics Committee of the Fourth Hospital of Hebei Medical University. DU145 cells stably transfected with sh-circSMARCA5 or sh-NC were harvested and resuspended in phosphate-buffered saline (PBS) solution. Then 1 × 106 DU145 cells were subcutaneously injected into the BALB/c nude mice (Orient Bio, Inc., Seongnam, South Korea), and the tumor size was detected per week after injection for 7 d. The weight of tumors was recorded following injection for 35 d.
Statistical analysis
Data are presented as the mean ± standard deviation (SD) from at least three independent experiments. Comparisons were assessed by Student's t-test or one-way analysis of variance (ANOVA) followed by Tukey's test by GraphPad Prism 7 software. p value less than 0.05 was considered statistically significant.
Results
The abundance of circSMARCA5 is notably upregulated in PCa tissues and cells
CircSMARCA5 was generated from exon 15 and exon 16 of the SMARCA5 gene using circRNADb and CircInteractome databases (Fig. 1A). As exhibited in Figure 1B and C, the expression of circSMARCA5 was prominently elevated in PCa tissues and cells compared with that in corresponding normal tissues and human normal prostate epithelial cells, RWPE-1. CircSMARCA5 had a longer half-life compared with linear SMARCA5 mRNA in PCa cells treated with transcriptional inhibitor actinomycin D (Fig. 1D, E). Besides, circSMARCA5 was resistant to RNase R compared with linear SMARCA5 mRNA in the two PCa cells (Fig. 1F, G). These findings suggested that circSMARCA5 was more stable than linear SMARCA5 mRNA in PCa cells. qRT-PCR revealed that circSMARCA5 was mainly found in the cytoplasm of PCa cells (Fig. 1H, I). To test the circular structure of circSMARCA5, Random primer or Oligo dT primer was used to amplify circSMARCA5 or SMARCA5 mRNA in the two PCa cells. The level of circSMARCA5 was lower in DU145 and 22RV1 cells using Oligo dT primer (Fig. 1J, K). The expression of SMARCA5 mRNA remained unchanged in Random primer group and Oligo dT group, suggesting that circSMARCA5 do not have poly-A tail. Collectively, circSMARCA5 was a circular and stable RNA, and it was highly expressed in PCa tissues and cells.
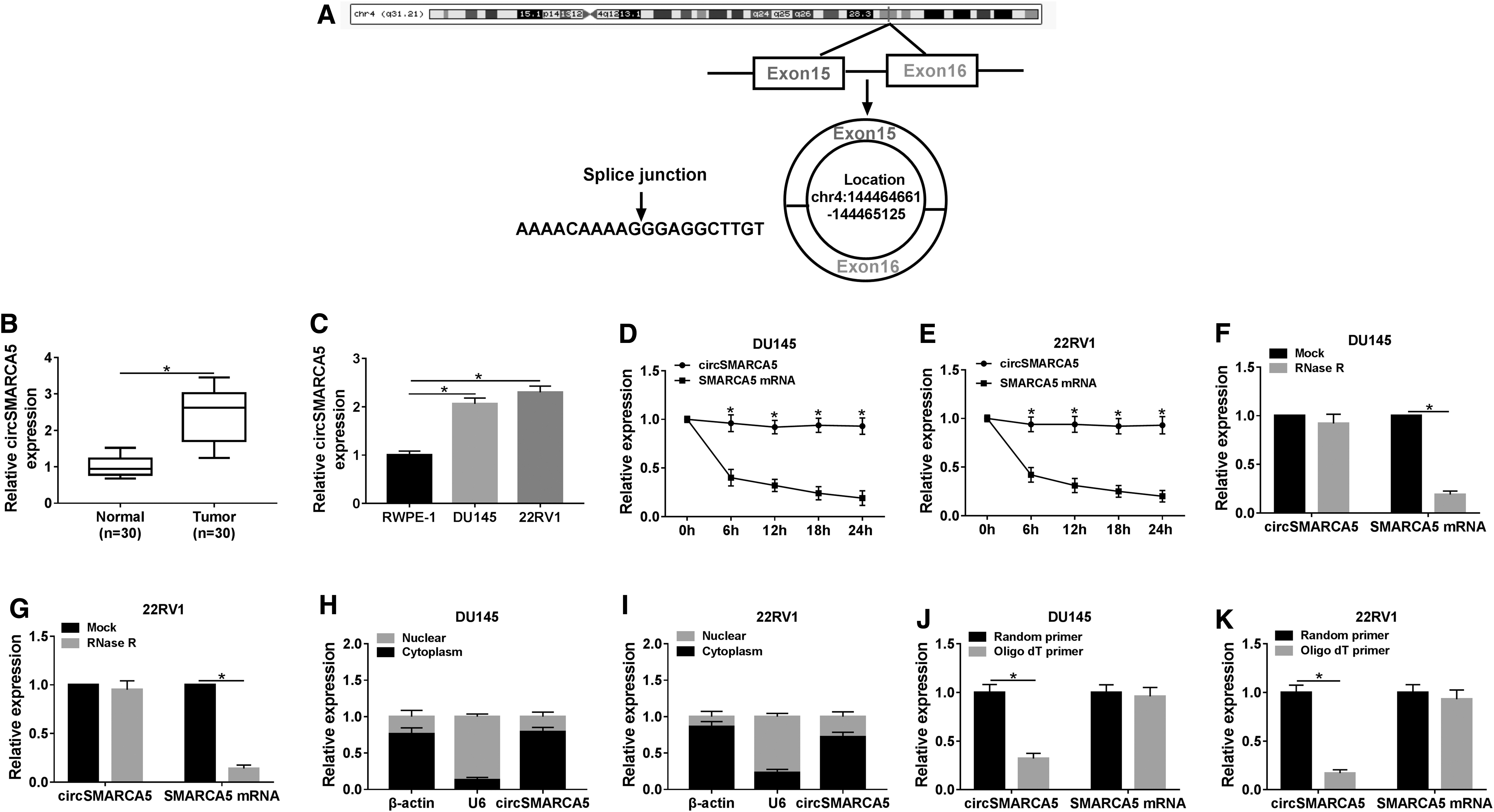
The abundance of circSMARCA5 is notably upregulated in PCa tissues and cells.
CircSMARCA5 depletion inhibits the proliferation, metastasis, and glycolysis of PCa cells
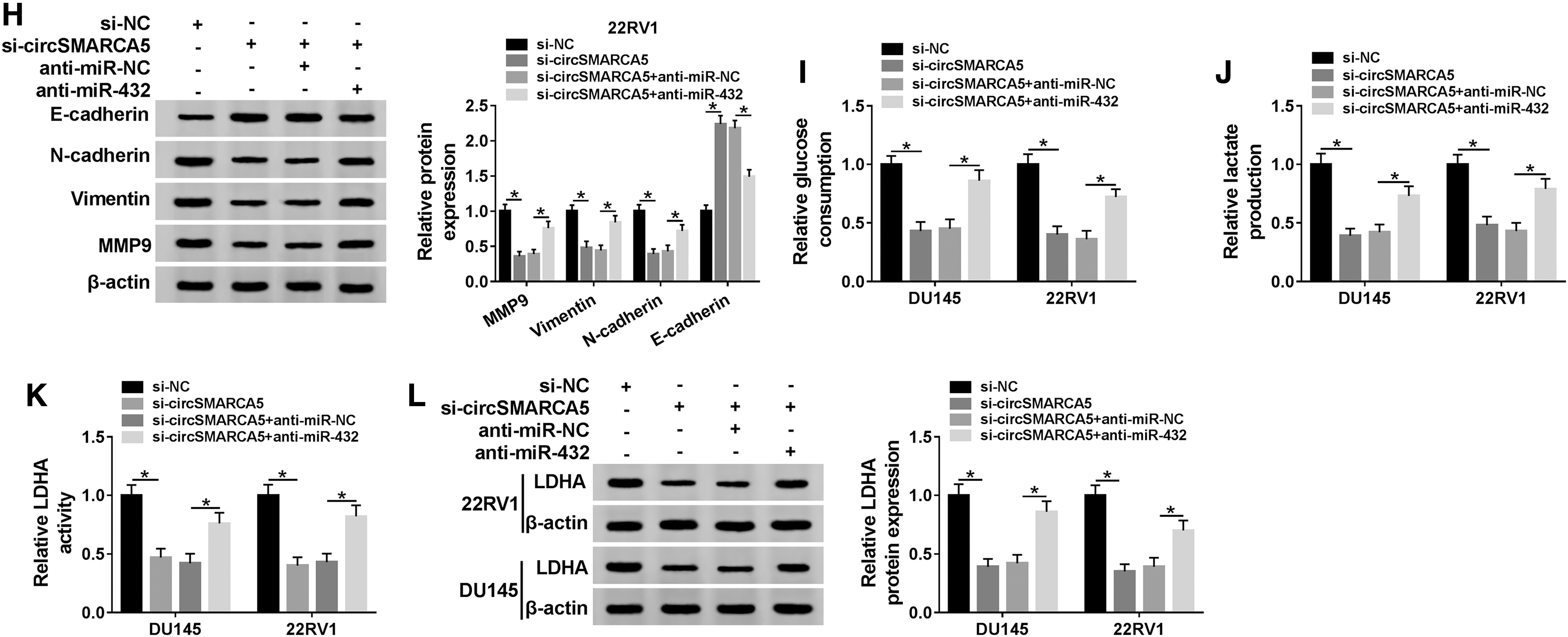
To clarify the role of circSMARCA5 in PCa, the authors conducted loss-of-function experiments. First, they evaluated the knockdown efficiency of si-circSMARCA5 in DU145 and 22RV1 cells. As shown in Figure 2A, the transfection of si-circSMARCA5 triggered a significant decrease in the expression of circSMARCA5 in PCa cells. Subsequently, they found that the depletion of circSMARCA5 restrained the proliferation of DU145 and 22RV1 cells (Fig. 2B, C). Transwell migration and invasion assays revealed that circSMARCA5 intervention inhibited the migration and invasion of PCa cells (Fig. 2D, E). Apart from this, they detected the epithelial/mesenchymal transition (EMT) markers (E-cadherin, N-cadherin, Vimentin, and MMP9) to assess the influence of circSMARCA5 depletion on the EMT of PCa cells. As mentioned in Figure 2F and G, circSMARCA5 depletion reduced the abundance of N-cadherin, Vimentin, and MMP9, whereas upregulated the level of E-cadherin, demonstrating that circSMARCA5 interference inhibited the EMT of PCa cells. The glucose consumption and lactate production were prominently decreased with the depletion of circSMARCA5 in PCa cells, suggesting that circSMARCA5 depletion restrained the glycolysis of PCa cells (Fig. 2H, I). LDHA is a crucial enzyme in the process of glycolysis, and they also assessed the impact of circSMARCA5 intervention on the catalytic activity and expression of LDHA in PCa cells. As exhibited in Figure 2J and K, the activity and level of LDHA were markedly reduced by the transfection of si-circSMARCA5 in PCa cells. Taken together, circSMARCA5 promoted the progression of PCa through facilitating the proliferation, metastasis, and glycolysis of PCa cells.

CircSMARCA5 depletion inhibits the proliferation, metastasis, and glycolysis of PCa cells.
MiR-432 is a direct target of circSMARCA5 in PCa cells
MiR-432 was predicted as a target of circSMARCA5 by CircInteractome and Starbase softwares (Fig. 3A). As mentioned in Supplementary Figure S1A, they selected five candidate targets that were predicted by Starbase software and were downregulated in PCa. 16,22 –25 Among these five miRNAs, miR-432 was the most significantly upregulated miRNA. Therefore, miR-432 was chosen for further studies. Luciferase activity remarkably declined in the miR-432 and circSMARCA5-WT cotransfected group in DU145, and in 22RV1 cells, suggesting that miR-432 was a direct target of circSMARCA5 in PCa cells (Fig. 3B, C). RIP assay also showed that circSMARCA5 was present in RNA-induced silencing complex, likely through the combination with miR-432 in PCa cells (Fig. 3D, E). Subsequently, they found that miR-432 was downregulated in PCa cells compared with that in human normal prostate epithelial cells, RWPE-1 (Fig. 3F). Meanwhile, miR-432 was negatively regulated by circSMARCA5 in DU145 and 22RV1 cells (Fig. 3G). Collectively, miR-432 was a target of circSMARCA5, and it was negatively regulated by circSMARCA5 in PCa cells.
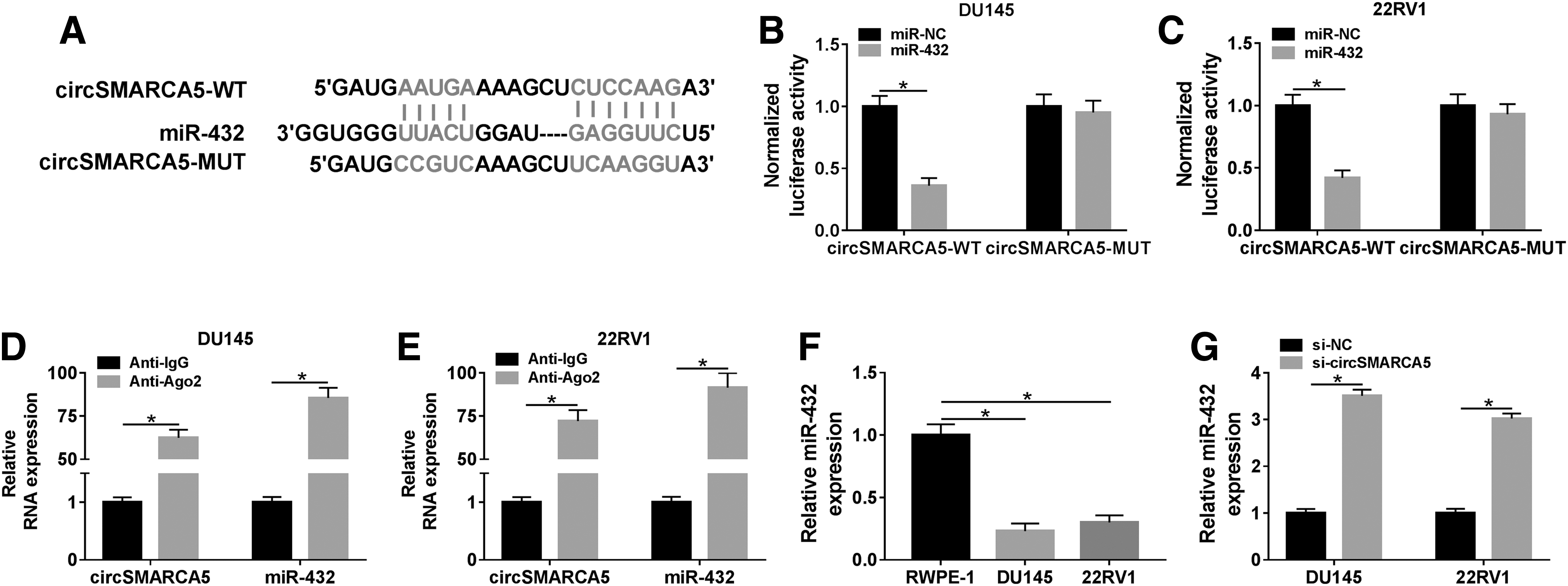
MiR-432 is a direct target of circSMARCA5 in PCa cells.
CircSMARCA5 promotes the proliferation, metastasis, and glycolysis of PCa cells through miR-432
To test whether miR-432 was involved in circSMARCA5-mediated development of PCa, they conducted the following experiments. The expression of miR-432 was notably reduced by the transfection of anti-miR-432 in PCa cells (Fig. 4A). DU145 and 22RV1 cells were transfected with si-NC, si-circSMARCA5, si-circSMARCA5 + anti-miR-NC, or si-circSMARCA5 + anti-miR-432. As indicated in Figure 4B, the transfection of anti-miR-432 decreased the abundance of miR-432, which was enhanced by the depletion of circSMARCA5 in PCa cells. The transfection of anti-miR-432 also alleviated the suppressive effects of circSMARCA5 intervention on the proliferation, migration, invasion, EMT, and glycolysis of PCa cells (Fig. 4C–L). Taken together, circSMARCA5 accelerated the progression of PCa through downregulating miR-432.
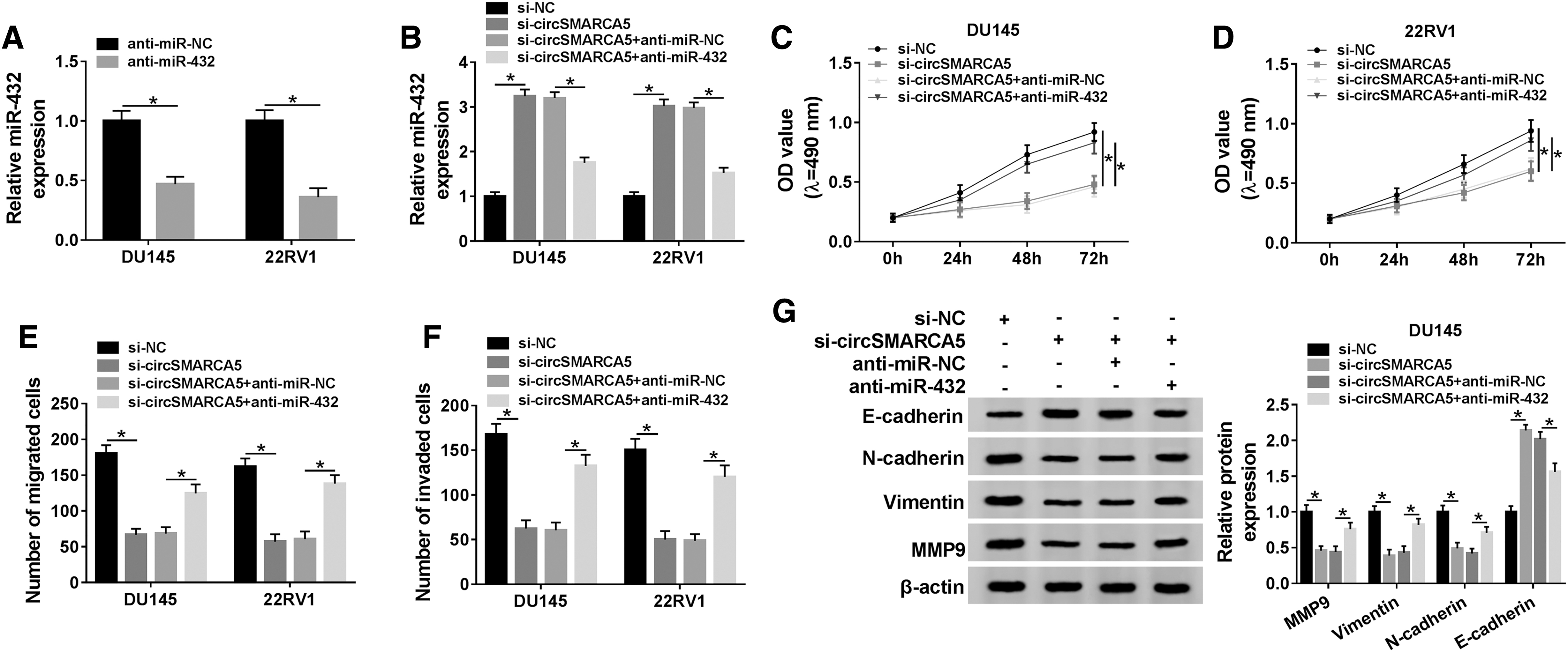
CircSMARCA5 promotes the proliferation, metastasis, and glycolysis of PCa cells through miR-432.
PDCD10 is a target of miR-432, and it is inversely regulated by miR-432 in PCa cells
To illustrate the potential mechanism by which circSMARCA5/miR-432 axis modulating the progression of PCa, they intended to investigate the downstream genes of miR-432 in PCa cells. There were five possible targets that were simultaneously predicted by Starbase software and reported to be upregulated in PCa. 20,26 –29 PDCD10 was selected for further experiments because it was the most significantly downregulated gene with the accumulation of miR-432 in DU145 cells (Supplementary Fig. 1B). As exhibited in Figure 5A, the complementary sequence between PDCD10 and miR-432 was predicted by Starbase bioinformatic software. MiR-432 accumulation markedly reduced the luciferase activity in PDCD10 3′ UTR-WT group, while the luciferase activity remained unchanged in miR-432 and PDCD10 3′ UTR-MUT cotransfected group in PCa cells (Fig. 5B, C), suggesting that PDCD10 was a direct target of miR-432 in PCa cells.

PDCD10 is a target of miR-432 in PCa cells, and it is inversely regulated by miR-432.
To clarify the role of PDCD10 in PCa, they first determined the abundance of PDCD10 mRNA and protein in PCa cells and human normal prostate epithelial cells, RWPE-1. As mentioned in Figure 5D and E, the mRNA and protein expression of PDCD10 was higher in PCa cells than that in human normal prostate epithelial cells, RWPE-1, implying the oncogenic role of PDCD10 in PCa. The enrichment of miR-432 was remarkably upregulated in DU145 and 22RV1 cells transfected with miR-432 mimic (Fig. 5F). MiR-432 accumulation notably decreased the mRNA and protein abundance of PDCD10 in DU145 and 22RV1 cells (Fig. 5G, H). Collectively, PDCD10 was a direct target of miR-432, and it was negatively modulated by miR-432 in PCa cells.
PDCD10 accumulation attenuates the inhibitory effects of miR-432 overexpression on the proliferation, metastasis, and glycolysis of PCa cells
The abundance of PDCD10 mRNA and protein was conspicuously elevated by the transfection of pcDNA-PDCD10 in DU145 and 22RV1 cells (Fig. 6A, B). To elucidate whether PDCD10 was involved in miR-432-mediated proliferation, metastasis, and glycolysis of PCa cells, DU145 and 22RV1 cells were transfected with miR-NC, miR-432, miR-432 + pcDNA, or miR-432 + pcDNA-PDCD10. As mentioned in Figure 6C–H, PDCD10 overexpression reversed the inhibitory effects of miR-432 addition on the proliferation and metastasis of PCa cells. Besides, the consumption of glucose, the production of lactate and the activity and the protein expression of LDHA were inhibited by the transfection of miR-432 mimic, and the cotransfection of miR-432 and pcDNA-PDCD10 counteracted these inhibitory effects caused by miR-432 overexpression (Fig. 6I–N). Taken together, miR-432 suppressed the proliferation, metastasis, and glycolysis of PCa cells through targeting PDCD10.
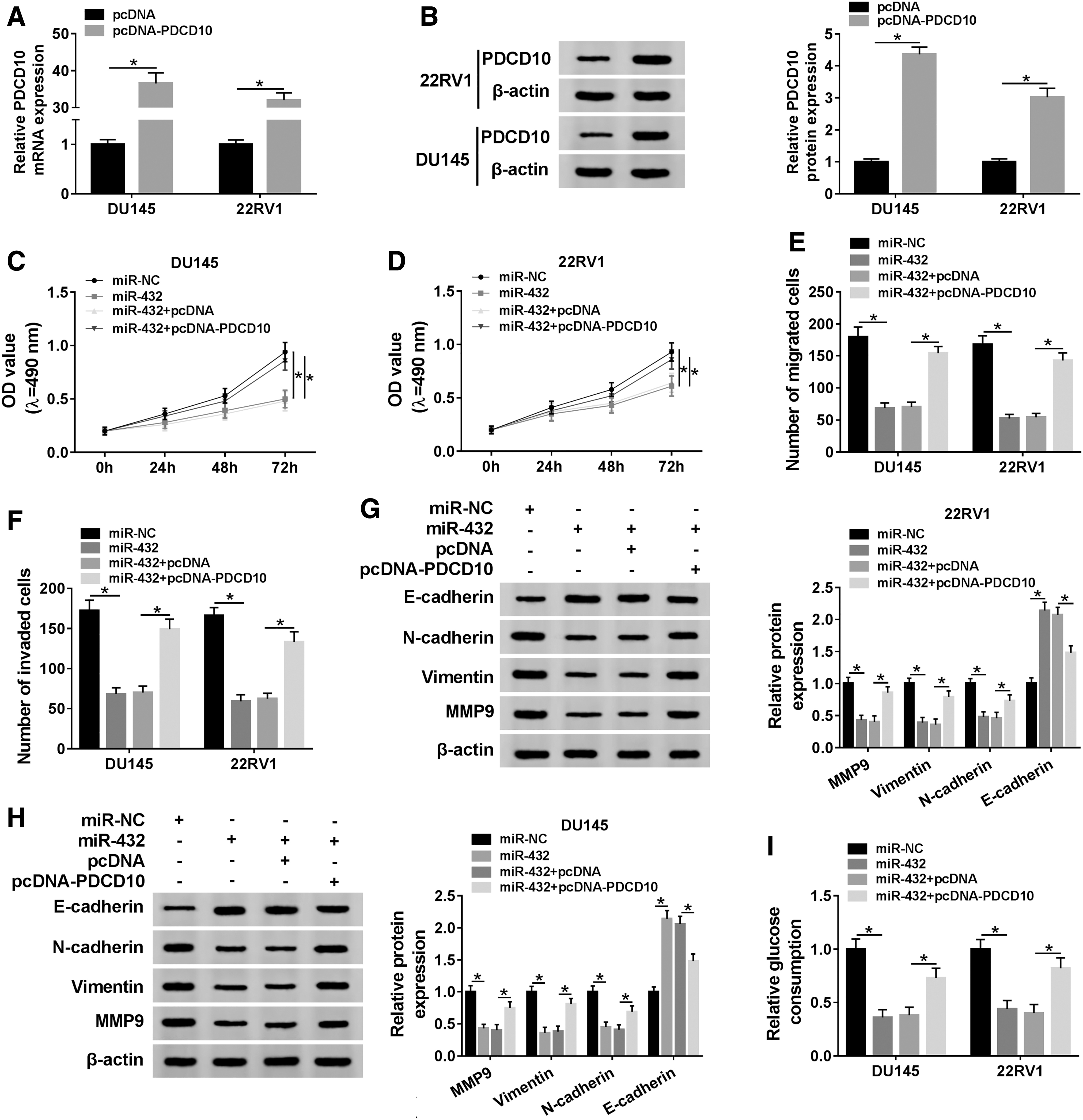
PDCD10 accumulation attenuates the inhibitory effects of miR-432 overexpression on the proliferation, metastasis, and glycolysis of PCa cells.
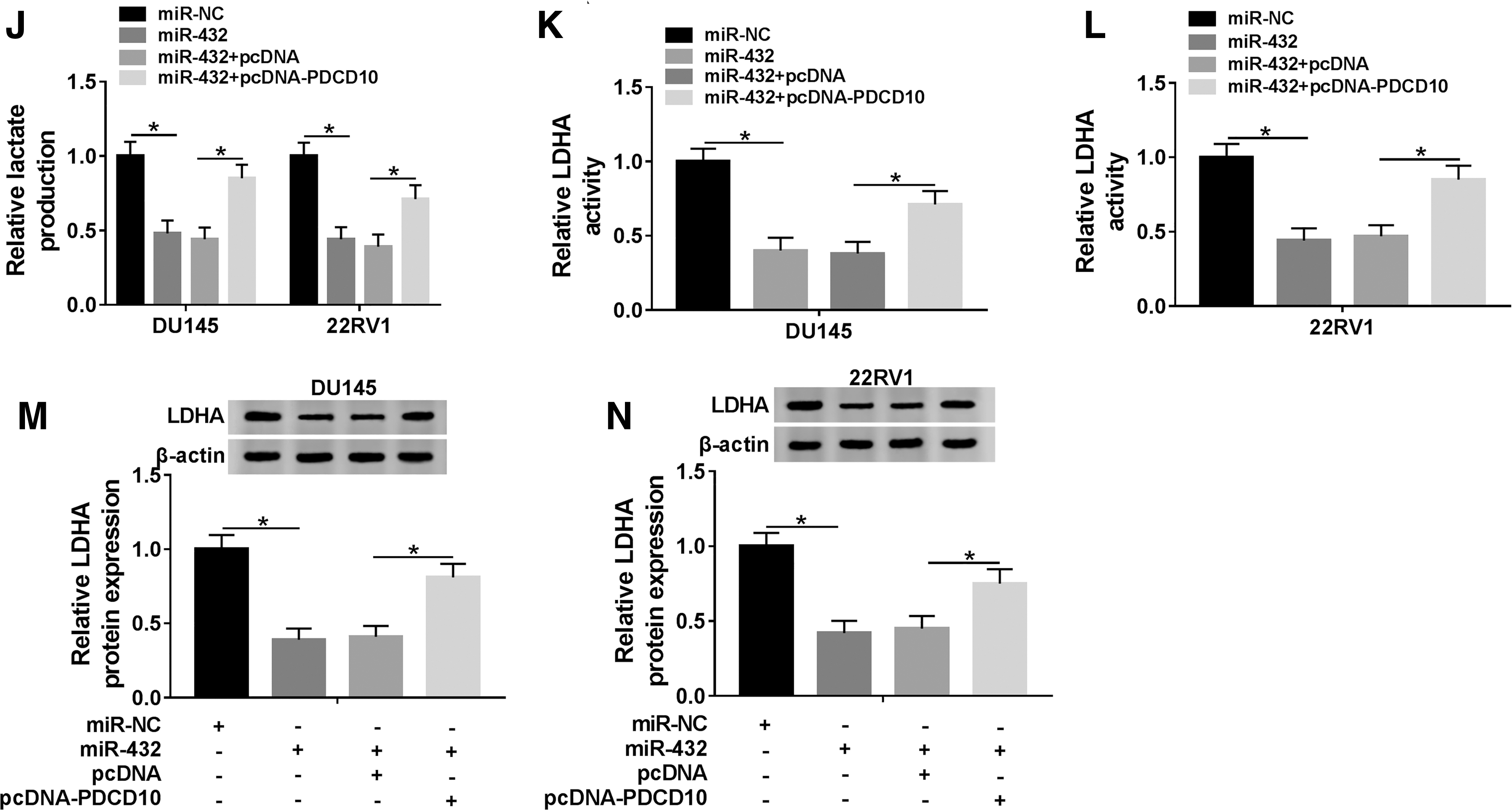
CircSMARCA5 upregulates the level of PDCD10 through sponging miR-432 in PCa cells
We transfected si-NC, si-circSMARCA5, si-circSMARCA5 + anti-miR-NC, or si-circSMARCA5 + anti-miR-432 into PCa cells to further clarify the relationship among circSMARCA5, miR-432, and PDCD10. As indicated in Figure 7A and B, the depletion of circSMARCA5 reduced the abundance of PDCD10 mRNA and protein, and the addition of anti-miR-432 recovered the mRNA and protein expression of PDCD10 in PCa cells. These findings suggested that PDCD4 was regulated by circSMARCA5/miR-432 axis in PCa cells.

CircSMARCA5 upregulates the level of PDCD10 through sponging miR-432 in PCa cells.
CircSMARCA5 promotes the progression of PCa through upregulating PDCD10 through sponging miR-432 in vivo
We built murine xenograft model using DU145 cells stably transfected with sh-NC or sh-circSMARCA5 to verify the function of circSMARCA5 in vivo. As exhibited in Figure 8A and B, the depletion of circSMARCA5 inhibited the growth of PCa tumors. Additionally, the expression of circSMARCA5 and PDCD10 mRNA and protein was decreased in sh-circSMARCA5 group compared with that in sh-NC group, while the abundance of miR-432 exhibited a reverse phenomenon (Fig. 8C–F). These findings revealed that circSMARCA5 facilitated the development of PCa through circSMARCA5/miR-432/PDCD10 in vivo.
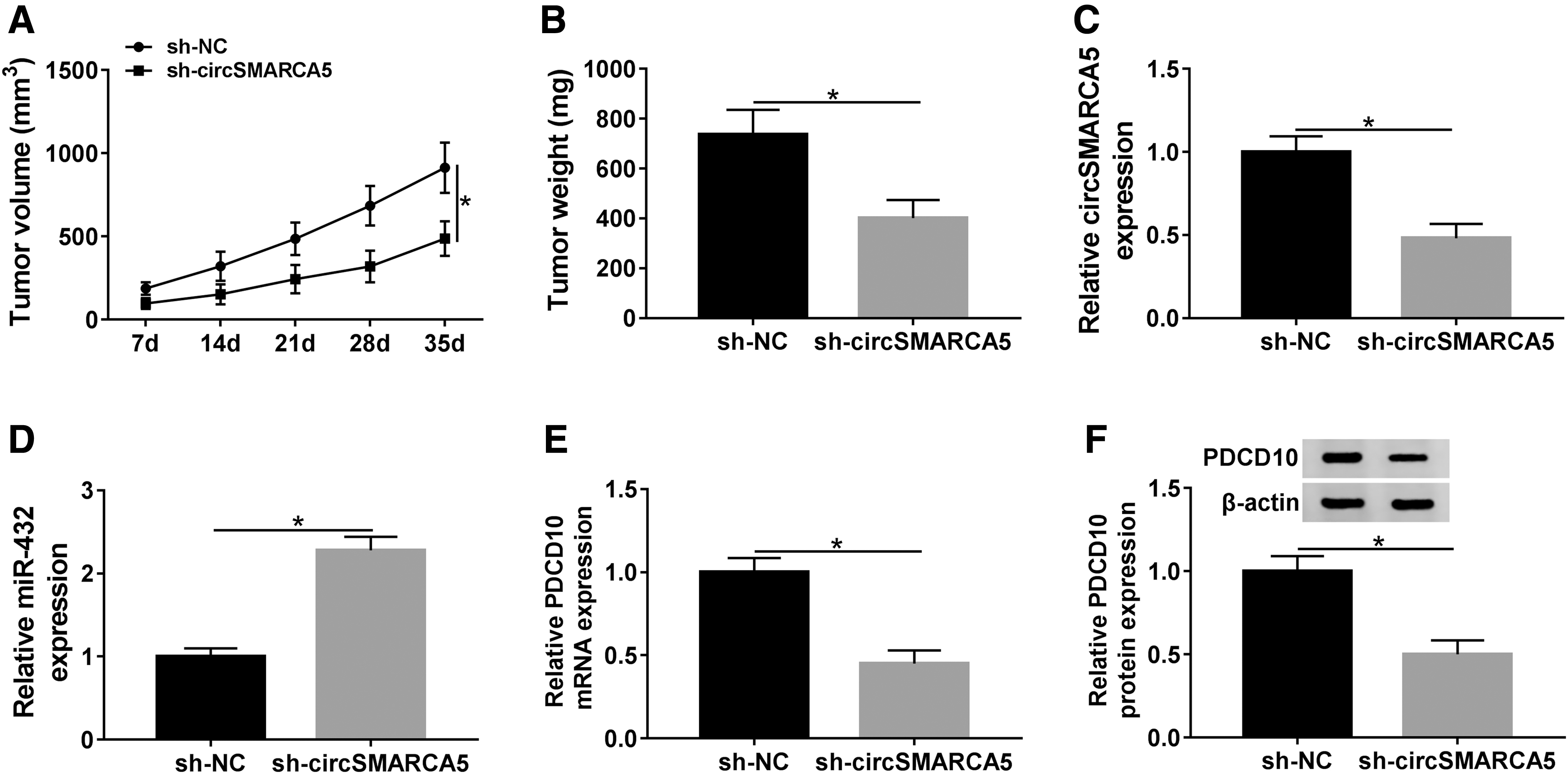
CircSMARCA5 promotes the progression of PCa through upregulating PDCD10 through sponging miR-432 in vivo.
Discussion
CircRNAs are a type of noncoding RNAs, which are characterized by the covalently closed loop structure. CircSMARCA5 derived from the SMARCA5 gene. SMARCA5 is a member of SWI/SNF family, and it has been reported to post-transcriptionally modulate the abundance of PSA in PCa cells. 30 CircSMARCA5 was upregulated in PCa tissues, and it facilitated the growth of PCa cells. 6 Herein, they found that the level of circSMARCA5 was prominently elevated in PCa tissues and cells, which was consistent with the above findings. Subsequently, they confirmed that circSMARCA5 was a circular and stable RNA. Loss-of-function experiments showed that circSMARCA5 promoted the proliferation, metastasis, and glycolysis of PCa cells.
MiR-432 was predicted as a target of circSMARCA5 by CircInteractome software, and dual-luciferase reporter assay and RIP assay confirmed the direct combination between miR-432 and circSMARCA5 in PCa cells. MiR-432 functioned as a tumor suppressor in multiple cancers. Jiang et al. demonstrated that the abundance of miR-432 was lower in hepatocellular carcinoma tissues and cells, and miR-432 inhibited the proliferation and tumorigenicity of hepatocellular carcinoma cells through Wnt/β-catenin signaling. 31 Chen et al. reported that miR-432 played a tumor suppressor role in lung adenocarcinoma through E2F3 and AXL. 32 Yang et al. demonstrated that lncRNA ZFAS1 facilitated the progression of glioma through downregulating miR-432-5p. 14 To test whether miR-432 was implicated in circSMARCA5-mediated progression of PCa, they conducted rescue experiments. The depletion of miR-432 ameliorated the inhibitory effects of circSMARCA5 intervention on the proliferation, metastasis, and glycolysis of PCa cells, suggesting that circSMARCA5 promoted the progression of PCa through downregulating miR-432. The tumor suppressor role of miR-432 was consistent with the above findings. 14,31,32
As mentioned above, miRNAs modulate the cellular biological processes through suppressing the translation or declining the enrichment of target mRNAs through the binding between miRNAs and the 3′ UTR of target mRNA. 7,8 To explore the precise mechanism of circSMARCA5/miR-432 in the progression of PCa, they attempted to search the targets of miR-432 in PCa cells. PDCD10 was predicted and validated as a direct target of miR-432 through Starbase software and dual-luciferase reporter assay. PDCD10 was an oncogene in PCa and bladder cancer. 20,33 Also, the authors performed rescue experiments to test whether miR-432 exerted its function through PDCD10 in PCa. PDCD10 addition counteracted the suppressive effects of miR-432 overexpression on the proliferation, metastasis, and glycolysis of PCa cells, demonstrating that PDCD10 served as a downstream gene of miR-432 to promote the development of PCa. Besides, they found that PDCD10 was modulated by circSMARCA5/miR-432 axis in PCa.
The murine xenograft model was used to investigate the effect of circSMARCA5 in vivo. The results revealed that the intervention of circSMARCA5 inhibited the growth of PCa tumors through circSMARCA5/miR-432/PDCD10 axis.
Conclusion
In conclusion, circSMARCA5 accelerated the progression of PCa through promoting the proliferation, metastasis, and glycolysis of PCa cells through circSMARCA5/miR-432/PDCD10 axis. Restoration of miR-432 expression might be an underlying therapeutic strategy for PCa.
Footnotes
Disclosure Statement
The authors declare that they have no financial conflicts of interest.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.