
Abstract
Background:
The resistance of glioma patients to temozolomide (TMZ) treatment is a limiting factor in clinical treatment. Circular RNA HIPK3 (circ-HIPK3) was found to be highly expressed in glioma, however, the role and potential mechanism of exosomal circ-HIPK3 from TMZ-resistant cells remain poorly unclear.
Methods:
Exosomes were characterized by transmission electron microscopy. The levels of all protein were detected by Western blot. Expression levels of circ-HIPK3, microRNA-421 (miR-421), and zinc finger protein of the cerebellum 5 (ZIC5) were measured by quantitative real-time polymerase chain reaction. The cell's 50% inhibitory concentration (IC50) of TMZ, apoptosis, and invasion were determined by methyl thiazolyl tetrazolium (MTT), flow cytometry, and Transwell assays, respectively. The correlation between miR-421 and circ-HIPK3 or ZIC5 was identified by dual-reporter luciferase and RNA immunoprecipitation (RIP) assays. The xenograft model was established to explore the effect of circ-HIPK3 in vivo.
Results:
circ-HIPK3 was obviously increased in TMZ-resistant glioma cells and their exosomes, miR-421, was downregulated in TMZ-resistant glioma. circ-HIPK3 directly targeted miR-421 and their expressions were negatively correlated in glioma tissues. Besides, circ-HIPK3 knockdown hampered the IC50 of TMZ, cell invasion, TMZ resistance, and triggered cell apoptosis, whereas these effects were reversed by transfection of anti-miR-421. ZIC5 was the target of miR-421 and ZIC5 overexpression weakened the inhibition effects of miR-421 on cell progression and TMZ resistance. More importantly, circ-HIPK3 depletion inhibited tumor growth by decreasing ZIC5 through sponging miR-421 in vivo.
Conclusion:
Exosomal circ-HIPK3 could promote cell progression and TMZ resistance by regulating miR-421/ZIC5 axis in TMZ-resistant glioma.
Introduction
Glioma, a tumor of the neuroepithelium, is not only aggressive but also the most lethal tumor in adults. 1,2 Despite the increasing effectiveness of surgical resection techniques and drug chemotherapy, the survival of glioma patients is still unsatisfactory. 3 Temozolomide (TMZ), an alkylating agent, can be converted into MTIC, which induces cell death through the mismatching repair of methylated admixture and thus produces antitumor effects. 4 Currently, TMZ is widely used as a first-line chemotherapy for patients with glioma. 5 However, it has been found that most patients gradually develop resistance to TMZ treatment leading to tumor recurrence. Therefore, understanding the underlying mechanism of patients' resistance to TMZ is of great significance for the treatment of glioma.
Exosomes, discoid vesicles with diameter ranging from 40 to 100 nm, contain complex and rich RNAs and proteins. 6 As important mediators of intercellular communication, these vesicles play an active role in tumor development and TMZ resistance, including glioma. 7 For instance, miR-1587 in exosomes released by mesenchymal stem cells could promote tumor tumorigenesis in glioblastoma. 8 Glioma cells could release exosomes containing lnc-CCAT2, thus driving angiogenesis and repressing apoptosis of endothelial cells. 9 However, the components of exosomes secreted by different cells are complex and different, there are still more RNA molecules from exosome to be discovered and explored.
Circular RNAs (circRNAs), a class of noncoding RNAs (ncRNAs), have strong stability due to their covalently closed continuous loop structures. 10 They have been found to be involved in regulating various physiological and pathological processes in human cancers. 11 In addition, circRNAs could act as a competitive endogenous RNAs (ceRNAs) of microRNAs (miRNAs) to modulate the biological activity of miRNA and the functional role of miRNA on target genes. 12 For example, circRNA NT5E could potentiate tumorigenesis through serving as miR-422a sponge in glioblastoma. 13 circ-HIPK3 is derived from Exon2 of the HIPK3 gene. Zheng et al. have previously demonstrated its circular structure by RNase R treatment. 14 circ-HIPK3 was considered to be a promoting cancer factor in gallbladder cancer 15 and hepatocellular carcinoma 16 by sponging miR-124. Interestingly, circ-HIPK3 was decreased and inhibited cell proliferation and metastasis in osteosarcoma. 17 So, the authors wanted to survey the function and potential mechanism of circ-HIPK3 in glioma.
MiR-421 has been shown to trigger cell proliferation, metastasis, tumorigenesis, and paclitaxel resistance in cancers. 18,19 Recently, miR-421 was found to reduce and hamper tumor development in glioma, 20 while little is known about the role of miR-421 in glioma and whether it can regulate TMZ resistance. Zinc finger protein of the cerebellum 5 (ZIC5), a member of the five-member ZIC family, has been widely reported to play a biological role as an oncogene in multiple tumors, 21,22 but, there are few reports about its functions in glioma. In this study, they aimed to explore the effects of exosome-transmitted circ-HIPK3 on glioma progression and TMZ resistance, and search for the molecular mechanism mediated by circ-HIPK3, hoping to find new targets for the treatment of TMZ-resistant glioma patients.
Materials and Methods
Patient samples
Tumor tissue and blood samples from 56 patients with glioma were collected at The Sixth Medical Center of PLA General Hospital. These glioma patients were divided into two groups based on their sensitivity to TMZ: sensitive (n = 25) and resistant (n = 31). Each patient signed a written informed consent. Serums were obtained by centrifuging these blood samples at 3000 g for 15 min and stored at −80°C. Our experiments were approved by the Ethics Committee of The Sixth Medical Center of PLA General Hospital.
Cell culture
Human astrocytes NHA and two glioma cell lines A172 and U251 were purchased from Shanghai Institutes for Biological Sciences (Shanghai, China). TMZ-resistant glioma cells A172/TR and U251/TR were established according to the study of Han et al. 23 A172 or U251 cells were exposed to TMZ (Sigma-Aldrich, St. Louis, MO), whose concentrations gradually increased from 5 μM to 100 μM for 6 months. The drug-resistant cell lines were named A172/TR and U251/TR, respectively. These cells were maintained in Roswell Park Memorial Institute 1640 (RPMI-1640; Hyclone, Shanghai, China) medium with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham, MA) and 1% penicillin/streptomycin at 37°C under 5% CO2.
Exosome isolation and identification
Exosomes were extracted from cells (NHA, A172, U251, A172/TR, and U251/TR) culture supernatants or the serum of glioma patients by ultra-high speed centrifugation method. Briefly, cell culture supernatants or the serum were differentially centrifuged (300 g for 10 min, 1000 g for 20 min, or 10,000 g for 30 min). After filtration, the supernatant ultracentrifuged at 100,000 g for 3 h. Then, the precipitations were washed twice with PBS and resuspended in PBS and stored at −80°C. Exosomes were visualized by transmission electron microscopy and affirmed by measuring the expressions of CD63 and CD9 using Western blot.
Western blot assay
The proteins of TMZ-resistant glioma tissues and cells were extracted by RIPA (Thermo Fisher Scientific). Then proteins were separated and transferred to polyvinylidene difluoride (PVDF; Beyotime, Shanghai, China) membranes. The membranes were blotted with primary antibodies against CD63 (1:1000; Abcam, Cambridge, MA), CD9 (1:2000; Abcam), P-glycoprotein (P-gp, 1:1000; Abcam), multidrug resistance associated protein 1 (MRP1, 1:2000; Abcam), Multidrug resistance 1 (MDR1, 1:1000; Abcam), ZIC5 (1:1000; Abcam), or Glyceraldehyde-3-phosphate dehydrogenase (GAPDH, 1:2000; Abcam) overnight at 4°C after sealing with 5% nonfat milk. Subsequently, the membranes were probed with horseradish peroxidase-conjugated goat anti-rabbit secondary antibody (1:3000, Abcam) for 1 h. The proteins were visualized by the BeyoECL Plus Kit (Beyotime).
Quantitative real-time polymerase chain reaction
TRIzol was applied to extract RNA from serum, glioma, and cultured cells (Thermo Fisher Scientific). For exosomal RNA extraction, TRIzol-LS (Thermo Fisher Scientific) was employed. The ABI® Reverse Transcription Kit (Applied Biosystems, Foster City, CA) was applied for reverse transcription. Then, quantitative real-time polymerase chain reaction (qRT-PCR) was performed on 7500 Real-Time PCR System (Applied Biosystems) using SYBR Select Master Mix (Applied Biosystems). GAPDH and U6 were served as internal references for circ-HIPK3 or ZIC5 and miR-421, respectively. Primer sequences: circ-HIPK3, 5′-TATGTTGGTGGATCCTGTTCGGCA-3′ (F), 5′-TGGTGGGTAGACCAAGACTTGTGA-3′ (R). GAPDH, 5′-GGGGCTCTCCAGAACATCATCC-3′ (F), 5′-ACGCCTGCTTCACCACCTCTT-3′ (R). miR-421, 5′-GATTGCACGTTTTCAGGTG-3′ (F), 5′-GCAGCACATCATTTATTACA-3′ (R). U6, 5′-CTCGCTTCGGCAGCACA-3′ (F), 5′-AACGCTTCACGAATTTGCGT-3′ (R). ZIC5, 5′-CGTTCCCGGCACACCC-3′ (F), 5′-GCCGTGGCGGAAAGGTC-3′ (R).
Transfection
Small interference RNA against circ-HIPK3 (si-circ-HIPK3: 5′-ACTACAGGTATGGCCTCACAA-3′) and the control si-NC (5′-TTCTCCGAACGTGTCACGTT-3′), miR-421 mimics (miR-421), miR-421 inhibitor (anti-miR-421), and their matched negative controls (miR-NC and anti-miR-NC) were synthesized by GenePharma (Shanghai, China). To construct circ-HIPK3 or ZIC5 overexpression plasmid, full length of circ-HIPK3 sequence or ZIC5 coding sequence (CDS) was cloned into pcDNA vector (Invitrogen, Carlsbad, CA), named as pcDNA-circ-HIPK3 (circ-HIPK3) or pcDNA-ZIC5 (ZIC5). Lentivirus packaged plasmid sh-circ-HIPK3 and its negative control sh-NC were synthesized by Sangon Biotech (Shanghai, China). Cells were transfected by using Lipofectamine 3000 (Invitrogen).
Fifty percent inhibitory concentration detection
A172/TR and U251/TR cells were transfected and incubated for 48 h, cells were treated with different concentrations of TMZ for 48 h. Then, 10 μL of methyl thiazolyl tetrazolium (MTT; Promega Corporation, Madison, WI) solution was added to the cells and incubation for another 2 h at 37°C. The absorbance at 490 nm was examined by a microplate reader and the 50% inhibitory concentration (IC50) of TMZ was calculated by SPASS 17.0.
Cell apoptosis assay
The six-well plates were applied for inoculating A172/TR and U251/TR cells. Transfection was performed for 48 h, then the harvested cells were stained with Annexin V Fluorescein Isothiocyanate and Propidium Iodide (BD Biosciences, Franklin Lake, NJ) for 15 min in the absence of light. Finally, cell apoptosis was analyzed by Flow cytometry using FlowJo 7.6 software.
Cell invasion assay
Transwell assay was employed to check cell invasion using the chambers coated with Matrigel (Becton Dickinson, Franklin Lakes, NJ). After 24 h of starvation, the transfected cells suspended with serum-free medium were inoculated into the upper chamber, and 600 μL of 10% FBS medium was added to the bottom chamber. After incubation for 48 h, the cells on the underside of the chamber were fixed and stained with Crystal Violet. The stained cells were observed with an inverted microscope and photographed.
Dual-luciferase reporter assay
To confirm the relationship between miR-421 and circ-HIPK3 or ZIC5, wide type (WT) sequence of circ-HIPK3 or ZIC5 3′UTR with miR-421-binding sites was cloned into the pmirGLO vector (YouBia, Changsha, China) to generate WT-circ-HIPK3 or ZIC5 3′UTR-WT. Their mutant reporter plasmids (MUT-circ-HIPK3 and ZIC5 3′UTR-MUT) without miR-421-binding sites were produced in the same way. These reporter plasmids were cotransfected with miR-421 or miR-NC in A172/TR and U251/TR cells. Forty-eight hours later, the luciferase activity was assessed by the Dual-Luciferase Reporter Kit (Promega Corporation).
RNA immunoprecipitation assay
RNA immunoprecipitation (RIP) assay was carried out to evaluate the correlation between miR-421 and circ-HIPK3 or ZIC5 by the EZ-Magna RIP Kit (Millipore, Billerica, MA). Briefly, A172/TR and U251/TR cells were harvested and lysed in RIP lysis buffer, following incubation with magnetic beads conjugated to human Ago2 antibody or IgG antibody. After being digested with proteinase K, the RNAs were extracted from the beads. Finally, qRT-PCR assay was performed to check the levels of circ-HIPK3, miR-421, or ZIC5.
Xenograft experiments
U251/TR cell-transfected sh-circ-HIPK3 or sh-NC was inoculated into the 4-week-old nude mice (n = 5 per group), and these mice were treated with TMZ (20 μg/mL) until the mice were euthanized. The tumor volume was measured 1 week after inoculation and then every 3 d for a total of seven times. Then, the mice were euthanized and the tumors were removed, and the weight was taken. Moreover, the RNAs and proteins from mice tumor tissues were extracted for qRT-PCR and Western blot analysis. The mouse experiments were permitted by the Animal Care and Use Committee of The Sixth Medical Center of PLA General Hospital.
Statistical analysis
Data were managed as the mean ± standard deviation (SD) from three independent experiments. Significance of differences between two groups or among multiple groups was assessed by Student's t-test or one-way analysis of variance (ANOVA). Pearson correlation coefficient was used to estimate the correlation between the two groups. It was deemed significant when the value of p was less than 0.05.
Results
circ-HIPK3 was elevated and miR-421 was decreased in TMZ-resistant glioma cells
Exosomes were first extracted from cell culture supernatant of TMZ-resistant glioma cell lines A172 and U251 (A172/TR and U251/TR). As shown in Figure 1A, the shape and size of the isolated exosomes were homogeneous and the vesicles showed a typical cup-shaped morphology. CD63 and CD9 have been shown to be marker proteins of exosomes, 24 and abundant expressions of CD63 and CD9 were detected by Western blot in cell exosomes, but not in supernatant of cell culture medium (Fig. 1B), which further confirmed our successful extraction of exosomes. Then, circ-HIPK3 expression in serum exosomes of glioma patients was detected by qRT-PCR. As displayed in Figure 1C, circ-HIPK3 was upregulated in TMZ-resistant serum exosomes compared with that in TMZ-sensitive serum exosomes. After A172 and U251 cells were treated with exosomes, circ-HIPK3 expression was augmented (Fig. 1D). Meanwhile, the expression of exosome circ-HIPK3 in A172/TR and U251/TR cells was higher than their parental cells (A172 and U251), and its expression in human astrocytes, NHA, was drastically declined compared with glioma cells A172 and U251 (Fig. 1E). The authors also checked circ-HIPK3 expression in different cell lines. As shown in Figure 1F, circ-HIPK3 was upregulated in glioma cell lines A172 and U251 compared with NHA cells, and its expression was highest in TMZ-resistant cell lines A172/TR and U251/TR. In addition, miR-421 expression was significantly decreased in TMZ-resistant glioma tissues (Fig. 1G). Compared with NHA, miR-421 was notably reduced in glioma cell lines A172 and U251, while its expression was lower in A172/TR and U251/TR cell lines (Fig. 1H). A significant negative correlation between circ-HIPK3 expression and miR-421 expression in glioma tissues was analyzed by Pearson correlation coefficient (Fig. 1I). These results indicated that circ-HIPK3 and miR-421 might be important regulators of glioma resistance to TMZ.
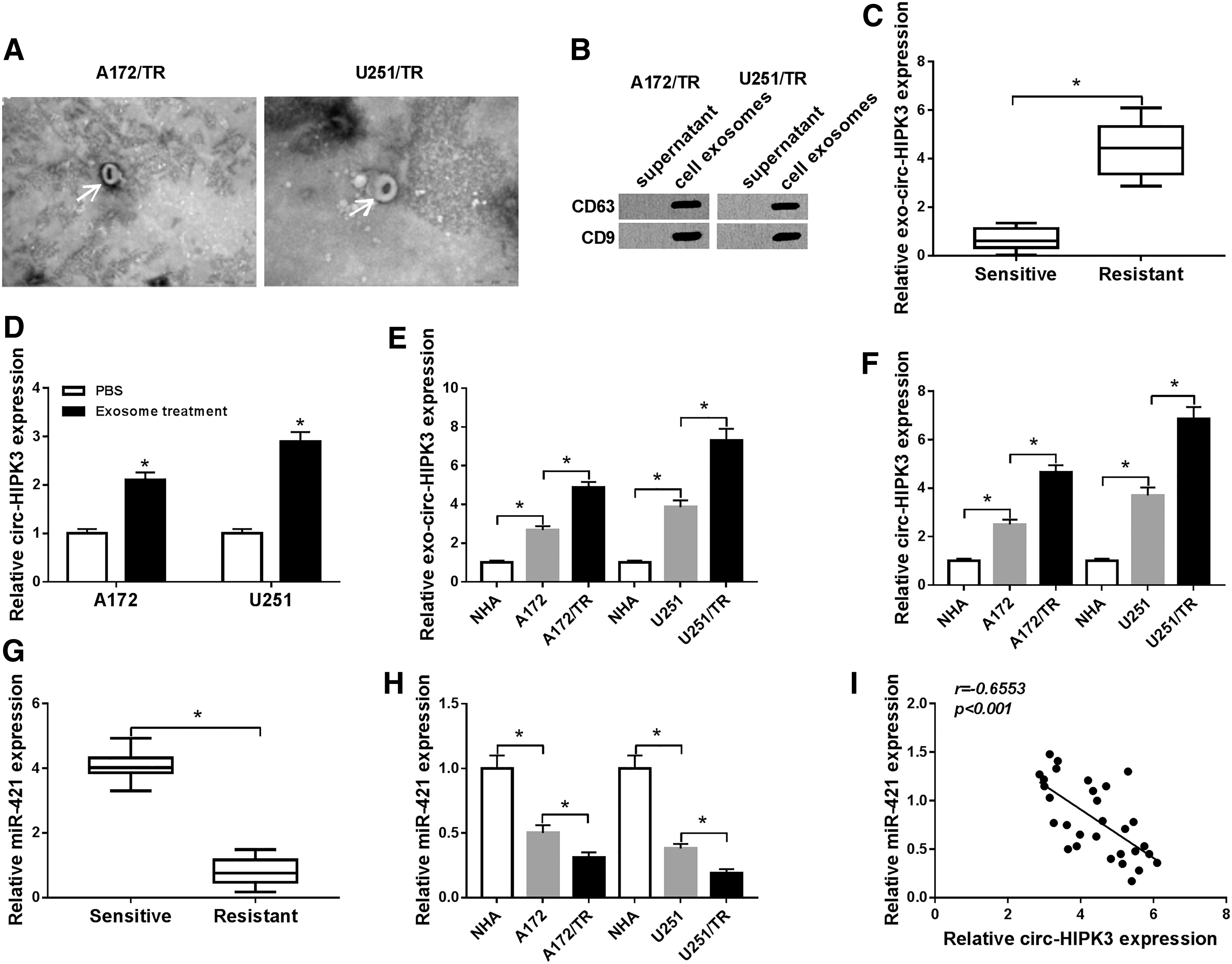
circ-HIPK3 was elevated and miR-421 was decreased in TMZ-resistant glioma cells.
Knockdown of circ-HIPK3 suppressed cell progression and TMZ resistance in TMZ-resistant glioma cells
To evaluate the function of circ-HIPK3, circ-HIPK3-silencing experiments were performed. First, they measured the IC50 of TMZ using MTT assay. Compared with the parental cells (A172 and U251), the IC50 of TMZ in TMZ-resistant cells (A172/TR and U251/TR) was enormously elevated (Fig. 2A). Then they transfected si-circ-HIPK3 into A172/TR and U251/TR cells to knock down circ-HIPK3, and circ-HIPK3 expression was remarkably decreased relative to the negative controls (Fig. 2B). After circ-HIPK3 was silenced, the IC50 of TMZ of A172/TR and U251/TR cells also reduced (Fig. 2C). Besides, flow cytometry results showed that the apoptosis rate of A172/TR and U251/TR cells were significantly increased by transfection of circ-HIPK3 (Fig. 2D). Transwell assay demonstrated that the invasive capacity of A172/TR and U251/TR cells was repressed by silencing circ-HIPK3 (Fig. 2E). P-gp, MRP1, and MDR1 have been identified as drug resistance markers in cancer, 25,26 our results showed that circ-HIPK3 knockdown in A172/TR and U251/TR cells obviously restrained their expression levels by Western blot (Fig. 2F, G). In brief, the above data supported that knockdown of circ-HIPK3 reduced the IC50 value of TMZ, inhibited cell invasion, TMZ resistance, and induced apoptosis in TMZ-resistant glioma cells.

Knockdown of circ-HIPK3 suppressed cell progression and TMZ resistance in TMZ-resistant glioma cells.
Silencing circ-HIPK3 enhanced TMZ sensitivity in TMZ-resistant glioma cells by negatively regulating miR-421
CircRNAs have been widely reported to exert their function as miRNA sponges. 27 As presented in Figure 3A, the potential binding sites between circ-HIPK3 and miR-421 were predicted by StarBase v2.0. To verify this prediction, dual-reporter luciferase assay was performed in A172/TR and U251/TR cells and found that miR-421 obviously decreased the luciferase activity in WT-circHIPK3 group while not in MUT-circHIPK3 group (Fig. 3B, C). To further confirm the interaction between circ-HIPK3 and miR-421, they also performed RIP assay in A172/TR and U251/TR cells. As shown in Figure 3D, E, both circ-HIPK3 and miR-421 could be enriched by Ago2 protein. These results confirmed that miR-421 was a target miRNA of circ-HIPK3. Furthermore, they found miR-421 was increased when circ-HIPK3 was silenced in A172/TR and U251/TR cells, while miR-421 was decreased after circ-HIPK3 overexpression (Fig. 3F, G). To determine whether the antitumor effects of si-circ-HIPK3 were mediated by miR-421, they cotransfected with si-circ-HIPK3 and anti-miR-421 into A172/TR and U251/TR cells, and the transfection efficiency was measured by qRT-PCR (Fig. 3H). The results showed that transfection of anti-miR-421 reversed the inhibitory effect of si-circ-HIPK3 on IC50 of TMZ (Fig. 3I) and cell invasion (Fig. 3K), and rescued the increased apoptosis rate (Fig. 3J) induced by si-circ-HIPK3 in A172/TR and U251/TR cells. Meanwhile, the reduction effects of circ-HIPK3 knockdown on levels of P-gp, MRP1, and MDR1 were abolished by interfering with miR-421 (Fig. 3L, M). Taken together, circ-HIPK3 regulated cell progression and TMZ resistance in TMZ-resistant glioma by targeting miR-421.
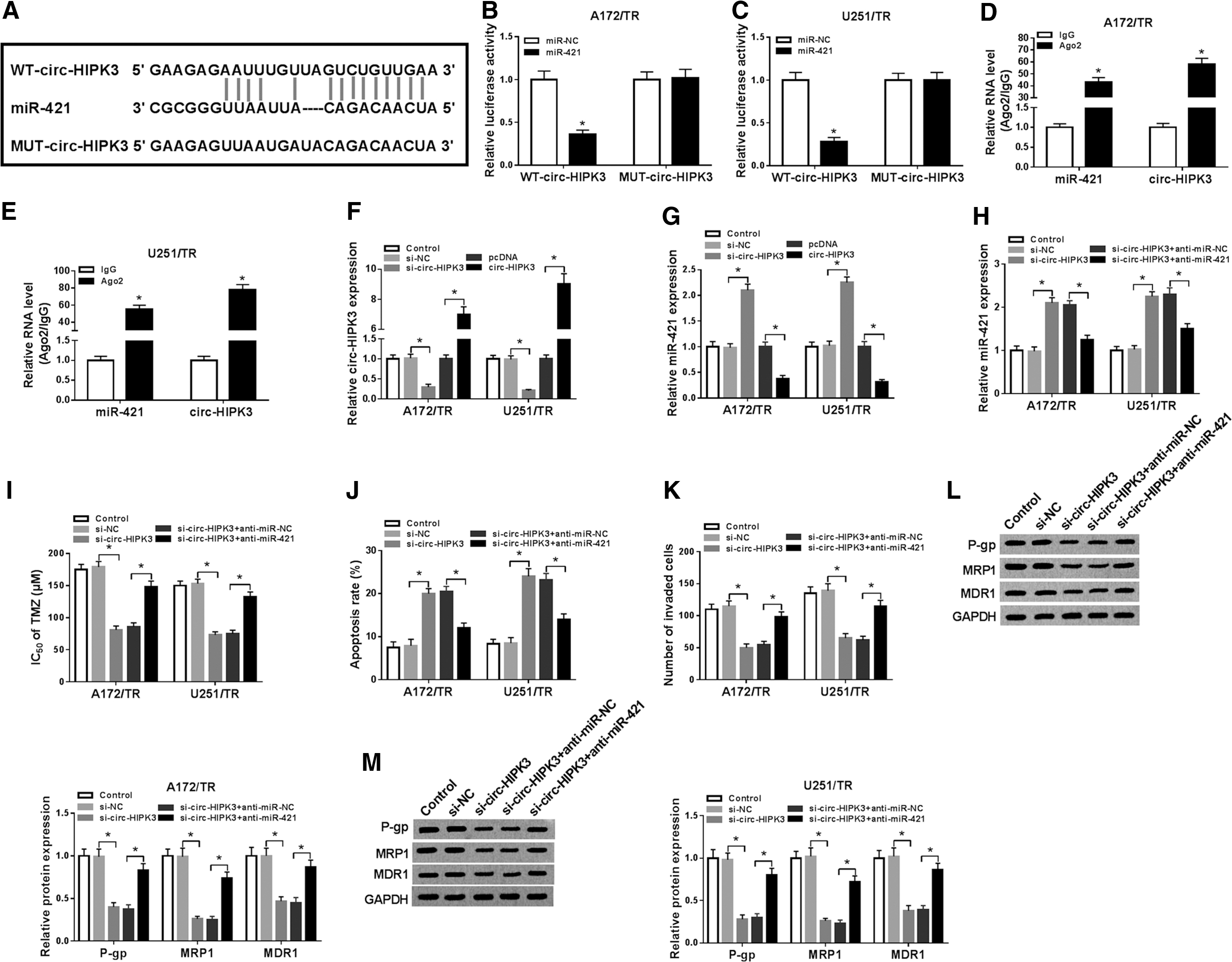
Silencing circ-HIPK3 enhanced TMZ sensitivity in TMZ-resistant glioma cells by negatively regulating miR-421.
ZIC5 was a target gene of miR-421
To investigate the mechanisms by which miR-421 regulated TMZ-resistant glioma cell progression and TMZ resistance, StarBase v2.0 tool was employed to search for the candidate targets and found that there were complementary sites between miR-421 and ZIC5 3′UTR (Fig. 4A). In the same way, dual-reporter luciferase assay showed that miR-421 overexpression evidently lowered the luciferase activity of ZIC5 3′UTR-WT report plasmid, but not the ZIC5 3′UTR-MUT report plasmid in A172/TR and U251/TR cells (Fig. 4B, C). RIP results indicated that the levels of miR-421 and ZIC5 were augmented in Ago2 immunoprecipitate compared with IgG immunoprecipitate (Fig. 4D, E). In addition, the mRNA and protein levels of ZIC5 in TMZ-resistant glioma tissues were fortified (Fig. 4F, G). As expected, the mRNA and protein levels of ZIC5 in A172 and U251 cells were upregulated relative to NHA cells, and ZIC5 expression was more strongly expressed in A172/TR and U251/TR cells (Fig. 4H, I). Notably, a distinct inverse correlation between the levels of miR-421 and ZIC5 was revealed by Pearson correlation coefficient in glioma tissues (Fig. 4J). Therefore, they concluded that miR-421 could directly target ZIC5 and negatively regulate its expression in TMZ-resistant glioma cells.
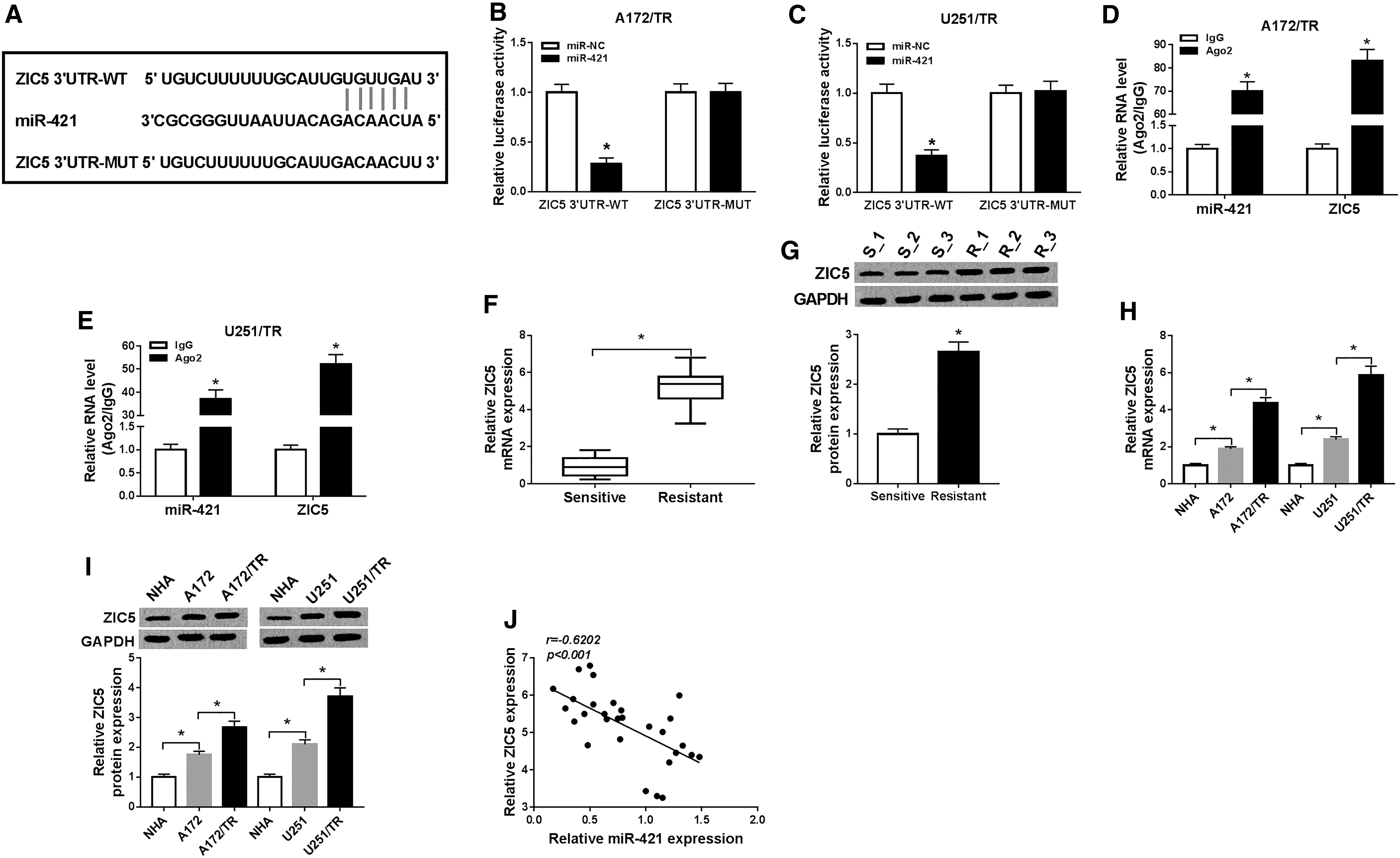
ZIC5 was a target gene of miR-421.
ZIC5 overexpression reversed the biological effects of miR-421 on TMZ-resistant glioma cells
To explore whether ZIC5 mediated the biological function of miR-421 on TMZ-resistant glioma cells, the rescue experiments were carried out. First, Western blot data showed that upregulation of miR-421 in A172/TR and U251/TR cells reduced ZIC5 protein expression, while transfection of ZIC5 counteracted the inhibition effect of miR-421 on ZIC5 expression (Fig. 5A, B). Then, they found that when miR-421 was overexpressed in A172/TR and U251/TR cells, the IC50 value of TMZ was declined (Fig. 5C), the ability of cells to invade was inhibited (Fig. 5E) and cell apoptosis rate was increased (Fig. 5D). However, overexpression of ZIC5 attenuated these effects of miR-421 on A172/TR and U251/TR cells. Furthermore, the inhibitory effects of miR-421 on levels of P-gp, MRP1, and MDR1 were weakened by overexpressing ZIC5 (Fig. 5F, G). These observations signified that miR-421 inhibited IC50 value of TMZ, cell invasion, and elevated apoptosis as well as TMZ sensitivity by regulating ZIC5.
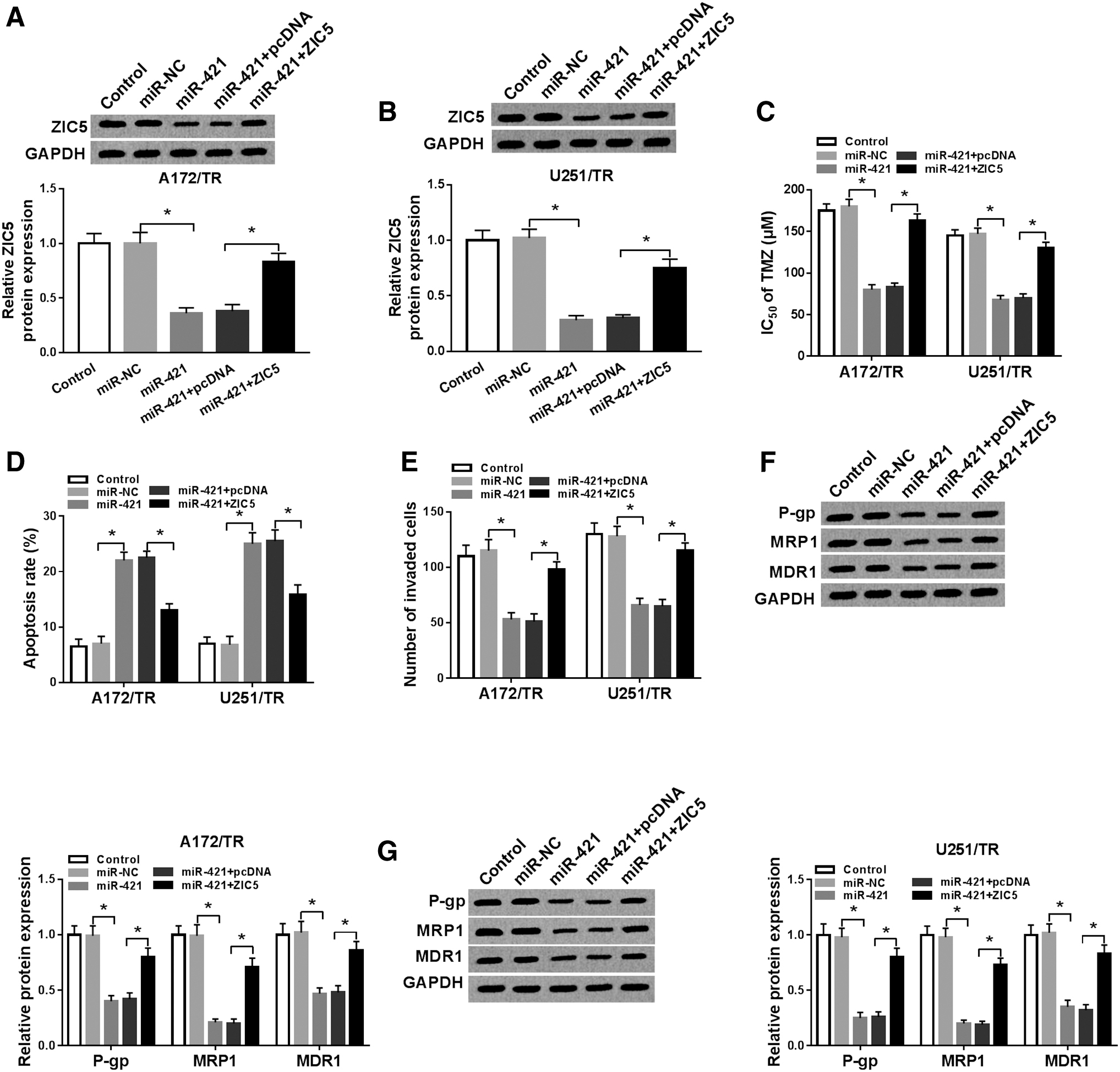
ZIC5 overexpression reversed the biological effects of miR-421 on TMZ-resistant glioma cells. After A172/TR and U251/TR cells were treated with miR-NC, miR-421, miR-421 +pcDNA, or miR-421 + ZIC5.
circ-HIPK3 actively modulated ZIC5 expression by acting as a sponge for miR-421
According to the above findings, the authors speculated that circ-HIPK3 might mediate the regulation of ZIC5 by sponging miR-421 in TMZ-resistant glioma cells. To confirm this guess, ZIC5 expression in A172/TR and U251/TR cells cotransfected with si-circ-HIPK3 and anti-miR-421 was examined by Western blot. As appeared in Figure 6A and B, repression of circ-HIPK3 reduced ZIC5 expression, whereas miR-421 deficiency could neutralize this inhibition impact. The results revealed that circ-HIPK3 could modulate ZIC5 expression by sponging miR-421 in TMZ-resistant glioma cells.
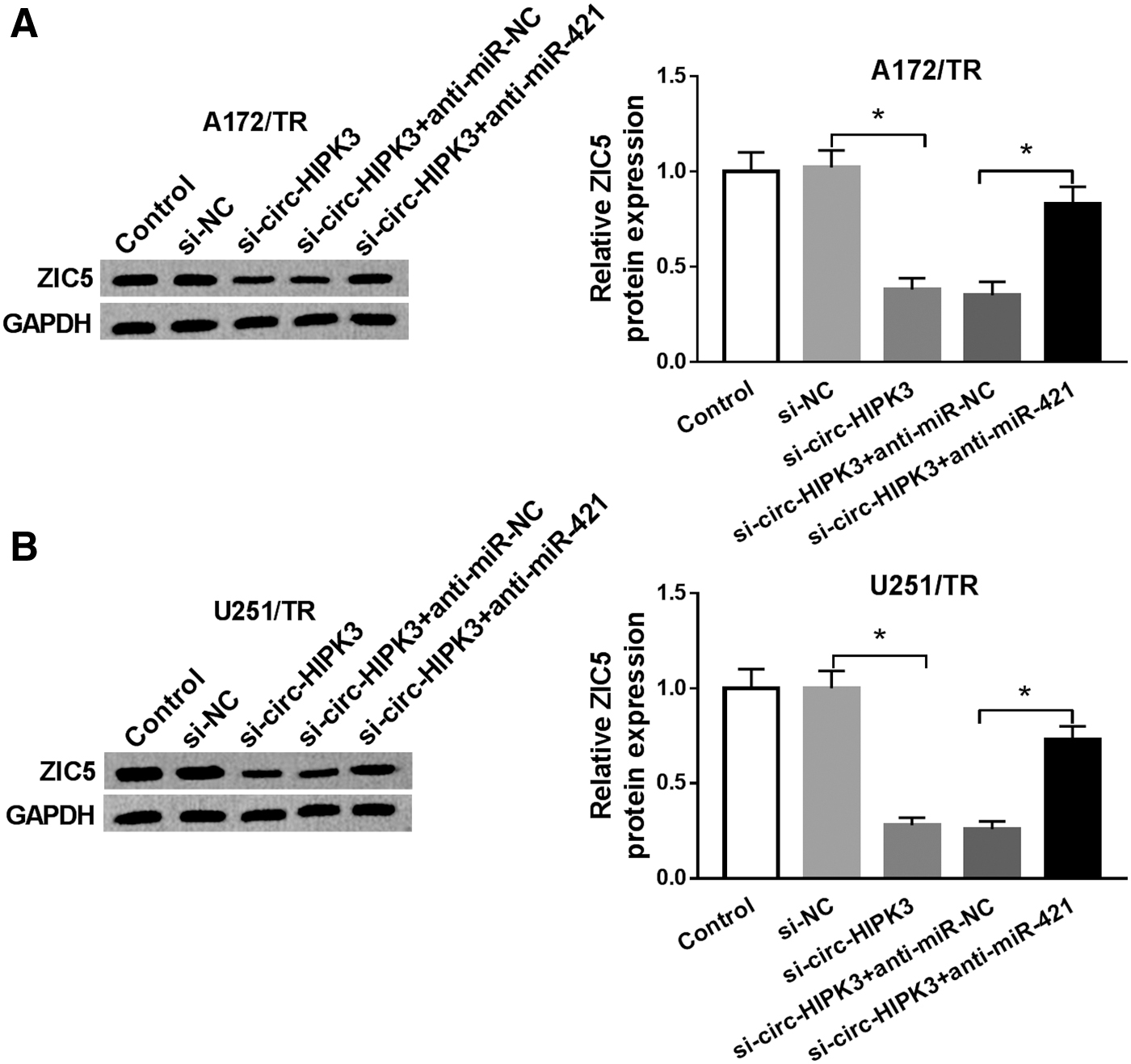
circ-HIPK3 actively modulated ZIC5 expression by acting as a sponge for miR-421.
Suppression of circ-HIPK3 expression hampered tumor growth in vivo
Subsequently, to further assess the effect of circ-HIPK3 on tumor growth in vivo, they established the xenograft mouse models, and U251/TR cells that stably knocked down circ-HIPK3 were inoculated into nude mice. The results were consistent with those of in vitro experiments; circ-HIPK3 silence group showed smaller tumor size and weight versus that the control group (Fig. 7A, B). Moreover, circ-HIPK3 expression (Fig. 7C) and ZIC5 protein expression (Fig. 7E) were downregulated in sh-circ-HIPK3 + TMZ group, while miR-421 was enhanced (Fig. 7D), suggesting that silencing circ-HIPK3 inhibited tumor growth by regulating the levels of miR-421 and ZIC5 in TMZ-resistant glioma.
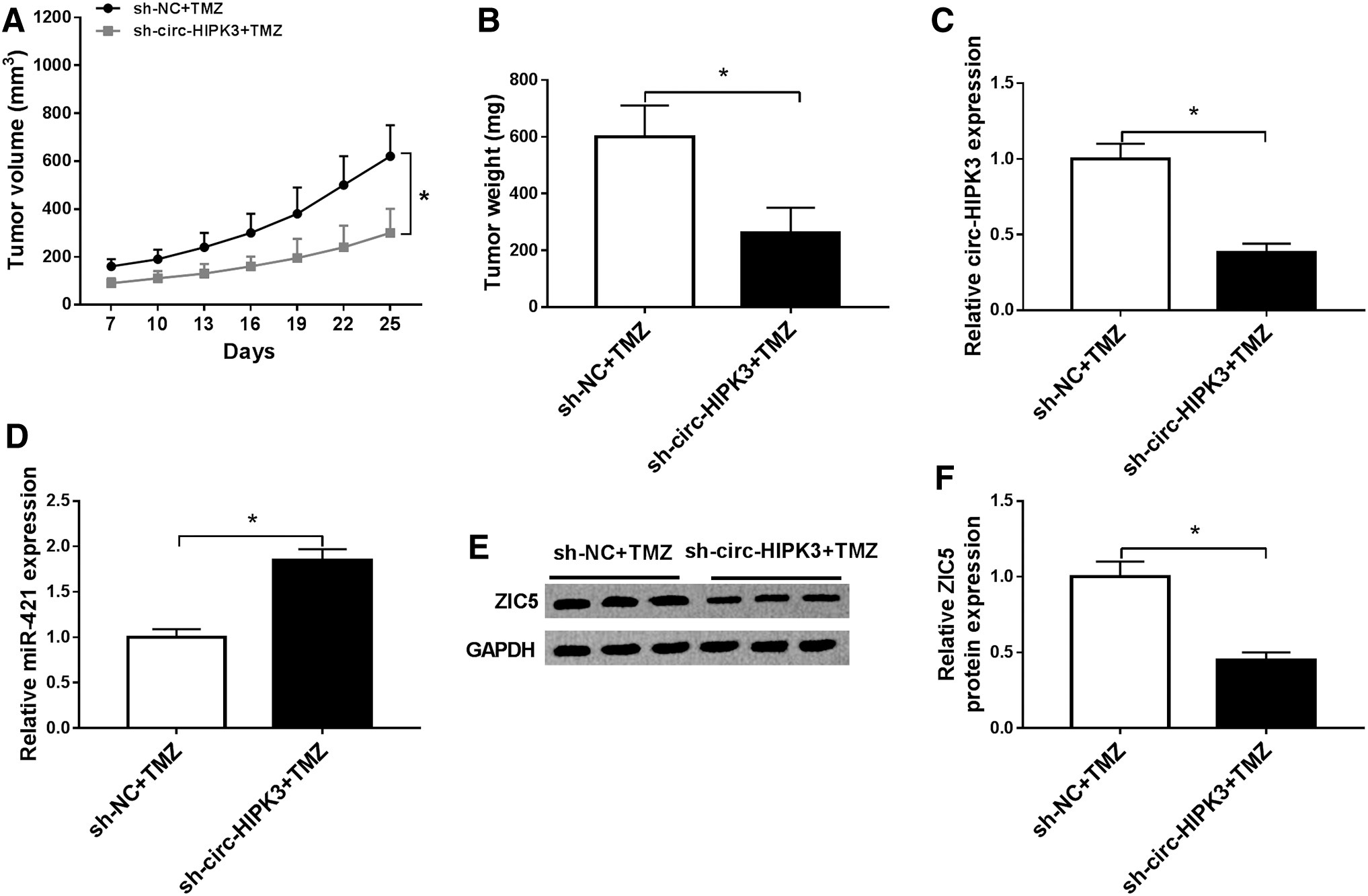
Suppression of circ-HIPK3 expression hampered tumor growth in vivo. U251/TR cells stably knocked down circ-HIPK3 were inoculated into nude mice.
Discussion
Recently, the research on exosome function has attracted wide attention, their secretions, including circRNAs, not only regulate the progression of various tumors, but also reflect cell resistance to TMZ in glioblastoma. 28 In this study, they validated that circ-HIPK3 expression in exosomes of TMZ-resistant glioma patients was higher than TMZ-sensitive glioma patients, and circ-HIPK3 expression in TMZ-resistant glioma cells A172/TR and U251/TR was significantly increased compared with A172 and U251 cells. Besides, circ-HIPK3 knockdown repressed the IC50 value of TMZ, cell invasion, and induced TMZ sensitivity and cell apoptosis in TMZ-resistant glioma cells. In accordance with previous data, Hu et al. found that circ-HIPK3 could promote the progression of glioma cells. 29 These results indicated that exosome circ-HIPK3 secreted by TMZ-resistant glioma cells could promote cell development and increase TMZ resistance.
circ-HIPK3 has previously been reported to facilitate glioma progression by regulating IGF2BP3 through sponging miR-654. 29 The authors hypothesized whether circ-HIPK3 could act as a sponge of miRNA in this study to regulate the progression of TMZ-resistant glioma cells. Coincidentally, the lowly expressed miR-421 in TMZ-resistant glioma tissues and cells was proved to be the target miRNA of circ-HIPK3, and their expression levels were negatively correlated in TMZ-resistant glioma tissues. Furthermore, silencing miR-421 could reverse the effects of si-circ-HIPK3 on IC50 value of TMZ, invasion, apoptosis, and TMZ resistance of TMZ-resistant glioma cells, which was in agreement with previous results. 30 Previously, miR-421 was thought to exert its function by acting as an oncogene in osteosarcoma 31 and non-small cell lung cancer. 18 In addition to glioma, miR-421 has been identified as a tumor suppressor for breast cancer, 32 and they suspected that the different expression patterns of miR-421 might be caused by different types of cancer. In a word, our results implied that circ-HIPK3 served as an oncogene partially by sponging miR-421 in TMZ-resistant glioma cells.
Then, they investigated the downstream signal of miR-421 and confirmed ZIC5 was the target for miR-421 by dual-reporter luciferase and RIP assays. Li et al. demonstrated that upregulation of ZIC5 triggered proliferation and migration of glioma cells, 33 and as a mediator of melanoma drug resistance, ZIC5 could promote tumor progression and drug resistance. 34 Our data showed that ZIC5 was upregulated in TMZ-resistant glioma, and ZIC5 overexpression could counteract the inhibitory effects of miR-421 on cell progression and TMZ resistance. Importantly, circ-HIPK3 depletion could directly decrease ZIC5 expression by sponging miR-421. These results suggested that circ-HIPK3 might induce the development of TMZ-resistant glioma cells in vitro by regulating miR-421/ZIC5 axis. Meanwhile, suppression of circ-HIPK3 restrained TMZ-resistant tumor growth by regulating the levels of miR-421 and ZIC5.
Conclusion
Exosome-mediated circ-HIPK3 could enhance TMZ resistance and promote tumorigenesis in vitro and in vivo by regulating the miR-421/ZIC5 axis. Additionally, the expression of circ-HIPK3 in human serum exosomes could be used as a biomarker for diagnosis of TMZ-resistant glioma patients.
Footnotes
Acknowledgment
The authors sincerely appreciate all members who participated in this study.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.