
Abstract
Background:
Melanoma is one of the most aggressive malignancies. Exploration of metastasis-related genes will improve the clinical outcomes of patients with melanoma. Recently, microRNAs (miRNAs) have been implicated in regulating the aggressiveness of melanoma. In the current study, the author demonstrated the expression of miR-548b and its functions in melanoma.
Materials and Methods:
The expression levels of miR-548b and high mobility group protein 1 (HMGB1) in melanoma specimens and adjacent normal tissues were examined using the quantitative real-time PCR method. The Cell Counting Kit-8 (CCK-8), wound healing test, and Transwell assays were conducted to examine the impact of miR-548b on aggressive phenotypes of melanoma cells. The protein expression of HMGB1 was detected by Western blot. The tumor growth of melanoma cells in vivo was analyzed using the transplanted tumor model. The expression of HMGB1 in vivo was assessed using immunohistochemistry assay.
Results:
miR-548b was significantly downregulated in the melanoma sample when compared with adjacent normal tissues. In addition, low levels of miR-548b were related to poor overall survival in patients with melanoma. As predicted, overexpression of miR-548b suppressed the growth and metastasis-associated traits of melanoma cells. Furthermore, the luciferase reporter gene assay and Western blotting revealed that HMGB1 was a target of miR-548b and its expression level was negatively modulated by miR-548b. Several rescue experiments indicated that reintroduction of HMGB1 abolished the inhibiting effects of miR-548b on melanoma cells. Finally, the author demonstrated that upregulation of miR-548b repressed melanoma cell growth in vivo.
Conclusions:
All these findings demonstrate that miR-548b serves as a cancer-suppressive miRNA in human melanoma by inhibiting HMGB1.
Introduction
Melanoma has increased in frequency in light-skinned populations worldwide and currently reaches an incidence (new cases per 100,000 inhabitants per year) of 10–25 cases in North America, 10–20 in Central Europe, and 50–60 in Australia. The lifetime risk for developing melanoma is around 2% (1:53). 1 –5 The survival rates of patients with advanced melanoma remain low; specifically, the 5-year survival rate in patients with stage IV melanoma is ≈6%. 6,7 Although most melanoma cases consist of localized disease that is curable with surgical resection, for those patients with regional or distant metastases, the prognosis is poor. 8 Hence, it is necessary to investigate the precise mechanisms behind melanoma progression and identify potential therapeutic targets. 9 –11
An increasing number of experimental studies have proved that microRNAs (miRNAs) exert cancer-suppressive roles or oncogenic activities in cancer and regulate a great variety of biological phenomena, including growth, cell apoptosis, epithelial–mesenchymal transition (EMT), and metastasis. 12 –15 In recent years, the therapeutic application of miRNAs in cancer is an exciting approach to interfere with the molecular mechanism of malignancies. The first possible application of miRNAs is based on reintroduction of single or multiple miRNA mimics (non-natural, double-stranded, miRNA-like RNA fragments) into a group of cancer cells, thus trying to reestablish the normal expression profile and restoring loss of function. 16,17 Another strategy for application of miRNAs as therapeutic tools is to inhibit oncogenic miRNAs by using the mRNA antagonists. Several investigations have explored miRNA expression profiling in melanoma and revealed that dysregulation of miRNAs is associated with progression of malignant melanoma. 18,19 In melanoma, miR-98 represses the metastatic ability of melanoma cells through a negative feedback loop with its target gene, interleukin 6 (IL-6). 20 Furthermore, downregulated miR-23a contributes to the metastasis of cutaneous melanoma by promoting autophagy. 21 miRNA-205 suppresses proliferation and induces senescence of melanoma cells by regulating E2F1 (E2F1). 22 In addition, miR-659-3p has been identified as a candidate predictive biomarker for patients with melanoma who were treated with platinum-based chemotherapy. 17 In the highly aggressive melanoma cell line, A375, miRNA-3662 targets zinc finger E-box binding homeobox 1 (ZEB1) and attenuates the invasion of melanoma cells in vitro. 23
Recently, miRNA-548b has been found to inhibit the aggressiveness of hepatocellular carcinoma (HCC) by regulating the high mobility group protein 1 (HMGB1). 24 In addition, miR-548b restrains the growth and aggressiveness of glioma by targeting metastasis tumor-associated protein-2 (MTAP2). 25 miR-548b is also expressed at a low level in HCC tissues and might serve as a tumor-suppressive miRNA in proliferation and invasion of HCC cells. 26 Nevertheless, the potential mechanisms behind miR-548b in regulation of melanoma development need to be deeply explored. HMGB1 is a nuclear nonhistone protein and implicated in cancerogenesis. 27 For instance, HMGB1 promotes prostate cancer cell metastasis by interacting with Brahma-related gene 1 and activating the AKT signaling pathway. 28 Higher expression of HMGB1 is closely associated with worse overall survival (OS) in thyroid cancer patients with lymph node metastasis. 29 A high level of HMGB1 is associated with the advanced stages and poor survival in patients with melanoma. 30 Several HMGB1-targeting agents have been used in experimental anticancer investigations. These agents include soluble RAGE (sRAGE), HMGB1-neutralizing antibody, A box protein, platinating agent, ethyl pyruvate, quercetin, and glycyrrhizin. 31 sRAGE functions as a decoy to prevent RAGE signaling and has been used successfully in blocking the HMGB1-RAGE signaling axis in animal tumor models. 32 Ethyl pyruvate, the first HMGB1 inhibitor used in animal models of sepsis by inhibition of the NF-kB pathway, inhibits liver tumor growth. 33 Additionally, glycyrrhizin and quercetin (potential HMGB1 inhibitors), by directly binding to HMGB1 or inhibition of PI3K, improve the effectiveness of anticancer agents in several cancer models. 34,35
In this study, the author demonstrates that miR-548b is strikingly downregulated in clinical melanoma samples and melanoma cell lines. Subsequent functional experiments reveal that miR-548b inhibits cell growth and the invasive abilities of melanoma cells in vitro. Thereafter, the author further finds that miR-548b suppresses the invasive abilities of melanoma cells by targeting HMGB1. Finally, the author proves that miR-548b prevents melanoma cell growth in vivo. Altogether, these findings imply that miR-548b restrains the progression of melanoma cells through the miR-548b-HMGB1-negative feedback loop.
Materials and Methods
Clinical tissues
Twenty-three pairs of melanoma tissues and adjacent samples were obtained from patients, from 2012 to June 2018, at the Yantai Yuhuangding Hospital affiliated to Qingdao University. This study was approved by the Ethics Committee (No: 20181109) of Yantai Yuhuangding Hospital affiliated to Qingdao University. Written informed consent was obtained from all participants involved in the study. The clinicopathological parameters in patients with melanoma are summarized in Table 1.
Relationship Between the Expression of miR-548b and Clinicopathological Parameters in Patients with Melanoma
Cell culture
Melanoma cell lines (A375, B16F10, B16BL6, and SK-MEL-2) and the human keratinocyte cell line, HaCaT, were bought from Nanjing KeyGen Biotech. Co., Ltd. (Nanjing, Jiangsu, China). Cells were cultured using DMEM (Thermo Fisher Scientific) containing 10% fetal bovine serum (FBS) (Thermo Fisher Scientific) at 37°C with 5% CO2.
miRNA expression profile data from Gene Expression Omnibus and The Cancer Genome Atlas
miRNA data (GSE20994) were downloaded from Gene Expression Omnibus (GEO) databases (
Cell transfection
miR-548b mimic, miR-548b inhibitor, and negative control (miR-NC or miR-NC inhibitor) were bought from RiboBio (Guangzhou, Guangdong, China). Cells were transfected with miR-548b or miR-548b inhibitor using Lipofectamine RNAiMAX (Thermo Fisher Scientific) for 24 h. The HMGB1 full-length plasmid and respective nontargeting sequence (negative control, NC) were synthesized by GenePharma (Shanghai, China).
Luciferase reporter gene assay
The fragment of 3′-UTR HMGB1 containing miR-548b binding sites (Wt) or fragment containing the site modified by site-directed mutagenesis (Mut) was inserted into a psiCHECK-2.0 vector (Promega). A375 or SK-MEL-2 cells were transfected with the miR-548b combination with Wt or Mut plasmid. Forty-eight hours post-transfection, the luciferase activity was measured using a luciferase reporter assay system (Promega).
Quantitative real-time PCR assay
Total RNAs were isolated using the TRIzol kit (Life Technologies Corporation). cDNA from miRNAs was generated using a TaqMan miRNA reverse transcription kit (Applied Biosystems, Foster City, CA). The One-Step SYBR PrimeScript RT-PCR Kit (Takara Bio, Inc., Japan) was selected to detect levels of miR-548b and HMGB1. Quantitative real-time PCR (qRT-PCR) was conducted using TaqMan Universal Master Mix II with TaqMan miRNA assays of miR-548b and U6. U6 or GAPDH was used as the endogenous control. To calculate levels of miR-548b and HMGB1, the 2−ΔΔCT method was used. The primers used are listed as follows: miR-548b forward, 5′-ACACTCCAGCTGGGCAAAAATCTCAAT-3′, and reverse, 5′-CTCAACTGGTGTCGTGGAAACTGGTGTC-3′; U6 forward, 5′-CTCGCTTCGGCAGCACAGCTTCGGCAGCACA-3′, and reverse, 5′-AACGCTTCACGAATTTGCGTCGCTTCACGAATT-3′; HMGB1 forward, 5′-TATGGCAAAAGCGGACAAGG-3′, and reverse, 5′-CTTCGCAACATCACCAATGGA-3′; and GAPDH forward, 5′-GGGCAGTATGTTGT-3′, and reverse, 5′-GCTATTGGCATTGGTGAA-3′.
Immunoblotting assay
Total proteins were extracted using RIPA buffer (Beyotime, Nanjing, Jiangsu, China). Thirty micrograms of protein lysates was subjected to 8% SDS-PAGE and transferred onto polyvinylidene difluoride (PVDF) membranes. After that PVDF membranes were blocked using 5% nonfat milk and then incubated with HMGB1 antibody at 4°C (Proteintech, Rosemont, IL). Then, PVDF membranes were incubated with HRP-conjugated secondary antibody (Cell Signaling Technology, Danvers, MA). Peroxidase activity was detected using an ELC kit (Bio-Rad Laboratories, Hercules, CA).
Cell proliferation assay
A375 or SK-MEL-2 cells (1 × 103) were seeded into 96-well plates and cultured for 24, 48, 72, or 96 h. Ten microliters of the Cell Counting Kit-8 (CCK-8) solution (Dojindo Laboratories, Osaka, Japan) was added to the 96-well plate. After a 2-h incubation, the optical density was determined at 450 nm using a microplate reader.
Migration assay
A375 or SK-MEL-2 cells were cultured in six-well plates and the confluent monolayers were wounded using a sterile pipette tip. The cells were cultured using FBS-free culture medium. Wound closure was analyzed in 48 h under an inverted microscope.
Transwell invasion assay
The invasion of cells was detected using a Transwell chamber with 8-μm pore size membrane filter inserts (Corning) coated with Matrigel (BD Biosciences). Two hundred microliters of 5 × 103 A375 or SK-MEL-2 cells was added into the upper chamber and the bottom chambers were filled with 600 μL of medium with 10% FBS. After 48 h, the invading cells were counted in five images of each membrane under an inverted microscope.
Colony formation assay
A375 or SK-MEL-2 cells were placed into six-well plates (1000 cells/well) and incubated for 2 weeks. After that cell colonies were fixed with methanol and stained with crystal violet (Sigma). The number of cell colonies was counted under an inverted microscope.
In vivo study
miR-NC or miR-548b-transfected A375 cells were suspended in 100 μL of FBS-free DMEM (4 × 106) and injected subcutaneously into the right flank of BALB/C-nu nude mice (n = 6 in each group). Tumor diameters were detected every week and tumor volumes were calculated. After 35 d, all nude mice were sacrificed. Tumors were removed and weighed. Tumor volume was calculated using the following formula: volume = 0.5 × length × [width] 2 . All animal experiments complied with ethical regulations and were approved by the Yantai Yuhuangding Hospital affiliated to Qingdao University.
Statistical analysis
Data are expressed as the mean ± standard deviation. Statistical analysis was performed with GraphPad Prism software using Student's t-test or the nonparametric Mann–Whitney U test. 36 The relationship between HMGB1 and miR-548b was analyzed using Pearson's correlation analysis; p < 0.05 was considered statistically significant.
Results
miR-548b is downregulated in melanoma
To analyze the expression of miR-548b in collected human melanoma tissues, the author first performed qRT-PCR analysis and observed that miR-548b was significantly downregulated in melanoma tissues (n = 23) (Fig. 1A). The author then divided patients into low miR-548b and high miR-548b groups using the median value of 0.094 as a cutoff value. As shown in Table 1, the miR-548b expression level was irrelevant to age, gender, and tumor subtype, but the low level of miR-548b was more prominent in metastatic melanoma (n = 9) compared with primary melanoma (n = 14) (Fig. 1B). Then, the author analyzed the differentially expressed miRNAs (DEMs) by retrieving the microarray data in the GEO dataset (GSE20994). Upregulation of miRNAs and downregulation of miRNAs are shown in the volcano plot (Fig. 1C). Cluster analysis indicated the DEMs between normal tissues and melanoma tissues (Fig. 1D). The expression of miR-548b in GSE20994 data was normalized using the logarithm (Fig. 1E). Furthermore, Kaplan–Meier survival analysis was conducted to determine the prognostic value of miR-548b in melanoma. The mean value of miR-548b (2.30) was used to separate patients into the high expression group (n = 40) and low expression group (n = 40) in TCGA datasets. As shown in Figure 4G, patients who had a higher level of miR-548b exhibited better OS. Finally, the author found that the level of miR-548b was lower in a panel of melanoma cell lines when compared with HaCaT (Fig. 1G). These observations verified that miR-548 was downregulated in melanoma and dysregulation of miR-548b might be associated with the prognosis of melanoma.
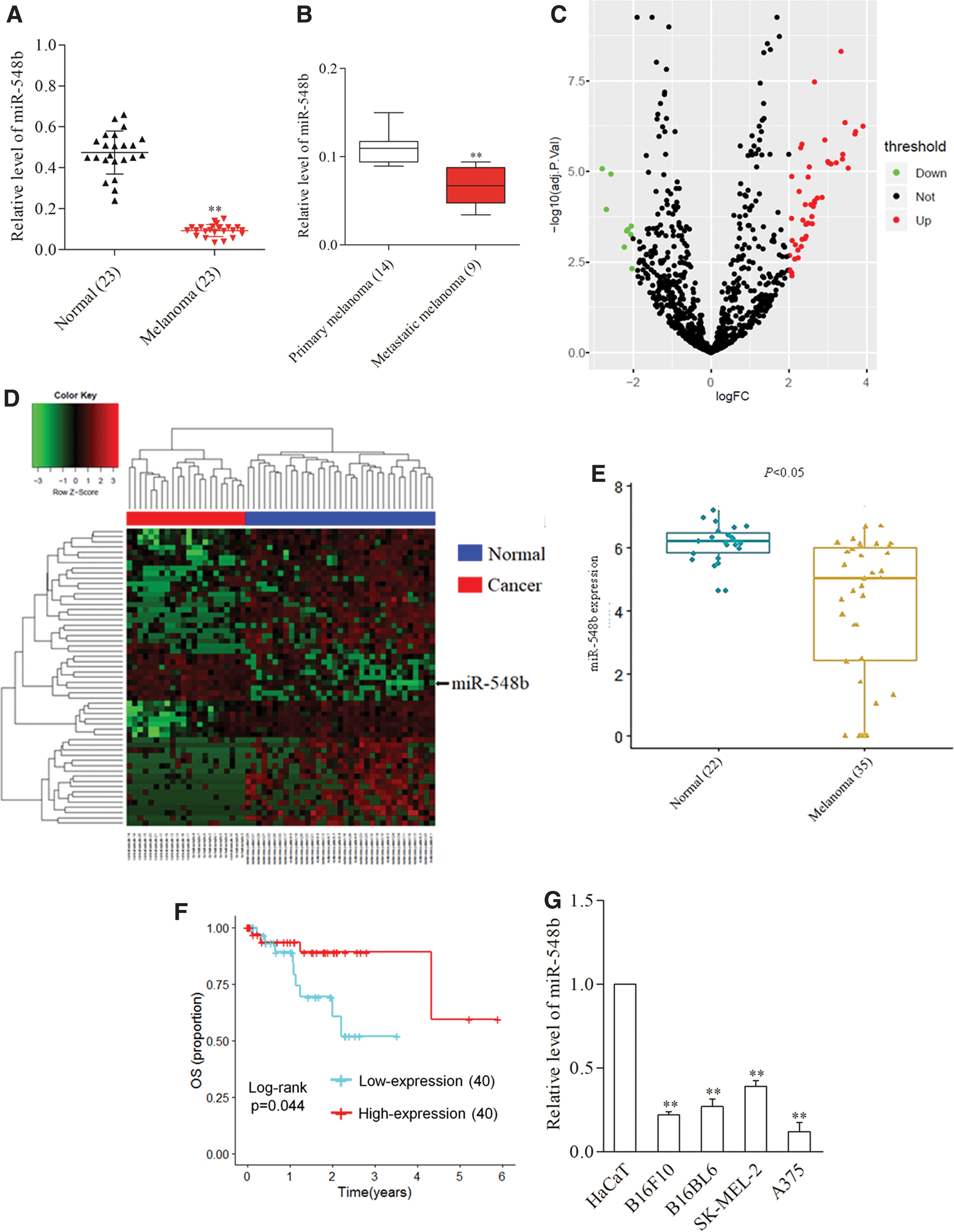
miR-548b is downregulated in melanoma.
miR-548b suppresses melanoma cell growth in vitro
To reveal the roles of miR-548b in melanoma, A375 cells were transfected with miR-548b and SK-MEL-2 cells were transfected with the miR-548 inhibitor. The qRT-PCR assay was carried out to verify transfection efficiency (Fig. 2A). Next, the author observed that miR-548b inhibited A375 cell proliferation in the CCK-8 assay (Fig. 2B), whereas the miR-548b inhibitor enhanced proliferation of SK-ME-2 cells (Fig. 2C). Additionally, colony formation assays indicated that transfection of miR-548b decreased the colony-forming ability of A375 cells, whereas the miR-548b inhibitor promoted colony formation of SK-MEL-2 cells (Fig. 2D, E). Furthermore, Transwell and wound healing assays suggested that transfection of miR-548b attenuated the invasiveness and migration ability of A375 cells (Fig. 3A, C), whereas the miR-548b inhibitor increased migration and invasion of SK-MEL-2 cells (Fig. 3B, D), indicating that miR-548b repressed the migration capacity and invasiveness of melanoma cells in vitro. These observations implied that overexpression of miR-548b impaired the growth and metastatic traits of melanoma cells.
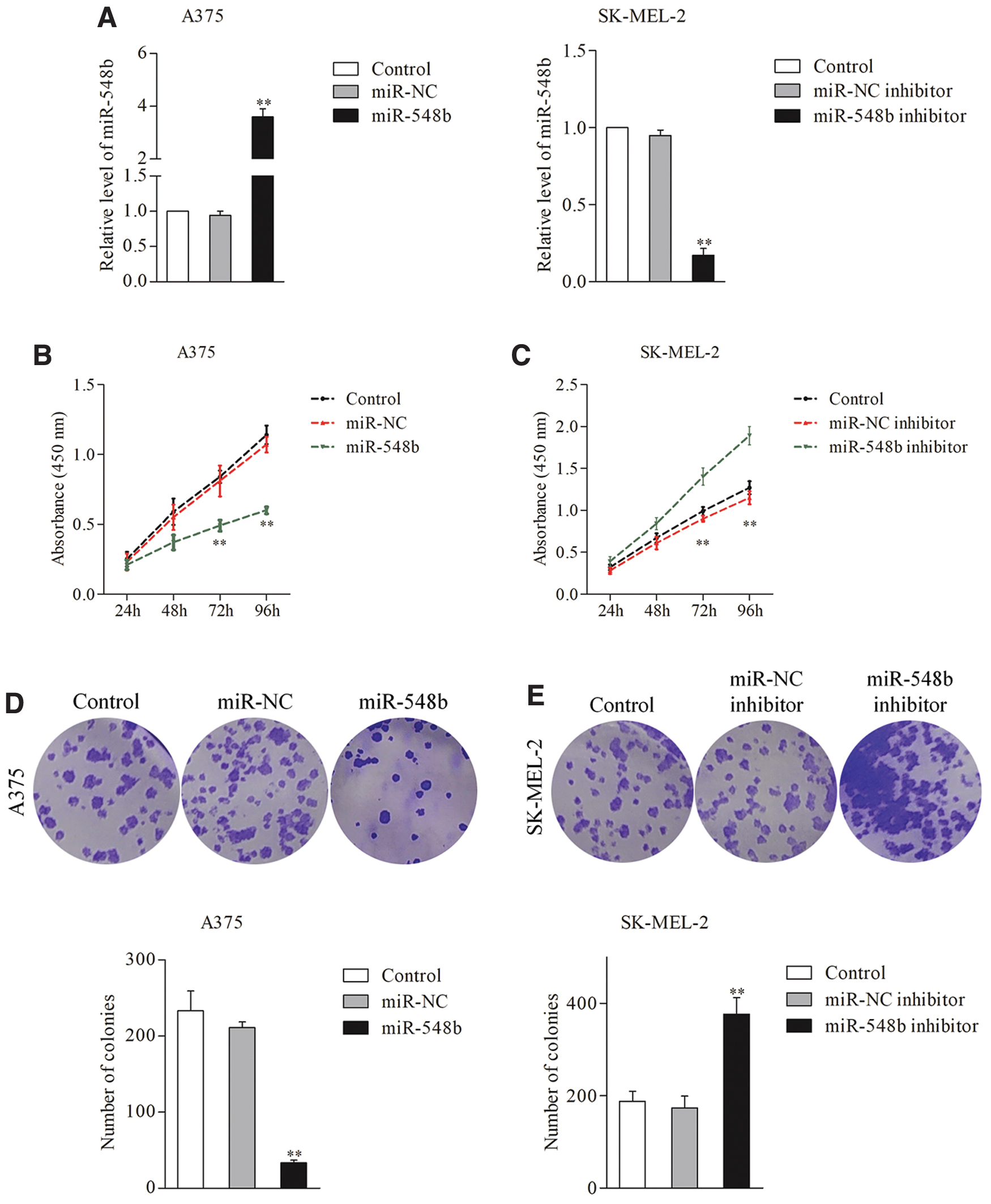
Overexpression of miR-548b inhibits growth of melanoma cells.
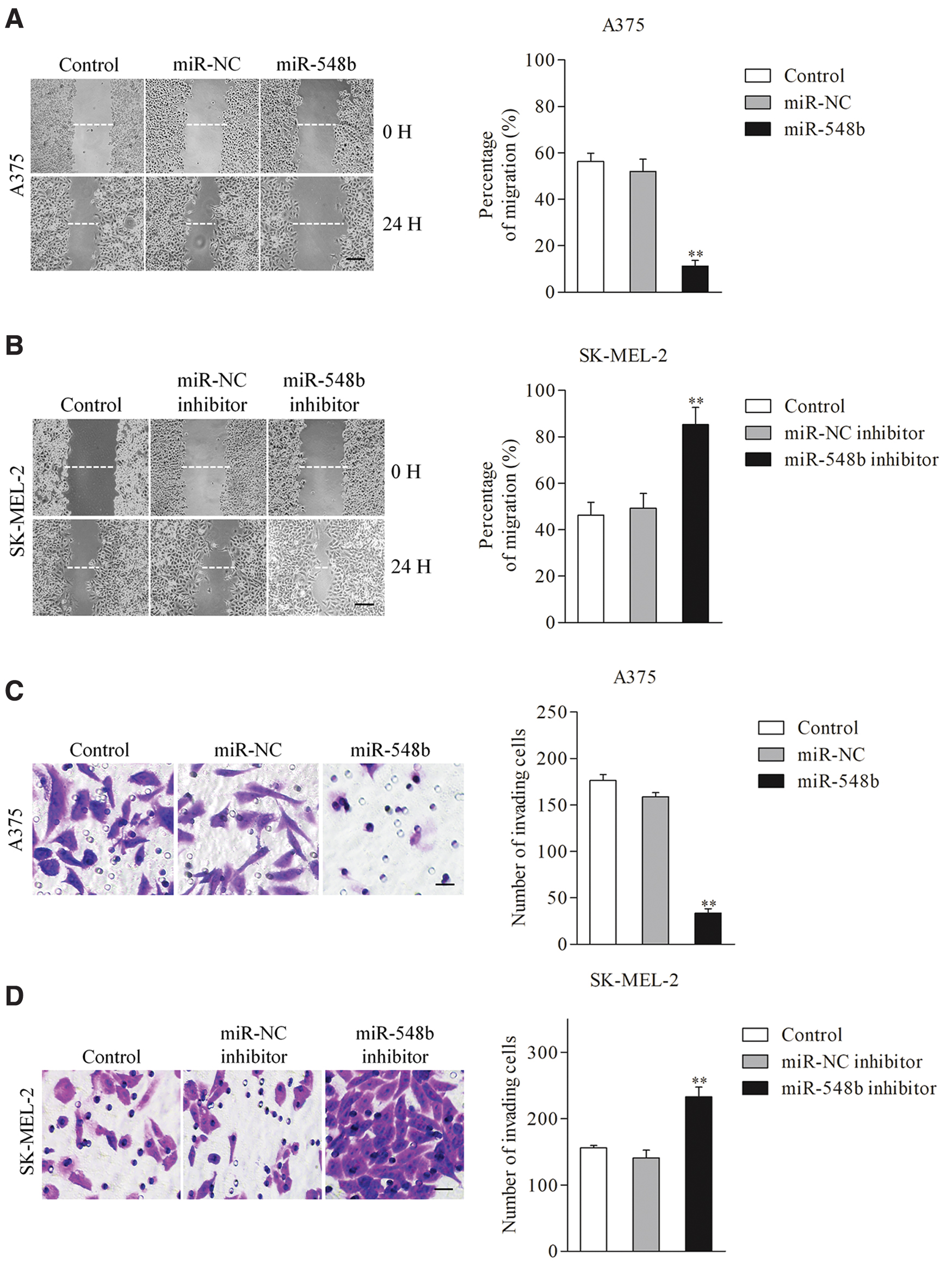
Overexpression of miR-548b inhibits migration and invasion of melanoma cells.
HMGB1 is a target of miR-548b
To explore the mechanism underlying the suppressive roles of miR-548b in melanoma cells, the online website TargetScan (

miR-548b directly regulates HMGB1.
miR-548b inhibits melanoma cells by targeting HMGB1
Because HMGB1 is a target of miR-548b, the author speculated that HMGB1 was involved in the regulatory function of miRNA in melanoma. To test this hypothesis, A375 or SK-MEL-2 cells were cotransfected with HMGB1 and miR-548b mimics. The expression levels of HMGB1 in A375 and SK-MEL-2 cells were detected using Western blotting assay (Fig. 5A). Then, the author observed that reintroduction of HMGB1 reversed miR-548b-mediated inhibition of A375 and SK-MEL-2 cell growth, colony formation, and metastasis-related phenotypes (Fig. 5B–E). These findings indicated that miR-548b modulated the invasive ability of melanoma cells by targeting HMGB1.
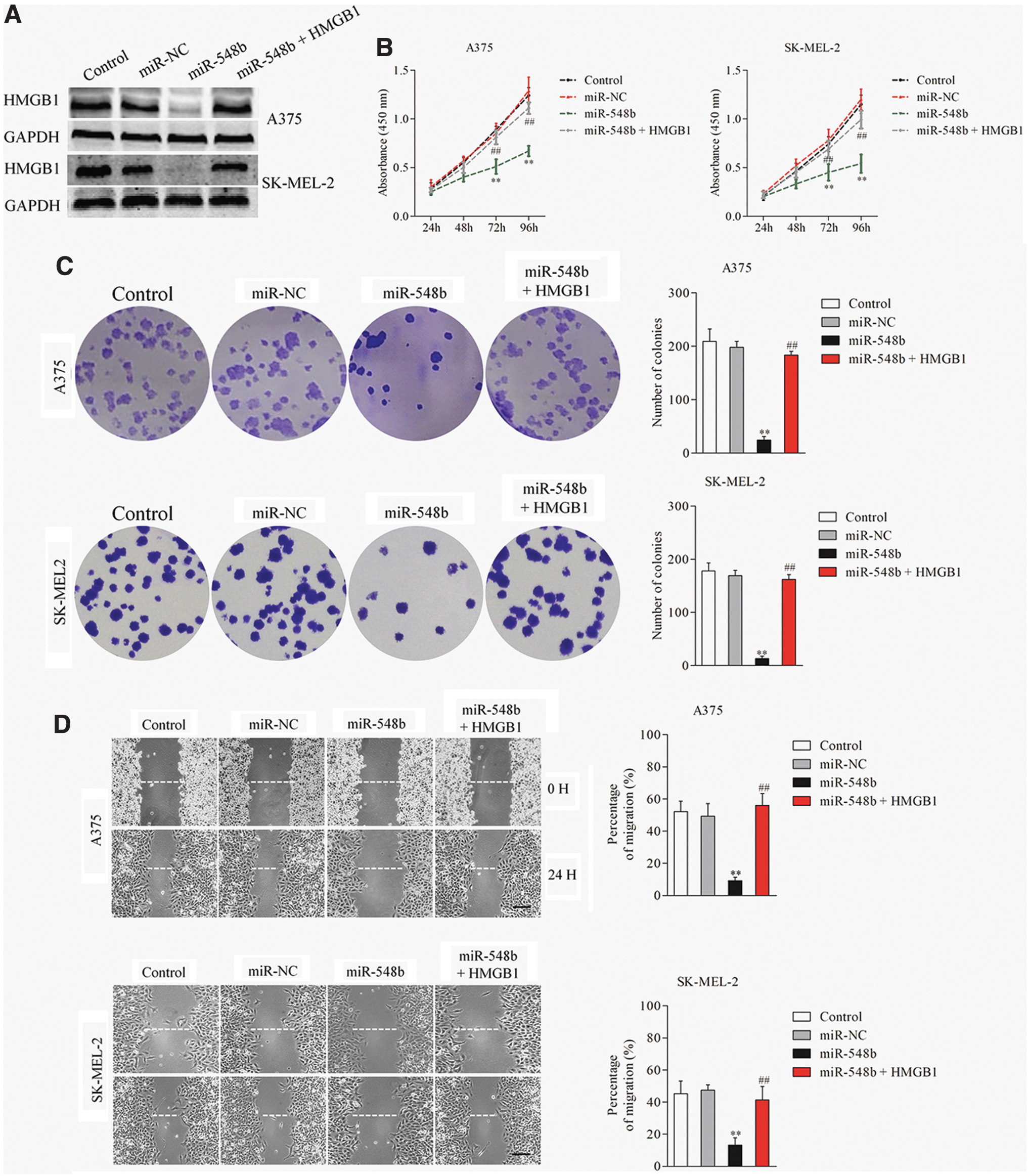
HMGB1 mediates the effect of miR-548b on melanoma.

miR-548b inhibits A375 cell growth in vivo
Finally, a transplanted tumor model was constructed to analyze the role of miR-548b in A375 cell growth in vivo. A375 cells transfected with miR-NC or miR-548b were subcutaneously inoculated into nude mice (n = 6 in each group). The tumor volume (Fig. 6A) and tumor weight (Fig. 6B) were significantly inhibited in mice that were injected with miR-548b mimic-transfected A375 cells. Additionally, the author detected levels of miR-548b in the tumor tissues (n = 3) formed by miR-NC or miR-548b-transfected A375 cells using qRT-PCR. As shown in Figure 6C, the levels of miR-548b in tumor tissues derived from miR-548b-transfected A375 cells were higher than that in the miR-NC group. After that immunohistochemistry staining and Western blot were carried out and the author found that the level of HMGB1 was markedly lower in the miR-548b group (Fig. 6D, E). These findings proved that miR-548b repressed A375 cell growth in vivo and inhibited the expression of HMGB1.

miR-548b inhibits melanoma cell growth in vivo.
Discussion
Dysregulation of miRNAs in malignant cancer is frequently reported. An increasing number of investigations have demonstrated that miRNAs are critical drivers of cancer progression, serving as suppressor genes or oncogenes. 37 –39 Hence, investigation of cancer inhibitory or oncogenic miRNAs and illumination of their functions could help identify potential biomarkers and therapeutic targets in cancers. Aberrant expression levels of miRNAs have been identified in various human malignant tumors, including melanoma. 4,5,9,40 In melanoma, miRNA-22 targets Formin-like 2 (FMNL2) to inhibit melanoma progression through regulation of the Wnt/β-catenin signaling pathway. 41 Additionally, downregulation of miR-126-3p facilitates acquired resistance to dabrafenib in melanoma cells through modulating ADAM metallopeptidase domain 9 (ADAM9) and vascular endothelial growth factor A (VEGF-A). 42 miRNA-429 suppresses proliferation and migration of melanoma cells by targeting AKT serine/threonine kinase 1 (AKT1). 43
Recently, miR-548b is identified as a cancer-associated miRNA in human squamous cell carcinoma of the tongue. 44 miRNA-548b represses the proliferation and invasion ability of HCC cells by regulating specificity protein 1 (SP1). 26 In addition, miR-548b represses the growth and invasiveness of glioma by regulating the expression of MTA2. 25 In HCC, miRNA-548b inhibits the invasive phenotypes of HCC cells by interacting with HGMB1. 24 All results point out that miR-548b plays an important role in cancer progression. However, the expression level and potential function of miR-548b in melanoma remain unclear.
In the current research, the author verified that miR-548b was distinctly downregulated in the melanoma sample when compared with adjacent tissue, using the qRT-PCR assay, indicating its possible involvement in regulating the progression of melanoma. Meanwhile, the low expression of miR-548b was also confirmed using the public GSE20994 and TCGA datasets. The author agrees that more experimental methods would be useful to confirm the low expression of miR-548b in melanoma. In a future study, the author will construct the digoxigenin LNA-modified miR-548b probe. The author hopes, in the future, to employ the miRNA in situ hybridization technique using formalin-fixed, paraffin-embedded melanoma tissues to determine the expression of miR-548b. 45 Endogenous miRNAs could be crucial for the growth and aggressiveness of cancer. 46,47 Data from previous investigations reveal that upregulation of miR-548b reduced the growth and metastatic behaviors of cancer cells. 26,48 Yun et al. have demonstrated that miRNA-548b reduces the invasive behaviors of HCC by modulating HMGB1. 24 Lin et al. performed an in vitro experiment of HCC cells and provided evidence that HCC cell growth and apoptosis are regulated by miR-548b. 48 Meanwhile, miR-548-3p is markedly downregulated in breast cancer and overexpression of miR-548-3p inhibits proliferation and promotes the apoptosis of breast cancer cells. 49 The author's in vitro experiments also indicated that miR-548b decreased the cell viability and colony-forming ability of melanoma cells, while transfection of miR-548b inhibitor caused completely opposite results. In addition, upregulation of miR-548b reduced the invasion and migration abilities of melanoma cells, whereas downregulation of miR-548b significantly increased the aggressiveness of melanoma cells.
miRNAs negatively regulate the expression levels of target mRNAs by inducing the degradation of target mRNAs or impairing their translation or both by specifically binding to the 3′-UTRs of target mRNAs. To identify the potential targets of miR-548b in melanoma, the author screened predicted targets of miR-548b using TargetScan (
Conclusions
Herein, the author proved the cancer-suppressive effects of miR-548b by targeting HMGB1, which broadened the understanding of underlying mechanisms associated with miR-548b-associated function in melanoma cells. In the current study, the author reported that HMGB1 was a downstream regulator of miR-548b in melanoma. Results indicated that miR-548b expression was downregulated in melanoma and served as a tumor-suppressive miRNA in melanoma by attenuating the expression of HMGB1.
Footnotes
Author Contributions
X.F. was involved in conception of the study and drafting the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.