
Abstract
Background:
Gastric cancer is one of the most common malignancies. Circular RNAs (circRNAs) are emerging as new players in the cancer paradigm. In the present study, the authors aimed to further explore the role and mechanism of circ_0081143 in GC progression under hypoxia.
Materials and Methods:
The expression levels of circ_0081143, miR-497-5p, and EGFR (epidermal growth factor receptor) were detected by quantitative real-time polymerase chain reaction (qRT-PCR). Cell migration and invasion abilities were determined by Transwell assay. The levels of vimentin, N-cadherin, E-cadherin, and EGFR were evaluated using Western blot. Dual-luciferase reporter assay was performed to confirm the targeted correlation between miR-497-5p and circ_0081143 or EGFR.
Results:
These data indicated that circ_0081143 expression was increased in GC tissues and cells. The knockdown of circ_0081143 relieved hypoxia-induced GC cell migration, invasion, and epithelial/mesenchymal transition (EMT). Moreover, circ_0081143 regulated the abundance of miR-497-5p through sponging miR-497-5p. The alleviative effects of circ_0081143 knockdown on GC cell migration, invasion, and EMT under hypoxia were mediated by miR-497-5p. Furthermore, miR-497-5p directly targeted EGFR, and miR-497-5p overexpression ameliorated hypoxia-induced GC cell migration, invasion, and EMT by downregulating EGFR. In addition, circ_0081143 modulated EGFR expression via acting as a competing endogenous RNA (ceRNA) of miR-497-5p in hypoxia-induced GC cells.
Conclusion:
The present work suggested that circ_0081143 knockdown ameliorated hypoxia-induced migration, invasion, and EMT in GC cells at least partly by the miR-497-5p/EGFR axis, illuminating a novel molecular target for GC treatment.
Introduction
Gastric cancer remains one of the most frequently diagnosed malignancies and the third leading cause of tumor-related deaths in 2018 globally. 1 Hypoxia, an important feature of locally advanced solid tumors, contributes to the malignant progression in human cancers, including GC. 2 Therefore, a better understanding for the molecular basis of GC progression under hypoxia is indispensable for the design of innovative treatments.
Circular RNAs (circRNAs) are a new type of noncoding transcripts that are characterized by their covalently closed-loop structures without 3′ poly-adenylated tails and 5′ caps. 3 Recently, it has become apparent that circRNAs function as vital regulators in the initiation and progression of human cancers, including GC. 4,5 For instance, Rong et al. reported that the circRNA proteasome 26S subunit (circPSMC3) was implicated in the progression of GC through the regulation of cell proliferation and metastasis. 6 Zhong et al. underscored that hsa_circ_0000993 repressed GC cell proliferation, invasion, and migration through acting as a sponge of microRNA (miRNA)-214-5p. 7 Zhang et al. highlighted that circRNA nuclear receptor interacting protein 1 (circNRIP1) acted as a tumor promoter in GC via sequestering miRNA (miR)-149-5p. 8 Interestingly, the findings by Xue et al. demonstrated that circ_0081143, derived from the back splicing of pro-α2 (I) collagen gene (COL1A2) mRNA, was highly expressed in GC tissues and its depletion weakened GC cell progression and enhanced cisplatin sensitivity in vitro and in vivo through sponging miR-646 and modulating CDK6 expression. 9 Thus, in the present study, the authors aimed to further research the impact and mechanism of circ_0081143 on GC progression under hypoxia.
MiRNAs play crucial roles in gene expression regulation and thus participate in the progression of numerous human cancers, including GC. 10 The competing endogenous RNA (ceRNA) hypothesis proposes that some circRNAs could protect against gene suppression through acting as a molecular sponge of miRNAs. 11 Previous documents had underscored that miR-497-5p functioned as a tumor suppressor to interfere with GC progression through involving in the ceRNA networks. 12 –14 More interestingly, two putative binding sites among miR-497-5p, circ_0081143, and epidermal growth factor receptor (EGFR) were identified by the starBase v.3 software, which prompted us to examine the miR-497-5p/EGFR axis as a potential molecular mediator of circ_0081143 on GC progression under hypoxia.
In the present work, the authors first explored the functional effect of circ_0081143 on GC cell migration, invasion, and epithelial/mesenchymal transition (EMT). Consequently, they investigated whether the miR-497-5p/EGFR axis was involved in the regulatory mechanism of circ_0081143 on GC cell malignant behaviors under hypoxia.
Materials and Methods
Clinical samples and ethical statement
The project set consisted of 60 tissue samples, including 30 GC tissues and 30 adjacent nontumor gastric tissues, from GC patients who underwent gastrectomy between 2015 and 2017 at Xiangyang No. 1 People's Hospital (Affiliated Hospital of Hubei University of Medicine). These samples were immediately stored in liquid nitrogen until use. All of these patients signed written informed consent, and the Ethics Committee from Xiangyang No. 1 People's Hospital (Affiliated Hospital of Hubei University of Medicine) approved all aspects of this study.
Cell culture and hypoxia treatment
Two human GC cell lines (AGS and HGC-27) and the human normal gastric epithelial (GES-1) cell line (all from Cobioer, Nanjing, China) were cultured in RPMI-1640 medium (Gibco; BRL Life Sciences Technologies, Breda, The Netherlands) containing 10% fetal calf serum (FCS, Hyclone; GE Healthcare, Buckinghamshire, UK) at 37°C with 5% CO2. For hypoxia stimulation, AGS and HGC-27 cells were cultured in a 1% O2, 5% CO2, and 94% N2 gas mixture in an incubator for various time points (0, 6, 12, 24, and 48 h) or 24 h.
Transfection of oligonucleotide and plasmid
Small interfering RNA (siRNA) targeting circ_0081143 (si-circ_0081143, 5′-GGUGAAACUGGUCUCCAUGGU-3′) and nontarget siRNA (si-NC, 5′-UUCUCCGAACGUGUCACGUTT-3′), the mature miR-497-5p mimic (5′-UGUUUGUGGUCACACGACGAC-3′) and a scrambled negative mimic (miR-NC, 5′-UCACAACCUCCUAGAAAGAGUAGA-3′), the inhibitor of miR-497-5p (anti-miR-497-5p, 5′-GUCGUCGUGUGACCACAAACA-3′) and the negative control inhibitor (anti-miR-NC, 5′-UUUGUACUACACAAAAGUACUG-3′), pcDNA-based EGFR overexpression plasmid (EGFR) 15 and pcDNA negative plasmid were synthesized by GenePharma (Shanghai, China). AGS and HGC-27 cells at 60% confluence were transfected with 50 nM of the indicated oligonucleotides or/and 100 ng of plasmids using the commercially available Lipofectamine 3000 reagent (Invitrogen, Zug, Switzerland).
Quantitative real-time polymerase chain reaction
Total RNA was isolated from tissues and cells using the Roche High Pure RNA Isolation Kit (Roche, Almere, The Netherlands) and analyzed for the concentration and purity by a spectrophotometer BioPhotometer (Eppendorf, Hamburg, Germany). One microgram of RNA was reverse transcribed into cDNA using the High-Capacity Reverse Transcription (RT) Kit (Applied Biosystems, Darmstadt, Germany) for circ_0081143 and EGFR mRNA and TaqMan MicroRNA RT Kit (Applied Biosystems) for miR-497-5p following the recommendation of manufacturers. Synthesized cDNA was then subjected to quantitative real-time polymerase chain reaction (qRT-PCR) using TaqMan gene expression assays (Applied Biosystems) on an ABI7900HT real-time PCR machine (Applied Biosystems), with β-actin or U6 as a housekeeping gene for normalization. The sequences of PCR primers were listed: circ_0081143 (sense, 5′-GGCAATATTGGTCCCGTTG-3′ and antisense, 5′-GACCAGGGAGACCAAACTCA-3′); miR-497-5p (sense, 5′-CAGCAGCACACTGTGGTT-3′ and antisense, 5′-CCAGTGCAGGGTCCGAGGTA-3′); EGFR mRNA (sense, 5′-GATGATATCATGCGACCCTCCGGGAC-3′ and antisense, 5′-ATCGATATCTCATGCTCCAATAAATTC-3′); β-actin (sense, 5′-GCACCACACCTTCTACAATG-3′ and antisense, 5′-TGCTTGCTGATCCACATCTG-3′); and U6 (sense, 5′-CTCGCTTCGGCAGCACA-3′ and antisense, 5′-AACGCTTCACGAATTTGCGT-3′).
Transwell migration and invasion assay
Transwell invasion and migration assays were implemented using Matrigel-precoated invasion chambers (8-μm pore size, Corning Costar, High Wycombe, UK) and 24-Transwell Boyden chambers (Corning Costar), respectively, as described previously. 16 Briefly, the transfected cells in serum-free medium were seeded at 1.0 × 105 per well into the upper compartment of Transwell chambers. RPMI-1640 medium containing 10% FCS was placed into the lower compartment. Twenty-four hours later, the penetrated cells through the pores of inserts were fixed with 4% paraformaldehyde and stained with 0.2% crystal violet. Images were photographed, and the number of migrated or invaded cells was counted in three random fields under a light microscope (100 × magnification, Nikon, Tokyo, Japan).
Western blot
Total protein was prepared from tissues and cells as previously described. 17 Protein samples (30–50 μg) were resolved on an 8% sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) under a denaturing condition and then electroblotted onto a Hybond enhanced chemiluminescence (ECL) nitrocellulose membrane (Amersham Biosciences, Little Chalfont, UK). After being blocked with 5% nonfat milk in TBS-Tween, the membranes were probed with primary antibodies, followed by incubation with a secondary antibody. The signals were detected using an ECL system (Pierce, Thermo Fisher Scientific, Cramlington, UK). Primary antibodies against vimentin (#5741), N-cadherin (#13116), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH, #5174) were purchased from Cell Signaling Technology (Danvers, MA). Primary antibodies against E-cadherin (ab40772) and EGFR (ab52894) were obtained from Abcam (Cambridge, UK). The horseradish peroxidase-conjugated goat anti-rabbit IgG antibody (ab6721; Abcam) was used as the secondary antibody.
Bioinformatics and dual-luciferase reporter assay
The starBase v.3 software was used to predict the miRNAs that potentially bind to circ_0081143 and the direct targets of miR-497-5p available at
Statistical analysis
Data were analyzed by a two-tailed Student's t-test, Mann–Whitney U test, or one-way analysis of variance followed by Tukey's post hoc test. Correlations among circ_0081143, miR-497-5p, and EGFR expression in GC tissues were tested using the Spearman test. The results are reported as mean ± standard deviation. P values at 0.05 or smaller were considered significant.
Results
Circ_0081143 was highly expressed in GC tissues and cells
First, the authors assessed the expression of circ_0081143 in GC tissues and cell lines. As shown by qRT-PCR, the circ_0081143 level was significantly increased in GC tissues compared with the nontumor tissues (Fig. 1A). In line with GC tissues, circ_0081143 expression was higher in GC cells than that of controls (Fig. 1B).
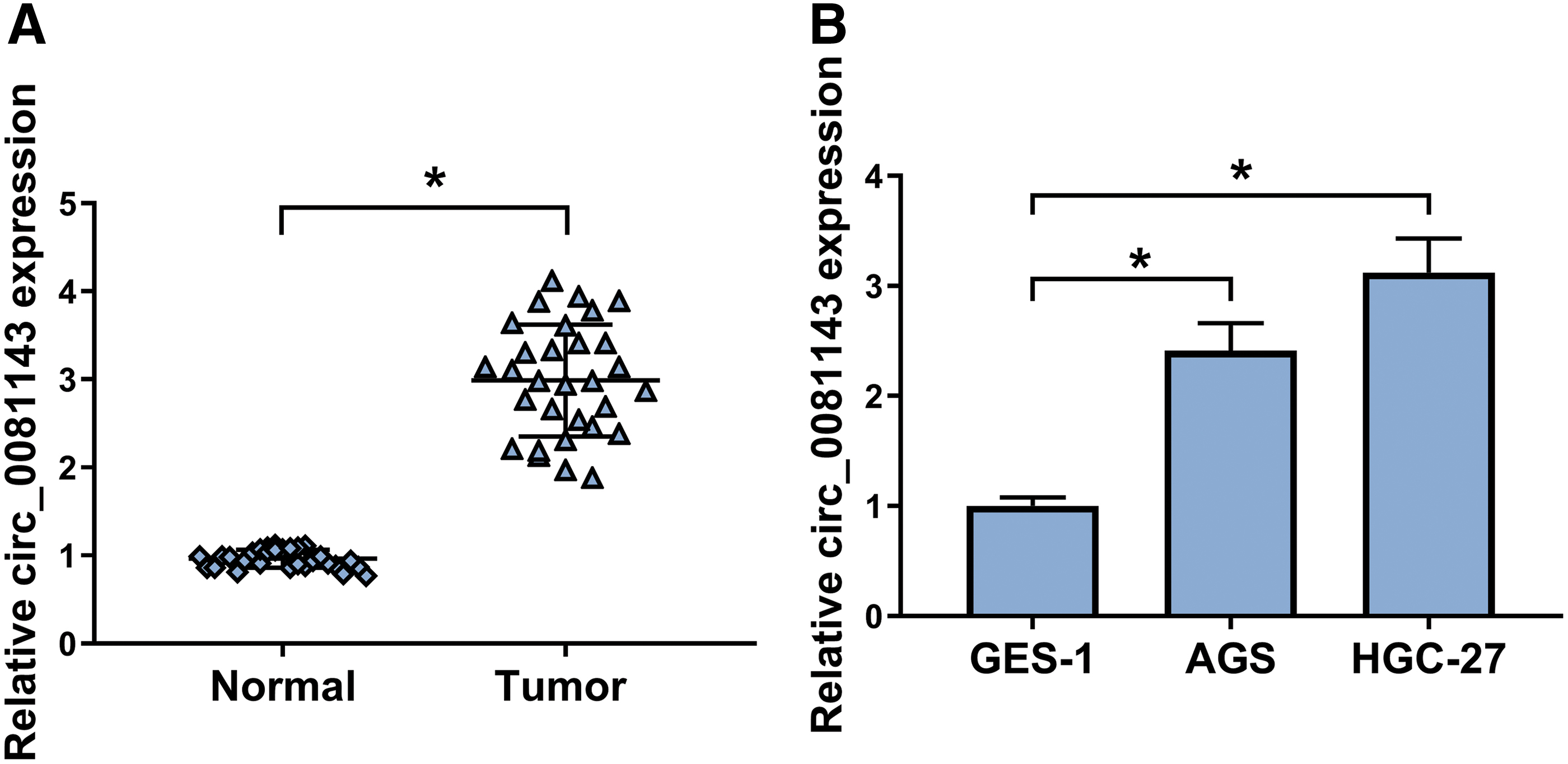
Circ_0081143 expression was elevated in GC tissues and cells. The expression of circ_0081143 by qRT-PCR in 30 pairs of GC tissues and adjacent nontumor gastric tissues
Circ_0081143 depletion relieved hypoxia-induced migration, invasion, and EMT in GC cells
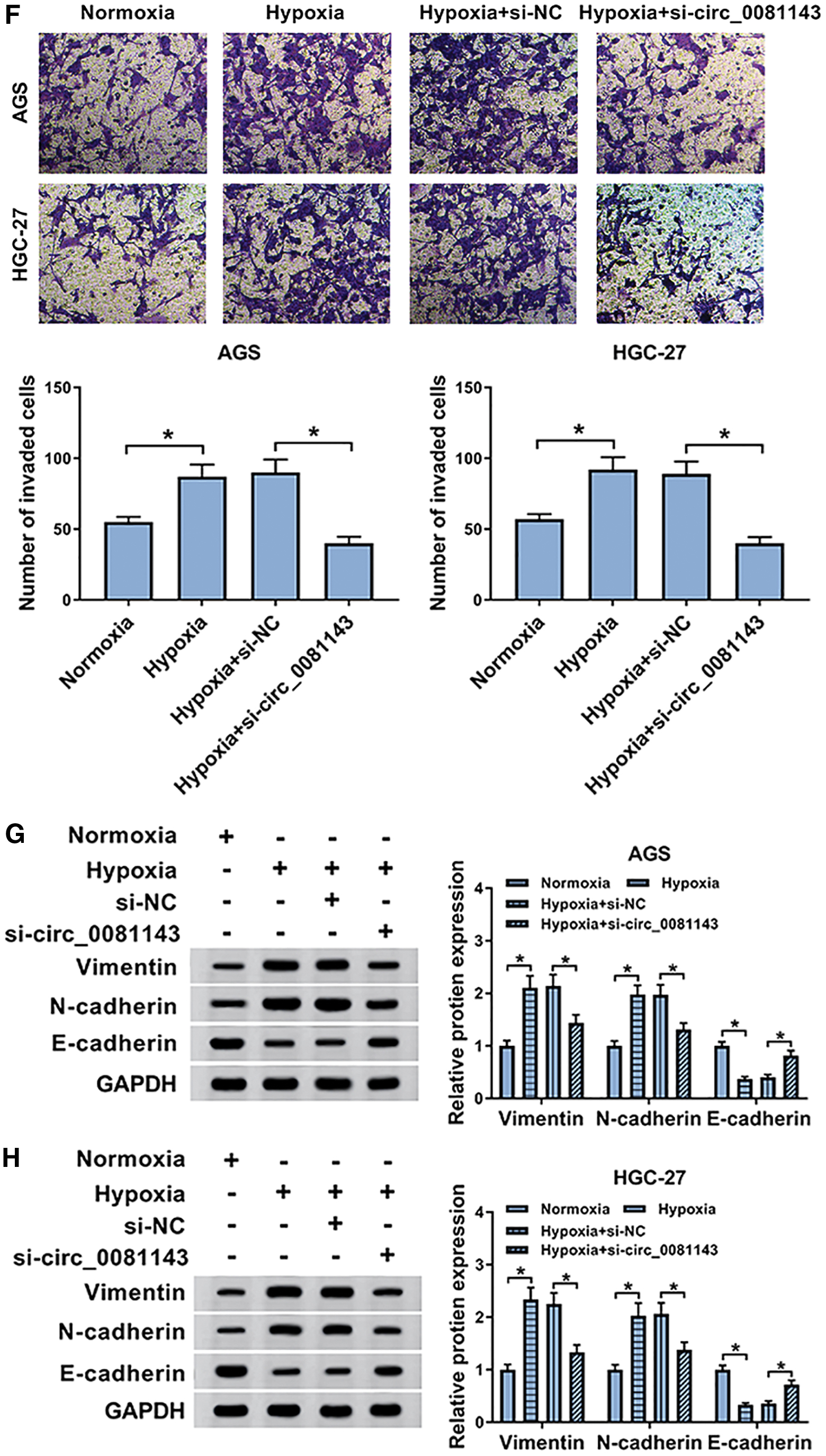
The authors then examined the influence of hypoxia on circ_0081143 expression in GC cells. The results of qRT-PCR revealed that hypoxia exposure resulted in increased levels of circ_0081143 in both AGS and HGC-27 cells (Fig. 2A–D). Transwell assays showed that compared with the negative control, cell migration and invasion abilities were remarkably accelerated by hypoxia exposure (Fig. 2E, F). Moreover, the data of Western blot demonstrated that hypoxia led to a significant increase in vimentin and N-cadherin levels, as well as a striking reduction in E-cadherin expression in the two GC cells (Fig. 2G, H), suggesting the promotion of hypoxia on cell EMT. To determine the impact of circ_0081143 on GC progression under hypoxia, loss-of-function experiments were carried out using si-circ_0081143. As demonstrated by qRT-PCR, transient transfection of si-circ_0081143 prominently declined the expression of circ_0081143 (about 59% downregulation in AGS cells and 53% in HGC-27 cells) in hypoxia-treated GC cells (Fig. 2C, D). Subsequent experiments' data revealed that hypoxia-induced promigration (Fig. 2E), proinvasion (Fig. 2F), and pro-EMT (Fig. 2G, H) effects were markedly abolished by circ_0081143 knockdown in the two cells.

Circ_0081143 depletion hampered hypoxia-induced migration, invasion, and EMT in GC cells.
Circ_0081143 acted as an miR-497-5p sponge in GC cells
To further determine the mechanism by which circ_0081143 regulated hypoxia-induced GC cell malignant behaviors, the authors performed a detailed analysis for its targeted miRNAs. In silico prediction by starBase v.3 database of circ_0081143 miRNA targets revealed that circ_0081143 harbored a putative complementary sequence for miR-497-5p (Fig. 3A). To validate this, the authors cloned the partial sequence of circ_0081143 containing the miR-497-5p-binding sites into a luciferase plasmid and mutated the target sites. Cotransfection of miR-497-5p mimic and circ_0081143 wild-type reporter into GC cells resulted in a striking decrease in luciferase activity (60% reduction in AGS cells and 64% in HGC-27 cells) (Fig. 3B, C). However, upon transfection of the mutant reporter, the downregulation of miR-497-5p in luciferase activity was completely abolished (Fig. 3B, C), suggesting the validity of the binding sites for interaction. The data of qRT-PCR demonstrated that miR-497-5p was downregulated in GC tissues and cells (Fig. 3D, E). Moreover, hypoxia exposure triggered a significant reduction in miR-497-5p expression in both AGS and HGC-27 cells (Fig. 3F, G). Importantly, miR-497-5p expression was prominently elevated by circ_0081143 depletion in the two GC cells under hypoxia (Fig. 3H, I), indicating that the miR-497-5p-binding sites were functional.
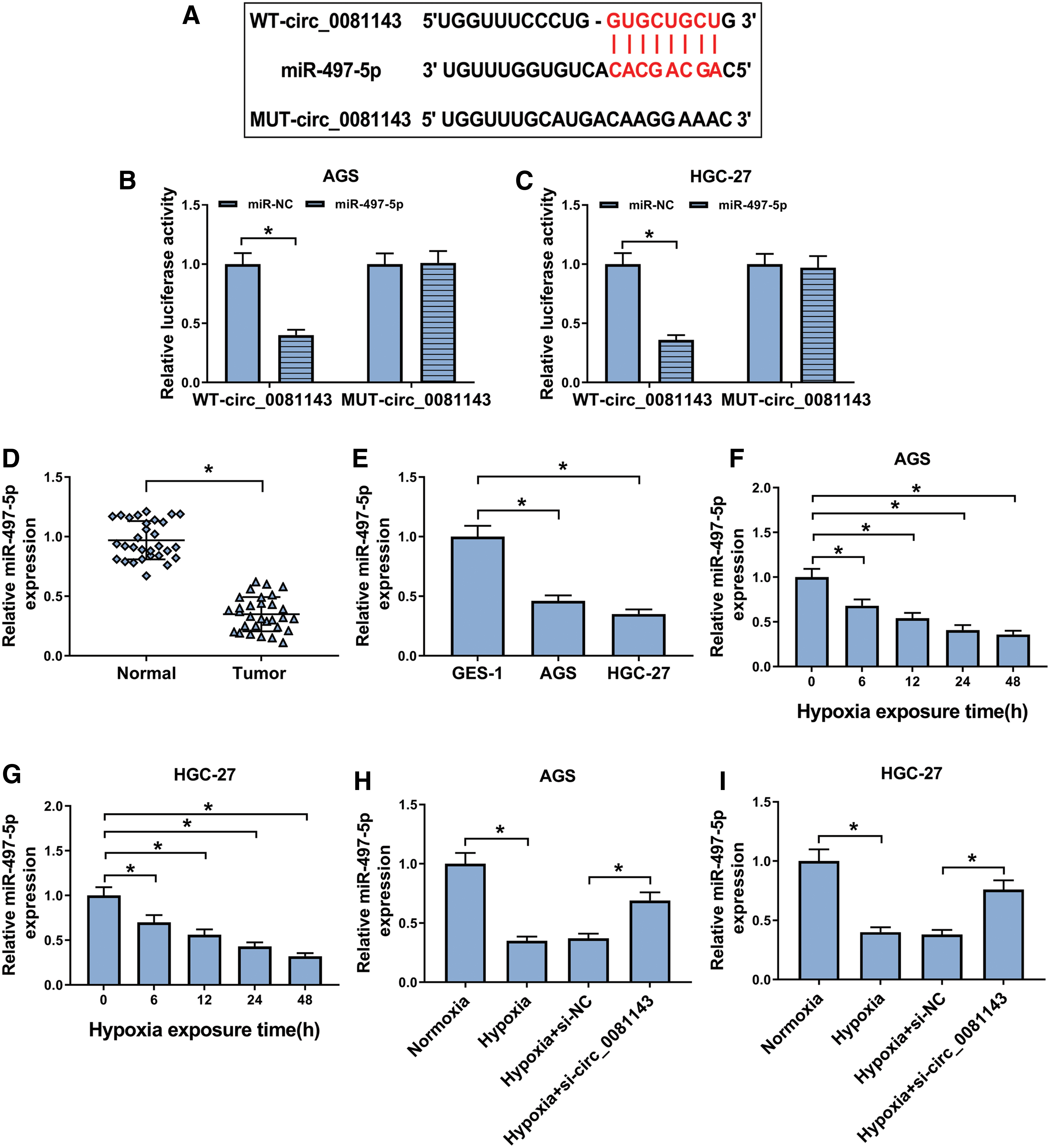
Circ_0081143 directly interacted with miR-497-5p through binding to miR-497-5p.
MiR-497-5p mediated the alleviative effects of circ_0081143 knockdown on GC cell migration, invasion, and EMT under hypoxia
To observe whether circ_0081143 depletion hampered GC cell malignant behaviors under hypoxia by miR-497-5p, si-circ_0081143 and anti-miR-497-5p were cotransfected into AGS and HGC-27 cells before hypoxia. qRT-PCR results showed that compared with the negative control, si-circ_0081143-mediated miR-497-5p augment was significantly reversed by anti-miR-497-5p cotransfection in the two cells under hypoxia (Fig. 4A, B). Subsequently, the analyses of Transwell and Western blot showed that si-circ_0081143-mediated antimigration (Fig. 4C, D), anti-invasion (Fig. 4E, F), and anti-EMT effects (Fig. 4G, H) were remarkably abrogated by the restored expression of miR-497-5p under hypoxia.
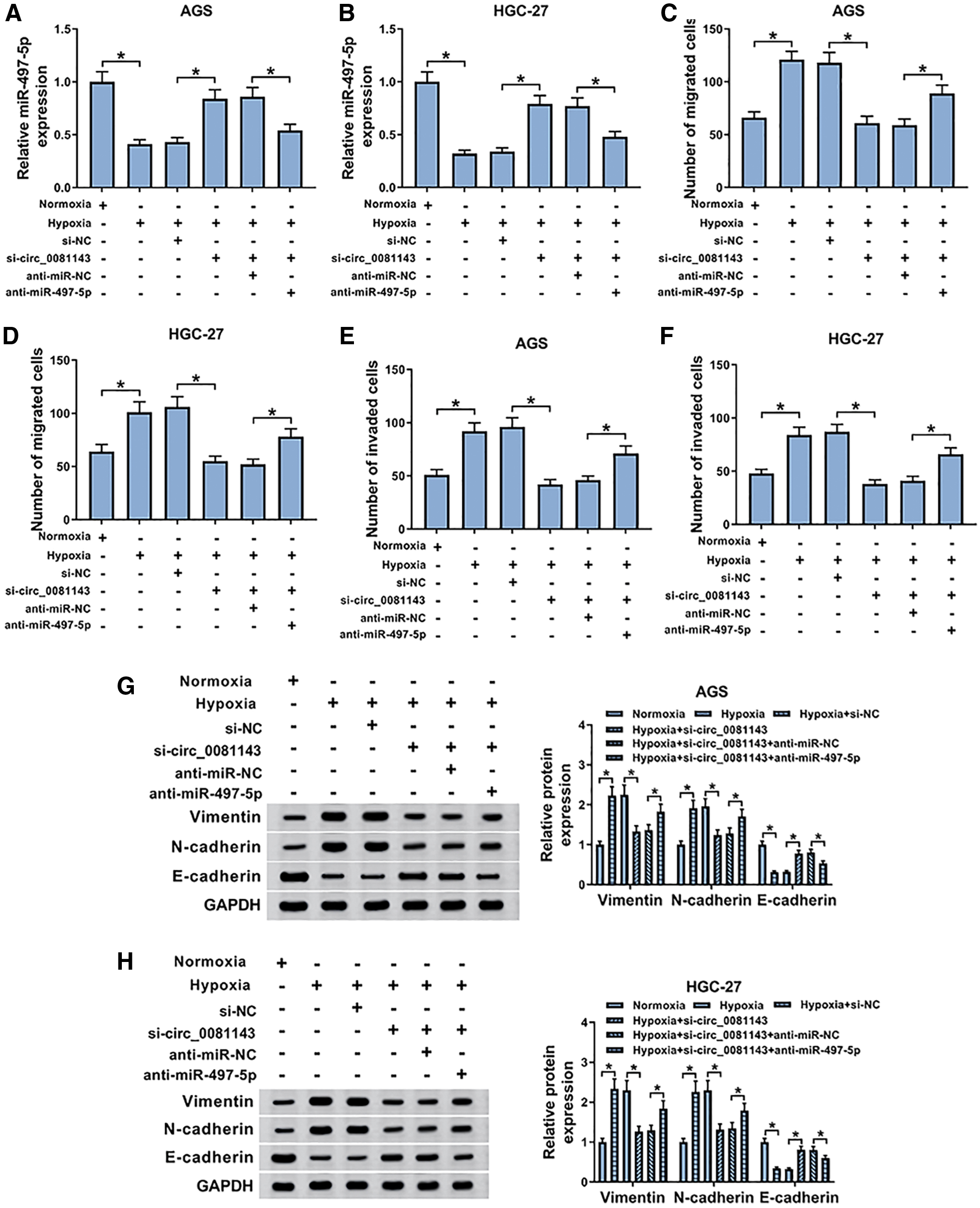
The alleviative effects of circ_0081143 knockdown on GC cell migration, invasion, and EMT under hypoxia were abolished by miR-497-5p expression restoration. The expression of miR-497-5p by qRT-PCR
MiR-497-5p directly interacted with the 3′-UTR of EGFR
To further understand the role of miR-497-5p in GC, the authors next used starBase v.3 software to help identify its molecular targets. Of interest, the data revealed a putative target sequence for miR-497-5p in EGFR 3′-UTR (Fig. 5A). To confirm whether EGFR was a target of miR-497-5p, they conducted dual-luciferase reporter assays. With EGFR 3′-UTR, luciferase reporter and miR-497-5p overexpression caused a striking downregulation in luciferase activity (59% in AGS cells and 64% in HGC-27 cells) (Fig. 5B, C). When the target sequence was mutated, little change was observed in luciferase activity in the presence of miR-497-5p mimic (Fig. 5B, C). The data of qRT-PCR and Western blot demonstrated that the EGFR mRNA and protein levels were significantly upregulated in GC tissues and cells (Fig. 5D–G). Moreover, hypoxia exposure resulted in increased EGFR expression in a time-dependent manner in both AGS and HGC-27 cells (Fig. 5H–K).
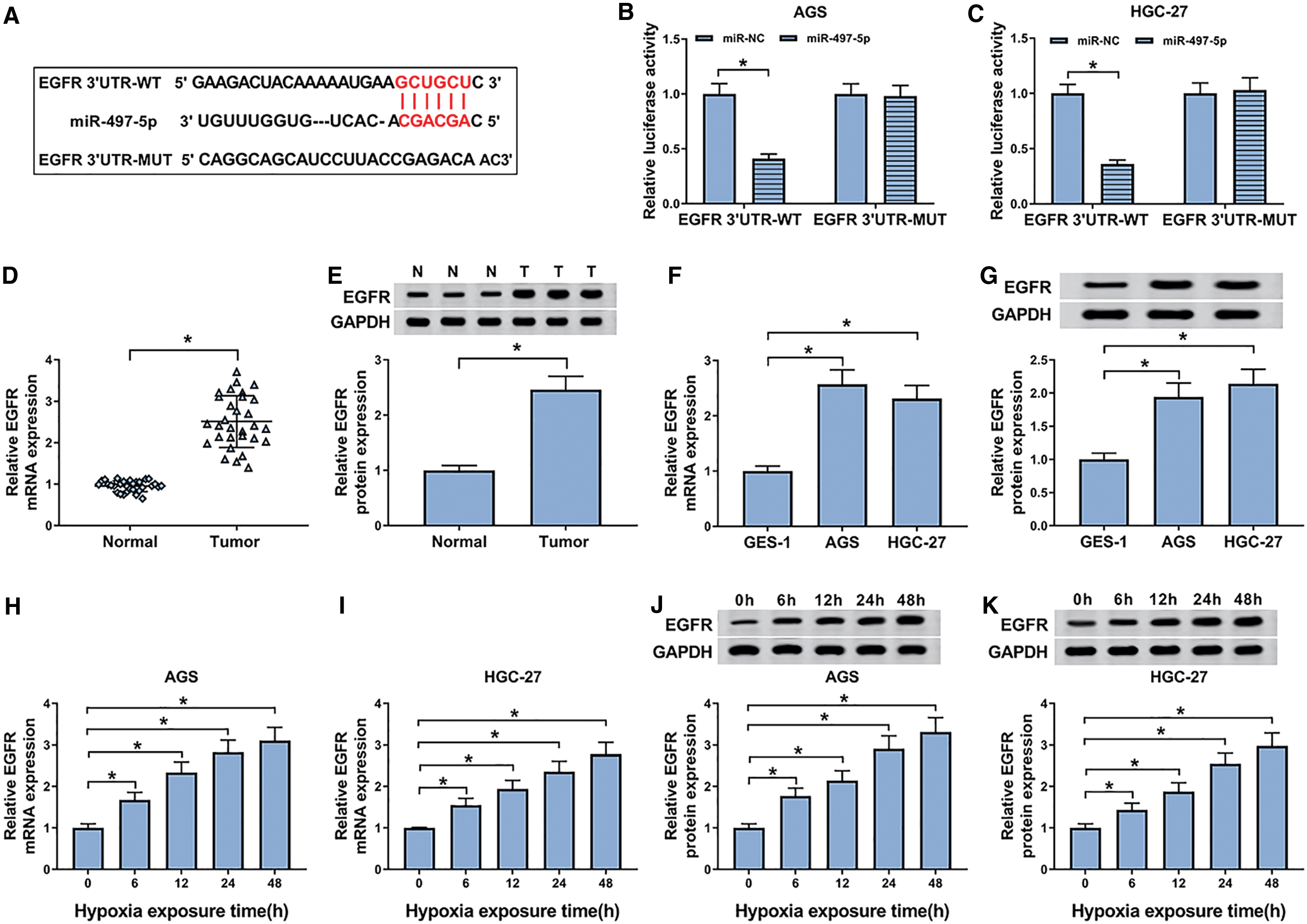
MiR-497-5p interacted directly with the 3′-UTR of EGFR.
MiR-497-5p overexpression mitigated hypoxia-induced GC cell migration, invasion, and EMT by downregulating EGFR
To assess whether miR-497-5p regulated EGFR expression in GC cells under hypoxia, AGS and HGC-27 cells were introduced with miR-497-5p mimic before hypoxia exposure. The analyses of qRT-PCR and Western blot revealed that compared with the negative group, EGFR expression was prominently reduced by miR-497-5p overexpression at both mRNA and protein levels in the two cells (Fig. 6A–D). Subsequent experiments data showed that the elevated expression of miR-497-5p led to the significant suppression in cell migration (Fig. 6E, F), invasion (Fig. 6G, H), and EMT (Fig. 6I, J) under hypoxia. To provide further mechanistic insight into the link between miR-497-5p and EGFR on GC cell malignant behaviors under hypoxia, miR-497-5p mimic and EGFR overexpression plasmid were cotransfected into the two cells before hypoxia exposure. These results demonstrated that compared with the negative control, the repression of miR-497-5p upregulation in EGFR expression was strongly reversed by the cotransfection of EGFR overexpression plasmid (Fig. 6A–D). More importantly, the mitigatory effects of miR-497-5p overexpression on GC cell migration (Fig. 6E, F), invasion (Fig. 6G, H), and EMT (Fig. 6I, J) under hypoxia were dramatically abrogated by EGFR expression restoration.
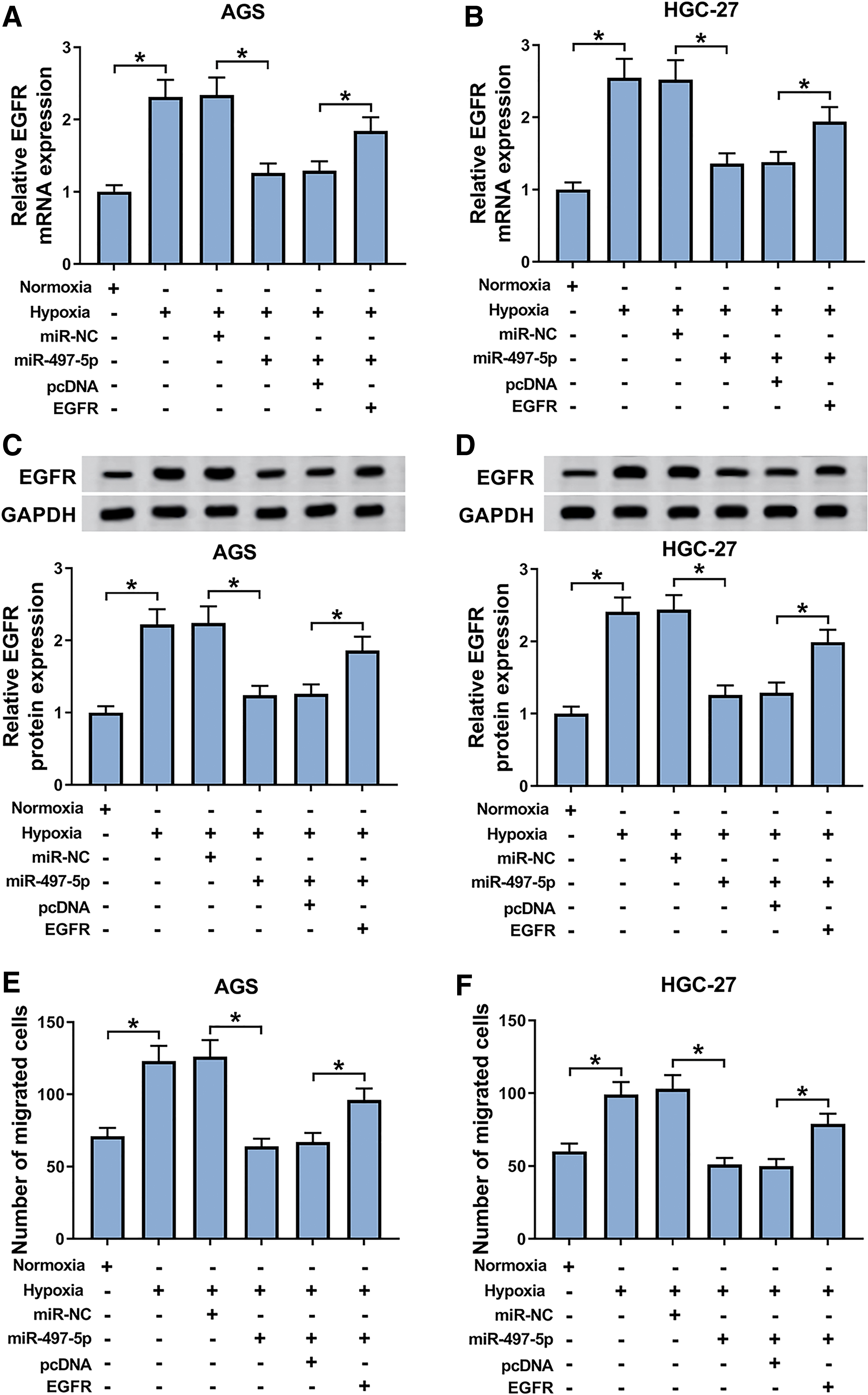
The mitigatory effects of miR-497-5p overexpression on GC cell migration, invasion, and EMT under hypoxia were mediated by EGFR. EGFR mRNA expression by qRT-PCR
Circ_0081143 regulated EGFR expression through sponging miR-497-5p
In GC tissues, the miR-497-5p level was inversely correlated with circ_0081143 and EGFR expression, and EGFR expression was positively correlated with circ_0081143 level (Fig. 7A–C). CircRNAs protect against gene expression inhibition through sequestering miRNAs via binding to miRNAs. 18 Hence, the authors explored whether circ_0081143 modulated EGFR expression in GC cells under hypoxia. As expected, EGFR expression was prominently repressed by circ_0081143 depletion at both mRNA and protein levels in the two GC cells under hypoxia (Fig. 7D–G). However, this effect was significantly abolished by the cotransfection of anti-miR-497-5p (Fig. 7D–G).
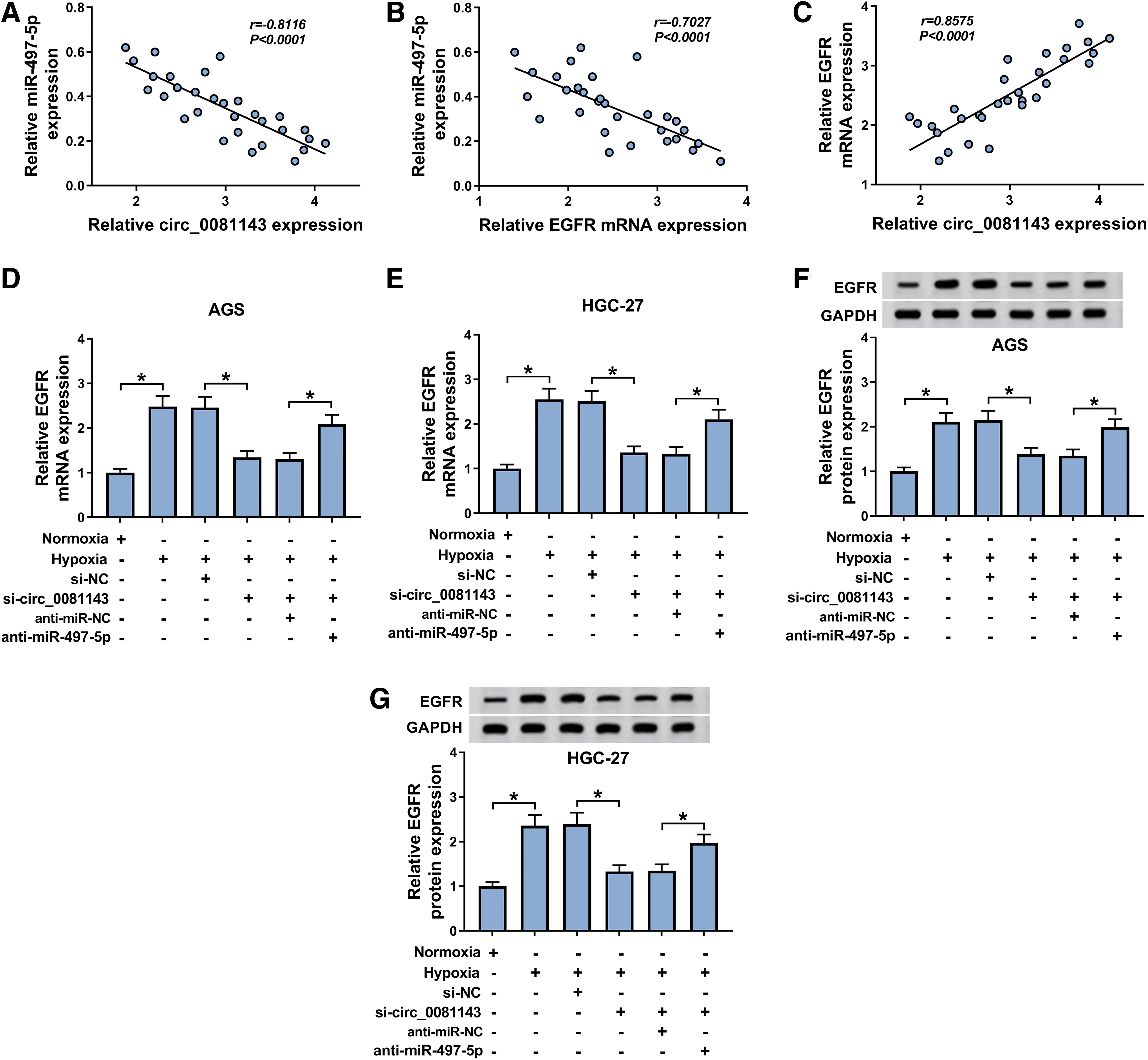
Circ_0081143 modulated EGFR expression via acting as a miR-497-5p sponge in GC cells under hypoxia.
Discussion
CircRNAs are being found as novel regulators in the cancer paradigm, demonstrating potential oncogenic and tumor-suppressive roles. 19 These new transcripts are frequently dysregulated in numerous human cancers; however, the biological roles and underlying mechanisms of the vast majority remain unclear. Hypoxia is a feature of most tumors, and circRNAs are associated with the regulation of hypoxia. 20 Among these GC-related circRNAs, circ_0081143 was of interest in this work owing to its oncogenic role in GC chemoresistance. 9 In the present study, the authors were first to identify that circ_0081143 depletion weakened hypoxia-induced migration, invasion, and EMT of GC cells through the miR-497-5p/EGFR axis.
In the present work, they first assessed the expression of circ_0081143 in GC tissues and cells, and these data validated a striking upregulation of circ_0081143 in GC, consistent with a recent report. 9 Previous research had demonstrated that several circRNAs, such as circRNA differentially expressed in normal cells and neoplasia domain containing 4C (circDENND4C) and zinc finger protein 292 (circZNF292), were induced by hypoxia. 16,20 Similarly, these data for the first time uncovered that hypoxia resulted in increased circ_0081143 expression in GC cells. Hypoxia was also underscored to drive tumor cell metastatic potential through the enhancement of cell migration, invasion, and EMT. 21,22 In the present study, the data supported that the migration, invasion, and EMT of GC cells were remarkably promoted by hypoxia. Importantly, the authors were first to confirm that circ_0081143 knockdown mitigated hypoxia-induced GC cell malignant behaviors. In short, circ_0081143 knockdown performed a tumor-suppressive role in GC under hypoxia, in line with a recent study. 9
It is widely accepted that circRNAs modulate gene expression through acting as ceRNAs of miRNAs via sequestering miRNAs. 18 To determine whether circ_0081143 could serve as a molecular sponge in GC cells, they performed a detailed analysis for its targeted miRNAs. Subsequently, the authors verified that circ_0081143 sequestered miR-497-5p by binding to miR-497-5p. MiR-497-5p has been demonstrated as a tumor suppressor in a series of human cancers, such as nonsmall-cell lung cancer, hepatocellular carcinoma, and melanoma. 23 –25 Moreover, miR-497-5p was involved in the ceRNA network in colorectal cancer and diffused large B cell lymphoma. 26,27 These data also demonstrated that miR-497-5p was downregulated in GC tissues and cells, in accordance with previous results. 12 In addition, under hypoxia, miR-497-5p expression was reduced in GC cells. Using gain-of-function experiments, the authors found that miR-497-5p overexpression relieved hypoxia-induced GC cell migration, invasion, and EMT. Furthermore, for the first time, they uncovered that miR-497-5p mediated the alleviative effects of circ_0081143 depletion on GC cell malignant behaviors under hypoxia.
MiRNAs exert biological function by regulating their target gene expression. the authors then predicted the targets of miR-497-5p using starBase v.3 software. Among these candidates, EGFR was particularly fascinating in the present research because its expression was an independent indicator for poor prognosis of GC patients. 28,29 A large amount of evidence showed that EGFR acted as a tumor promoter in GC. 30,31 Zhang et al. highlighted that EGFR enhanced live metastasis of GC through regulating the liver microenvironment. 32 These data indicated a striking augmentation of EGFR expression in GC tissues and cells, in line with earlier documents. 33,34 The authors were first to confirm that EGFR was directly targeted and inhibited by miR-497-5p. Previous studies had manifested that EGFR was involved in the regulation of hypoxia, and hypoxia was associated with antitumor effects of EGFR inhibitors. 35,36 In the present work, the data also validated that hypoxia induced EGFR expression in GC cells. Moreover, for the first time, they substantiated that the mitigatory effects of miR-497-5p overexpression on GC cell migration, invasion, and EMT under hypoxia were mediated by EGFR. More importantly, this study illuminated that circ_0081143 regulated EGFR expression through acting as a sponge of miR-497-5p in hypoxia-induced GC cells.
In conclusion, the present work suggested that the knockdown of circ_0081143 mitigated hypoxia-induced migration, invasion, and EMT in GC cells at least partly through sponging miR-497-5p and modulating EGFR expression. Targeting circ_0081143 might be a promising therapeutic strategy for GC management.
Availability of Data and Materials
The analyzed data sets generated during the present study are available from the corresponding author on reasonable request.
Footnotes
Authors' Contributions
Conceptualization and methodology: H.Z. and J.L. Formal analysis and data curation: J.L. and H.W. Validation and investigation: J.T. and H.W. Writing—original draft preparation and writing—review and editing: J.T., H.Z., and J.L. Approval of final article: all authors.
Ethics Approval and Consent to Participate
The present study was approved by the Ethics Review Committee of Xiangyang No. 1 People's Hospital (Affiliated Hospital of Hubei University of Medicine).
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.