
Abstract
Aim:
Breast cancer is one of the leading causes of cancer-associated death. This work aimed to explore the expression, function, and mechanism of circ_0000526 on the progression of breast cancer.
Materials and Methods:
The expressions of circ_0000526, miR-492, and suppressor of cytokine signaling 2 (SOCS2) in breast cancer samples were measured using quantitative real-time polymerase chain reaction. The correlation between pathological indexes of patients and the expression of circ_0000526 was also analyzed. Human breast cancer cell lines MCF-7 and MDA-MB-231 were applied to investigate the associated mechanism. Cell counting kit-8 assays were used to assess the effect of circ_0000526 on proliferation, and together with the wound healing assay, transwell assay was conducted to detect the effects of circ_0000526 on migration and invasion. Furthermore, luciferase reporter assay was used to verify the targeting relationship between miR-492 and circ_0000526. In addition, Western blot was performed to detect the regulatory effects of circ_0000526 on SOCS2.
Results:
Circ_0000526 expression in breast cancer samples was downregulated, which was correlated with unfavorable pathological indexes. It significantly inhibited the proliferation and metastasis of breast cancer cells, and facilitated apoptosis. The overexpression of circ_0000526 remarkably inhibited the expression of miR-492 by sponging it, and in turn promoted the expression of SOCS2.
Conclusions:
Circ_0000526 served as a tumor suppressor, which could function as the sponge of miR-492, an oncogenic microRNA, in breast cancer, and enhance the expression of tumor suppressor SOCS2 indirectly.
Introduction
Breast cancer is one of the most common malignant tumors in women, which seriously threatens women's life and health. 1 Even though the mortality rate of breast cancer might be controlled by population screening, the incidence of breast cancer is still at a high level worldwide. 2 With the development of biological research on cancer-associated molecule, molecular target therapy for breast cancer has been widely applied in the treatments of breast cancer, which has achieved remarkable results, while the tumor tissues derived from some patients are unresponsive to the current target therapy, and some other patients still suffer drug resistance after a period of treatment. 3 As a consequence, it is of importance to clarify the mechanism of breast cancer progression for the prevention and treatment of breast cancer.
Circular RNA (circRNA) is a class of noncoding RNA with stable circular structure. It is involved in the transcription and post-transcriptional regulation of genes. 4 In recent years, accumulating studies indicate that circRNAs are related to the occurrence and progression of tumors, providing a new direction for the diagnosis and treatment of tumor. 5 In breast cancer, multiple circRNAs have been identified as regulator in cancer progression, including circRNA_100876, circKDM4C, and circPLK1. 6 –8 Many circRNAs exert function as a competitive endogenous RNA (also called molecular sponge) by inhibiting the expression of downstream microRNA (miRNA). 6 –8 Compared with circRNA, the functions of miRNA in human disease have been more widely and deeply investigated. 9,10 It is reported that miR-492 can promote the progression of multiple cancers as an oncomiR, including prostate cancer, liver cancer, and breast cancer. 11 –14 Its downstream genes include PTEN, SOX7, CD44, and suppressor of cytokine signaling 2 (SOCS2). 11 –14 Interestingly, bioinformatic analysis suggested that miR-492 could be specifically regulated by circ_0000526.
This work aimed to explore the expression and function of circ_0000526 in breast cancer. The authors also tried to validate the binding relationship between miR-492 and circ_0000526. The associated results demonstrated that circ_0000526 was lowly expressed in breast cancer tissues, and it repressed the expression of miR-492, but promoted the expression of tumor suppressor SOCS2. This work depicted a novel mechanism that could contribute to the progression of breast cancer, namely the circ_0000526/miR-492/SOCS2 axis, providing target for the diagnosis and treatment of breast cancer.
Materials and Methods
Tissue samples
The breast cancer samples derived from forty patients who were subjected to surgery in Xianning Central Hospital from April 2017 to April 2018 were enrolled. In the control group, the specimens were obtained from the adjacent tissues of the same patient (at least 3 cm away from the surgical margin), and no cancer cells were found by pathological examination. All specimens were removed during the surgery and then immediately stored in liquid nitrogen at −196°C for subsequent RNA extraction. The collection and use of the samples were approved by the Research Ethics Committee of Xianning Central Hospital, and informed consent from all patients was obtained.
Cell culture and cell transfection
Human normal breast epithelial cell line (MCF-10A) and breast cancer cell line (MDA-MB-231, MCF-7, ZR751) were purchased from the Cell Center of Chinese Academy of Sciences (Shanghai, China). The above cells were maintained in RPMI 1640 Medium (Thermo Fisher Scientific, Waltham, MA) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific) and 1% penicillin/streptomycin (Invitrogen, Carlsbad, CA) at 37°C in 5% carbon dioxide (CO2). pcDNA empty vector serving as negative control (NC), pcDNA-circ_0000526 (circ_0000526), short hairpin RNA (shRNA) negative control, shRNAs interfering with circ_0000526, miRNA negative control, miR-492 mimics, and miR-492 inhibitors were designed and provided by GenePharma Co., Ltd. (Shanghai, China). The sequence of shRNA targeting circ_0000526: ATCTCCTGGCTTCAGTCCTTA. MDA-MB-231 and MCF-7 cells were inoculated on a 24-well cell culture plate at a density of 3 × 105 cells per well. The cells were incubated at 37°C in 5% CO2 for 24 h and then subjected to transfection, which was performed using Lipofectamine 3000 (Invitrogen and Thermo Fisher Science) in compliance with the supplier's instructions. Subsequently, the transfection efficiency was detected by quantitative real-time polymerase chain reaction (qRT-PCR) after 24 h of the transfection.
Quantitative RT-PCR
Total RNA in tissues and breast cancer cells was extracted using the TRIzol reagent (Invitrogen, Shanghai, China). Complementary DNA was synthesized using the PrimeScript Reverse Transcription Kit (TaKaRa, Dalian, China). Following that, RT-PCR was conducted on ABI7500 Real-Time PCR System (Applied Biosystems, San Francisco, CA) using SYBR premix EX TAQ II (TaKaRa) in accordance with the manufacturer's instructions. Besides, GAPDH served as the internal reference for circ_0000526 and SOCS2. And U6 was used as the internal reference of miR-492. The results of gene expression were analyzed using the 2(−ΔΔCt) method. Specific primer sequences are demonstrated as follows: circ_0000526: forward: 5′-GGGAAGACGTCAGCTCT-3′ and reverse: 5′-AGCTATGCCAGGGTCTT-3′; miR-492: forward: 5′-CGGGATATTATCGAGGTATTC-3′ and reverse: 5′-AACTAACAAACCCTACCG-3′; SOCS2: forward: 5′-ACTCTTGCCAAGTCGTCTCGTCG-3′ and reverse: 5′-ATCCTGGAGGACGGATGACA-3′; GAPDH forward: 5′-ATGCCTCCTGCACCACCAACTGCTT-3′ and reverse: 5′-TGGCAGTGATGCATGGACTGTGGT-3′; and U6: forward: 5′-GCTTCGCAGCATATACTAAT-3′ and reverse: 5′-CGCTTCACGAATTTGGTGTCAT-3′.
Cell proliferation assay in vitro
Cells, 2 × 103, in each group were inoculated into a 96-well plate. After being cultured for 24 h, 10 μL cell counting kit-8 (CCK-8) solution (Beyotime, Beijing, China) was added to each well. Then the culture was maintained for 1 more hour. After that, the absorbance value at 450 nm wavelength in each well was measured using a microplate reader. The same method was applied to detect the absorbance values in each well at 48, 72, and 96 h after the inoculation. Ultimately, the curve of proliferation was plotted based on the absorbance values.
Wound healing assay
MDA-MB-231 and MCF-7 cells at logarithmic phase were inoculated into 24-well plates at a density of 2.5 × 105 cells per well, and maintained until the cell monolayer covered the bottom of each well. Subsequently, the medium was removed. After each well being rinsed with phosphate-buffered saline (PBS) twice, a scratch was drawn evenly using the head of a sterile 200 μL pipette tip. After that, the culture plate was rinsed with PBS again, and serum-free medium was added to each well. Following that, the scratches were observed and photographed under a microscope. After another 24 h of culture, the scratches were observed and photographed again.
Transwell assay
Cells were trypsinized, harvested, and resuspended in serum-free medium at a density of 1 × 105 cells/mL. Then 200 μL of the above medium containing cells were added into the upper chamber of the Transwell system with 8 μm pore membrane (BD Biosciences, San Jose, CA) coated with Matrigel, and 600 μL of the above medium supplemented with 10% FBS was added into the bottom chamber. After being cultured for 24 h, cells on the upper surface of the membrane were wiped off using a cotton swab. Following that, the cells passing through the membrane were fixed with 4% polyformaldehyde and stained with crystal violet. After the membrane was rinsed and dried, five randomly selected fields under the microscope on the membrane were used to calculate the number of invasive cells, and the mean value was adopted to evaluate the invasiveness of the cells.
Dual-luciferase reporter gene assay
All luciferase reporter assay-associated vectors circ_0000526-wt, circ_0000526-mut, SOCS2-wt, and SOCS2-mut were constructed by Promega (Madison, WI). Circ_0000526-wt or circ_0000526-mut as well as SOCS2-wt or SOCS2-mut vectors were cotransfected into MDA-MB-231 and MCF-7 cells with miR-492 mimics or control microRNA, respectively, using Lipofectamine 3000. After 48 h of transfection, luciferase activity was determined in compliance with the manufacturer's instructions.
Western blot
RIPA lysate (containing 1% phenylmethanesulfonyl-fluoride [PMSF]) was applied to the extraction of the total protein from the cells. After being purified and determined, 5 μg of the total protein was subjected to electrophoresis using 12% polyacrylamide gel at 100 V for 2 h. Subsequently, the protein was electrotransferred to polyvinylidene fluoride membrane (Millipore, Bedford, MA). After being blocked using 5% skimmed milk at room temperature for 1 h, the membrane was rinsed three times with tris buffered saline tween (TBST) for 10 min each time, and then incubated overnight with the primary anti-SOCS2 antibody (ab3692, rate of dilution 1:1000; Abcam) at 4°C. Following that, after being rinsed using TBST three times (15 min each time), the membrane was incubated with a secondary antibody at room temperature for 2 h. Subsequently, the membrane was rinsed for three times using TBST. Ultimately, the enhanced chemiluminescence detection kit (Millipore) was used to visualize the bands.
Statistical analysis
SPSS 17.0 statistical software (SPSS, Inc., Chicago, IL) was applied to statistical analysis. The data are expressed as mean ± standard deviation (x ± s). Student's t-test was used for the comparison between every two groups. In addition, to be notable, the counting in wound healing assay was expressed by a fourfold table, and the difference between every two groups was analyzed using the χ 2 test. p-Values <0.05 were considered to be statistically significant.
Results
Expressions of circ_0000526, miR-492, and SOCS2 in breast cancer
To investigate the role of circ_0000526, miR-492, and SOCS2 in breast cancer, first of all, the authors detected the expressions of circ_0000526, miR-492, and SOCS2 in breast cancer tissues. RT-PCR results demonstrated that the expression of circ_0000526 in breast cancer tissue was significantly downregulated, while that of miR-492 was upregulated (Fig. 1A, B). In addition, RT-PCR manifested that the expression of SOCS2 messenger RNA (mRNA) was significantly downregulated in breast cancer (Fig. 1C). Then the χ 2 analysis was used to evaluate the correlation between circ_0000526 and clinicopathological features of breast cancer patients. Also, the data indicated that low expression of circ_0000526 was significantly correlated with the increased incidence of lymph node metastasis (Table 1). The above results implied that circ_0000526 and SOCS2 could probably inhibit the progression of breast cancer, while miR-492 served as an oncogenic miRNA. 14
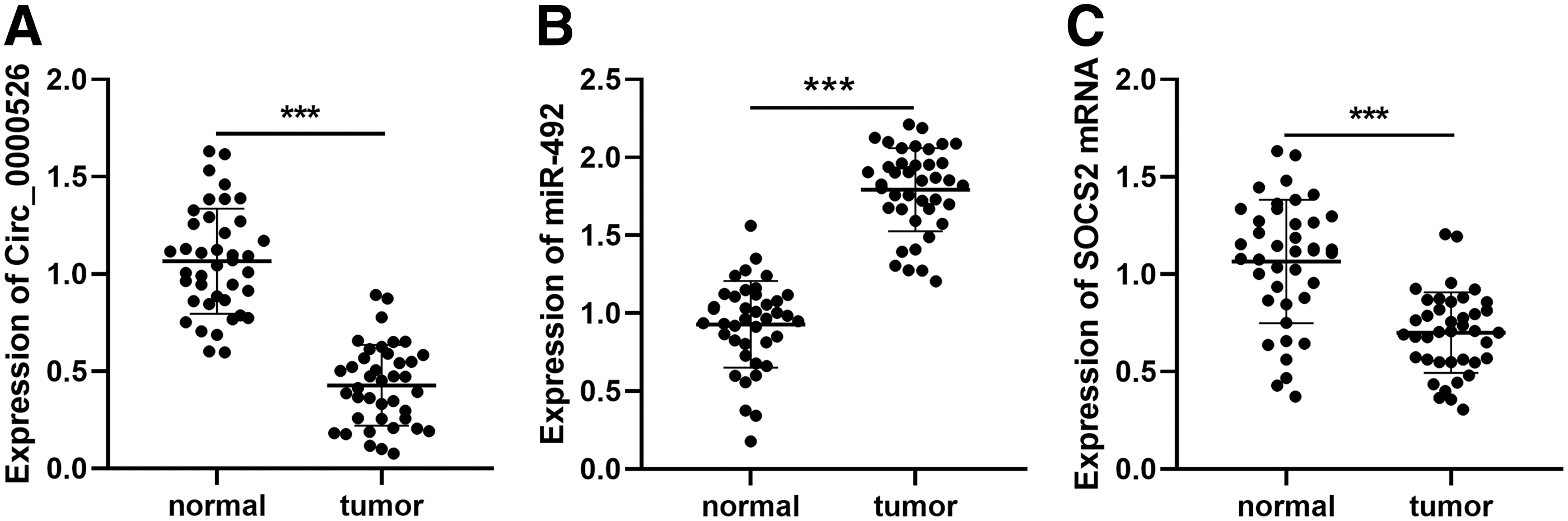
Expressions of circ_0000526, miR-492, and SOCS2 in breast cancer.
Association Between Circ_0000526 Expression and Clinicopathological Characteristics
Presents the statistic difference < 0.01.
ER, estrogen receptor; HER-2, human epidermal growth factor receptor-2; PR, progesterone receptor.
Effects of circ_0000526 on proliferation and metastasis of breast cancer cells
To clarify the effects of circ_0000526 on proliferation and metastasis, the authors detected the expression of circ_0000526 in breast cancer cell lines. RT-PCR results demonstrated that the expression of circ_0000526 in breast cancer cell lines, including MDA-MB-231, MCF-7, and ZR751, was significantly lower than that in the normal breast epithelial cell line MCF-10A (Fig. 2A). After that, they successfully constructed circ_0000526 overexpression and knockdown cell lines using breast cancer cell lines MDA-MB-231 and MCF-7, respectively (Fig. 2B, C). CCK-8 results suggested that the proliferation of breast cancer cells was inhibited after the overexpression of circ_0000526, but promoted after the knockdown of circ_0000526 (Fig. 2D, E). The results of wound healing assay and Transwell assay indicated that the migration and invasion of breast cancer cells were suppressed significantly after the overexpression of circ_0000526, while the reverse effects were observed after knocking down circ_0000526 (Fig. 2F, G). These results further demonstrated that circ_0000526 was associated with the malignance of breast cancer and it could function as a tumor suppressor.
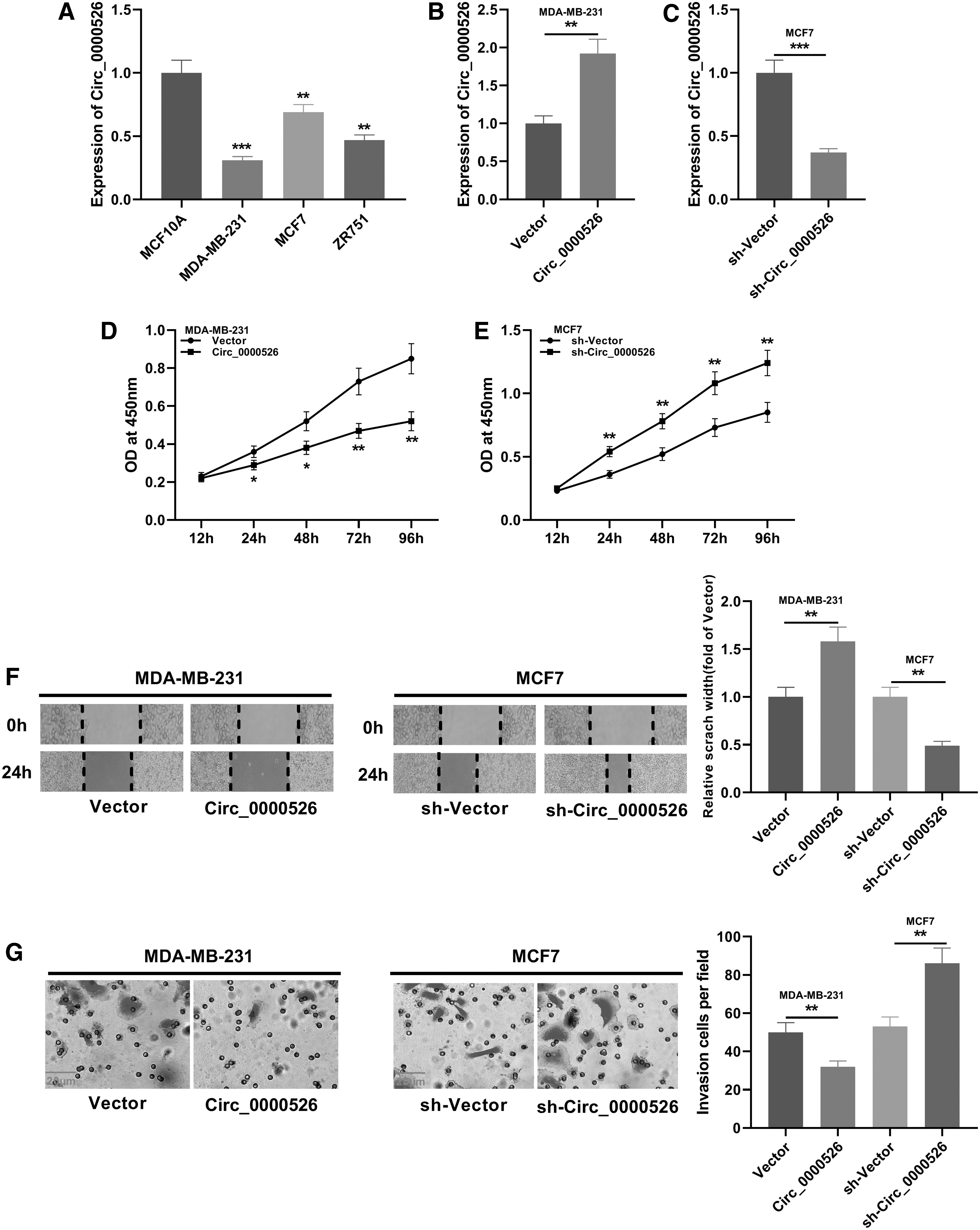
Effects of circ_0000526 on proliferation and metastasis in breast cancer cells.
Circ_0000526 could target miR-492
To further explore the downstream mechanism of circ_0000526, the authors adopted the bioinformatic database to screen the potential targets of circ_0000526. Interestingly, it was found that circ_0000526 contained a site could bind with miR-492 (Fig. 3A). The binding site was further verified using luciferase reporter assay. The results suggested that miR-492 could significantly inhibit the luciferase activity of circ_0000526-wt, but had no significant effect on the luciferase activity of circ_0000526-mut (Fig. 3B). Furthermore, they analyzed the correlations between the expression of circ_0000526 and miR-492 expression in breast cancer tissues. As demonstrated, there was a negative correlation between them (Fig. 3C). RT-PCR results showed that after circ_0000526 was overexpressed or knocked down, the level of miR-492 in breast cancer cell lines was downregulated or upregulated, respectively (Fig. 3D, E). After the transfection of miR-492 mimics or inhibitors, there was no significant change on the level of circ_0000526 in breast cancer cells (Fig. 3F, G). Therefore, the authors concluded that circ_0000526 functions as the upstream molecule of miR-492 and inhibits the expression of miR-492 in breast cancer cell lines.

Circ_0000526 and miR-492 have a targeting relationship.
Effects of regulation of miR-492 on proliferation and metastasis of breast cancer cells
To verify the function of miR-492 in breast cancer, the authors examined the level of miR-492 in breast cancer cell lines, including MDA-MB-231, MCF-7, and ZR751. RT-PCR results manifested that the level of miR-492 in breast cancer cell lines was higher than that in the normal breast epithelial cell line MCF-10A (Fig. 4A). Following that, the authors constructed MCF-7 and MDA-MB-231 cell models using the transfection of mimics and inhibitors of miR-492, respectively (Fig. 4B, C). Besides, CCK-8 assay, wound healing assay, and Transwell assay were used to detect cell proliferation, migration, and invasion. As shown, the proliferation, migration, and invasion of breast cancer cells were significantly promoted after the transfection of miR-492 mimics, while the malignance of breast cancer cells was suppressed using miR-492 inhibitors (Fig. 4D–G). The above results implied that miR-492 served as oncogenic miRNA, which was consistent with a previous report. 14
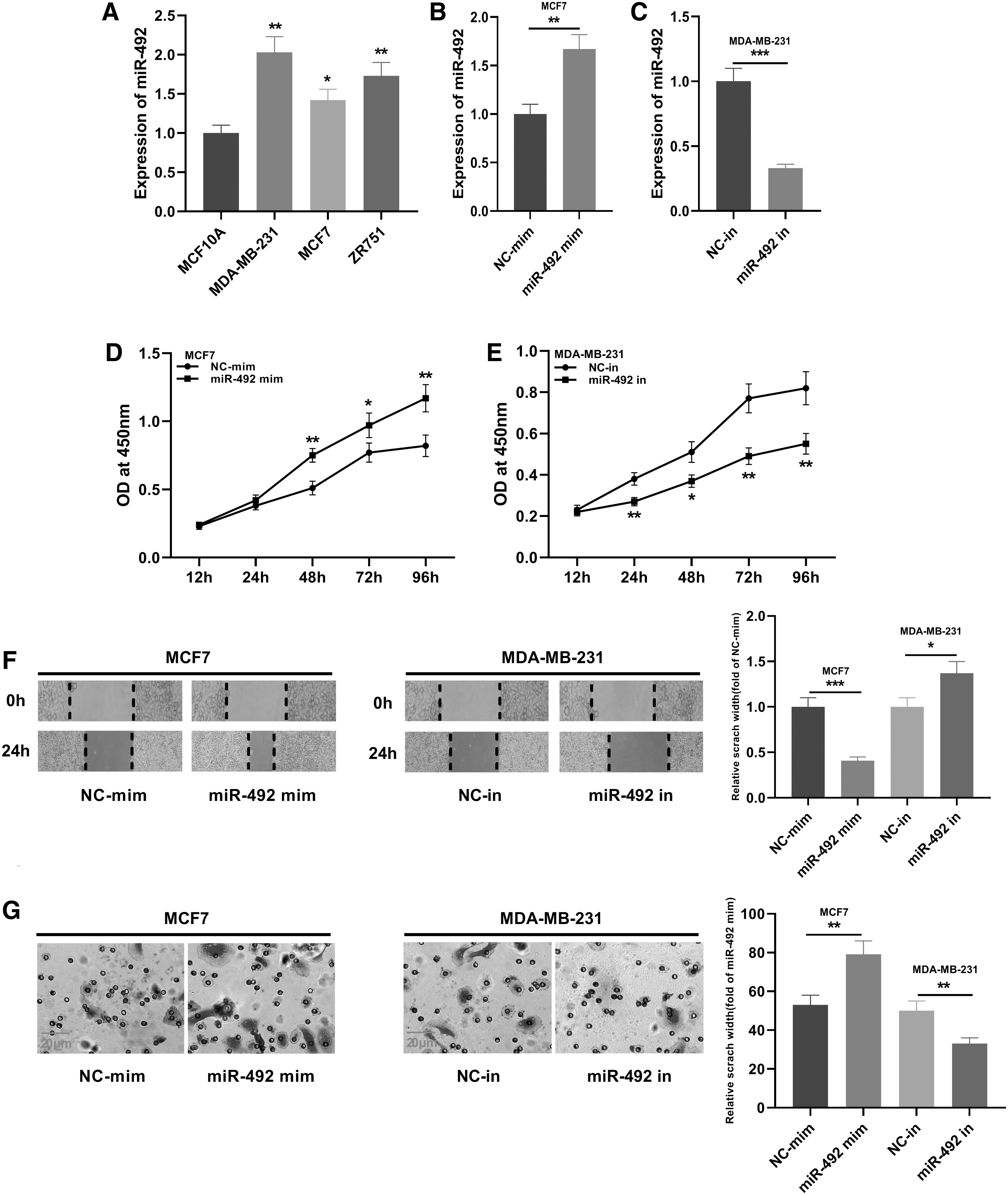
Effects of miR-492 on proliferation and metastasis of breast cancer cells.
SOCS2 functions as the target of miR-492
To explore the role of miR-492 in breast cancer, TargetScan was used to screen targets of miR-492. The results suggested that miR-492 could complement and bind with 3′-untranslated terminal region of SOCS2, indicating that SOCS2 may be the target of miR-492 (Fig. 5A). Therefore, the authors further examined the level of SOCS2 mRNA in breast cancer cell lines using RT-PCR. And the results demonstrated that the level of SOCS2 in breast cancer cell lines was significantly lower than that of normal breast epithelial cell line (Fig. 5B). In addition, the results of dual-luciferase reporter gene assay indicated that miR-492 could significantly inhibit the luciferase activity of SOCS2-wt, but had no significant effect on that of SOCS2-mut (Fig. 5C). They further detected the relationship among SOCS2, miR-492, and circ_0000526 in breast cancer cell lines using Western blot. As demonstrated, after transfection of mimics and inhibitors of miR-492, the levels of SOCS2 in breast cancer cells were downregulated and upregulated, respectively (Fig. 5D). When circ_0000526 was overexpressed and knocked down, the levels of SOCS2 in breast cancer cells were upregulated and downregulated, respectively (Fig. 5E). Importantly, in clinical samples, the expression of SOCS2 was negatively correlated with the expression of miR-492, but positively correlated with the expression of circ_0000526 (Supplementary Fig. S1). Collectively, these results indicated that SOCS2 served as a target of miR-492 in breast cancer cells and was positively regulated by circ_0000526.
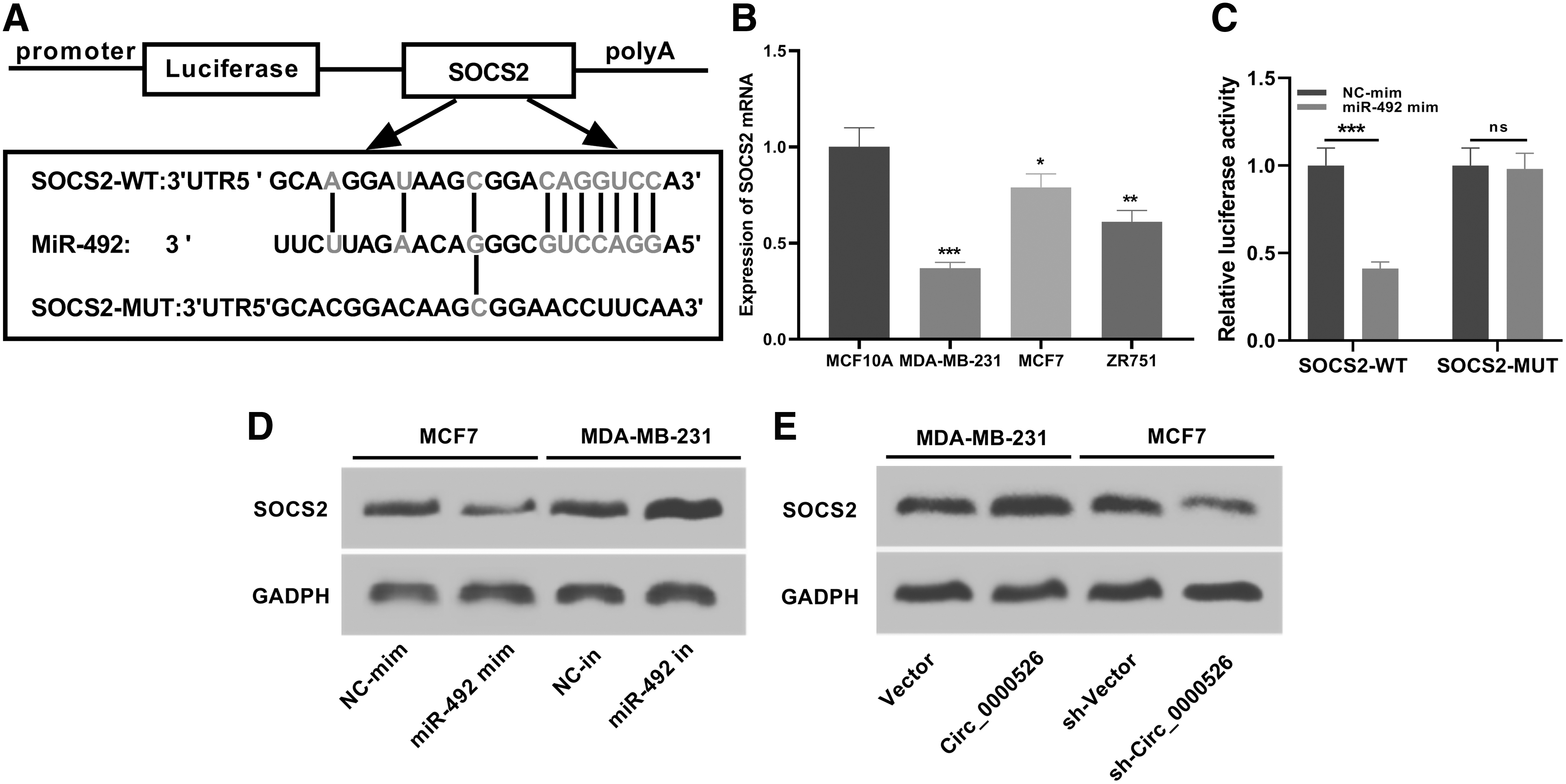
SOCS2 is a target of miR-492.
Circ_0000526 affects the proliferation and metastasis of breast cancer cells through miR-492/SOCS2 axis
To further investigate the role of the circ_0000526/miR-492/SOCS2 axis in the proliferation and metastasis of breast cancer cells, together with miR-492 mimics, circ_0000526 overexpression plasmid was transfected into MDA-MB-231 cells (Fig. 6A–C). As demonstrated, by comparison with the circ_0000526/NC group, the proliferation of MDA-MB-231 cells in the circ_0000526/miR-492 mimics group was significantly facilitated (Fig. 6D). Moreover, the results of wound healing assay and Transwell assay indicated that the migration and invasiveness of MDA-MB-231 cells were significantly rescued after the transfection of miR-492 mimics (Fig. 6E, F). To further verify the effect of circ_0000526/miR-492/SOCS2 axis on breast cancer, the authors transfected shRNA targeting circ_0000526 and SOCS2 overexpression plasmids into MCF-7 cells. Overexpression of SOCS2 did not change the expression of miR-492 (Supplementary Fig. S2A). As shown, SOCS2 overexpression could partially reverse the effect of circ_0000526 knockdown on the proliferation, migration, and invasion of MCF-7 cells (Supplementary Fig. S2B–D). These results further indicated the function of circ_0000526 on suppressing the progression of breast cancer, which was mediated by the miR-492/SOCS2 axis.

Circ_0000526 regulates the proliferation and metastasis of breast cancer cells via the miR-492/SOCS2 axis.
Discussion
Many molecules, which are abnormally expressed in breast cancer tissues and cells, partake in the occurrence and progression of breast cancer. 15 Present studies have proven that abnormal expression of circRNAs may affect the progression of tumors. For instance, in colonic cancer, circRNA PIP5K1A is significantly overexpressed, which can promote cancer progression 16 ; In cervical cancer, circ_0005576 promotes cancer progression via the miR-153/KIF20A axis. 17 Similarly, during the progression of breast cancer, there are a variety of dysregulated circRNAs, which are widely involved in regulating the phenotypes of cancer cells, including proliferation, apoptosis, metastasis, and drug resistance. For example, the upregulation of circ_0008945 in breast cancer facilitates cancer progression. 18 In this study, the authors found that circ_0000526 was downregulated in breast cancer tissues and cells by comparison with the control group. Moreover, the biological function of circ_0000526 in breast cancer was studied through both gain-of-function and loss-of-function models. Therefore, they demonstrated that the overexpression of circ_0000526 could inhibit the proliferation and metastasis of breast cancer cells, while knocking down circ_0000526 could promote the proliferation and metastasis of breast cancer cells. These results suggested that circ_0000526 was a tumor suppressor in breast cancer.
miRNAs also play key roles in cancer progression as tumor promoters or suppressors. 10 For example, miR-214-3p, the expression of which is significantly downregulated in breast cancer tissues, can inhibit the malignant phenotypes of breast cancer cells by targeting survivin protein 19 ; miR-937 functions as oncomiR in breast cancer, which is capable of targeting APAF1 to promote cancer progression. 20 In addition, it has been reported that miR-492 contributes to breast cancer progression by suppressing the expression of SOX7. 14 Consistent with the above report, this study demonstrated that miR-492 expression was significantly upregulated in breast cancer tissues and cells and the transfection of miR-492 mimics promoted the proliferation and metastasis of breast cancer cells, and thus, miR-492 inhibitors had the opposite effects. Importantly, an accumulating number of reports have found that circRNAs can function as molecular sponges, which can adsorb miRNAs, thus affecting the effects of miRNAs on corresponding target genes. 21 For example, circRNA_0025202 inhibits the progression of breast cancer by modulating the miR-182-5p/FOXO3a axis in breast cancer. 22 Circ_0007534 regulates MUC19 by targeting miR-593 to regulate the proliferation and migration of breast cancer cells. 23 Intriguingly, a recent study reports that miR-492 is negatively regulated by circ_0072309 in breast cancer, which partly explains the mechanism of miR-492 overexpression in breast cancer. 24 In this work, the authors validated the targeting relationship between circ_0000526 and miR-492. These results demonstrated that the expression of miR-492 was increased significantly after circ_0000526 was knocked down, while overexpressed circ_0000526 inhibited the expression of miR-492. Considering that circ_0000526 was downregulated in breast cancer, they concluded that circ_0000526 was another crucial circRNA that contributed to the dysregulation of miR-492 in breast cancer.
The gene of SOCS2 is located on human chromosome 12q and its encoded protein consists of 197 amino acid residues. It can be induced by different cytokines and can regulate the biological functions of cytokines, growth factors, and hormones through various signaling pathways such as JAK/STAT. 25 Apart from its role in the regulation of immune responses, its tumor-suppressing role has also been demonstrated in various cancers, including breast cancer. 26,27 In this work, the authors identified SOCS2 as a target of miR-492, which could negatively regulate SOCS2 in breast cancer, and such targeting relationship was also verified in prostate cancer. 11 In addition, they demonstrated that the knockdown of circ_0000526 could suppress the expression of SOCS2, while the overexpression of circ_0000526 promoted the expression of SOCS2. Furthermore, the overexpression of miR-492 reversed the effects of circ_0000526 on inhibiting the proliferation and metastasis of breast cancer cells. These results revealed that circ_0000526 exerted its biological role, in part, by regulating the expression of miR-492 and SOCS2.
Taken together, the authors demonstrated that circ_0000526 was downregulated in breast cancer tissues and cells, and the experiments in vitro indicated that circ_0000526 could inhibit the proliferation and metastasis of breast cancer cells via modulating the miR-492/SOCS2 axis. This study depicted a novel mechanism in the progression of breast cancer, and provided potential biomarkers and therapeutic targets for breast cancer.
Authors' Contributions
Conceived and designed the experiments: P.R. Performed the experiments: W.-B.W., F.-h.R. and X.C. Statistical analysis: W.-B.W. Wrote the article: W.-B.W. and M.H. All authors read and approved the final article.
Data Availability Statement
The data used to support the findings of this study are available from the corresponding author upon request.
Footnotes
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
The Science Foundation of the Health Commission of Hubei Province of China (No. WJ2019M096) and the National Natural Science Foundation of China (No. 81502635).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.