
Abstract
Background:
Non-small cell lung cancer (NSCLC) is the leading cause of cancer deaths all over the world. Exosomes exert central roles in intercellular communication. Circular RNA Rho GTPase activating protein 10 (circARHGAP10) was related to the development of NSCLC. Nevertheless, it was unclear whether circARHGAP10 can be mediated by serum-derived exosomes in NSCLC.
Materials and Methods:
Protein expression of CD63, CD81, family with sequence similarity 83F (FAM83F), glucose transporter 1 (Glut1), and lactate dehydrogenase were evaluated through Western blot analysis. The expression of circARHGAP10, miR-638, and FAM83F was examined by quantitative real-time polymerase chain reaction. Cell proliferation, migration, and invasion were evaluated through 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) or transwell assays. Glucose consumption and lactate production were analyzed with special commercial kits. The relationship between circARHGAP10 or FAM83F and miR-638 was identified by dual-luciferase reporter or RNA immunoprecipitation (RIP) assays. The role of circARHGAP10 in vivo was confirmed through xenograft assay.
Results:
circARHGAP10 was upregulated in NSCLC tissues, cells, and serum-derived exosomes. Serum-derived exosomes boosted the expression of circARHGAP10 in NSCLC cells. circARHGAP10 depletion repressed proliferation, migration, invasion, and glycolysis of NSCLC cells in vitro, and curbed tumor growth in vivo. Also, miR-638 acted as a target of circARHGAP10, miR-638 overexpression overturned circARHGAP10 upregulation-mediated acceleration of proliferation, migration, invasion, and glycolysis of NSCLC cells. Besides, miR-638 targeted FAM83F and FAM83F overexpression abolished miR-638 enhancement-mediated proliferation, migration, invasion, and glycolysis of NSCLC cells.
Conclusions:
Inhibition of serum-derived exosomes-mediated circARHGAP10 curbed NSCLC progression through the miR-638/FAM83F axis.
Introduction
Lung cancer is the main cause of cancer-related deaths worldwide. 1 It mainly includes two subtypes of small-cell lung cancer and non-small cell lung cancer (NSCLC), of which NSCLC accounts for about 80%–85%. 2,3 Although treatment strategies have been greatly improved, the prognosis of NSCLC patients is still poor, with a 5-year survival rate of only 15%. 4 However, high mortality in NSCLC patients might be related to early metastasis. 5,6 Therefore, it is imperative to explore the mechanism of NSCLC progression to provide a basis for developing possible treatment strategies.
Exosomes are small extracellular vesicles with a diameter of about 30–100 nm that have a lipid bilayer member structure. 7 It can be secreted by most cells and contains proteins, lipids, and nucleic acids, which can be transmitted to other cells as signal molecules to change the function of other cells. 8,9 The literature indicated that cancer cells could secrete exosomes, and circular RNAs (circRNAs) have been found to be enriched and stable in exosomes. 10,11
circRNAs are a class of covalent circular endogenous RNAs formed by the 3′ splice donor of pre-mRNA covalently linked to the 5′ splice acceptor in the reverse order. 12 Increased research revealed that circRNAs were involved in the progression of a variety of cancers, including NSCLC. 13 Also, circRNA could enter tumor cells or normal cells through exosomes to participate in tumor progression. 14 –16 For example, it has been reported that circRNA IARS entered human microvascular vein endothelial cells through exosomes and promotes tumor invasion and metastasis. 16 Circular RNA Rho GTPase-activating protein 10 (circARHGAP10), also termed as circRNA has_circ_0008975, is formed by the exon of the ARHGAP10 gene through circularization. Furthermore, circARHGAP10 was pointed out to be unfavorable in NSCLC. 17
Nevertheless, to date, little is known about the biological function of exosomes-mediated circARHGAP10 in NSCLC.
MicroRNAs (miRNAs), a type of noncoding RNAs (∼21 nucleotides in length), are post-transcriptional regulators of gene expression and control multiple cellular processes and organ development in eukaryotes. 18 Studies indicated that miRNAs were implicated in the progression of various tumors and might play different roles in different tumors. 19 MicroRNA-638 (miR-638) was pointed out to be served as a tumor suppressor in a range of cancers, such as breast cancer, 20 gastric cancer, 21 hepatocellular cancer, 22 and colorectal cancer. 23 Nevertheless, whether miR-638 is regulated by circARHGAP10 in NSCLC has not been reported.
Family with sequence similarity 83 (FAM83) is a new protein family that contains eight genes from FAM83A to FAM83H. Members of the FAM83 family have been shown to exhibit oncogenic properties in a variety of human tumors. 24 Family with sequence similarity 83F (FAM83F) was revealed to play a cancer-promoting role in papillary thyroid cancer. 25 Also, FAM83F was connected with the progression of esophageal squamous cell cancer 26 and glioma. 27 Furthermore, FAM83F was also related to the development of NSCLC. 28,29 However, it is unclear whether FAM83F is modulated by miR-638 and circARHGAP10 in NSCLC.
Hence, the authors examined circARHGAP10 expression in serum and tissue patients with NSCLC. Moreover, the influence of serum exosomes on the expression of circARHGAP10 in NSCLC cells was explored. Besides, the effects of circARHGAP10 on proliferation, migration, invasion, and glycolysis of NSCLC cells and the molecular mechanism of the circARHGAP10/miR-638/FAM83F axis were surveyed.
Materials and Methods
Subject
This study was delegated by the Ethics Committee of the High-Tech Hospital, First Affiliated Hospital of Nanchang University. Forty NSCLC patients and 40 healthy volunteers were obtained from the High-Tech Hospital, First Affiliated Hospital of Nanchang University. Whole blood (20 mL) was drawn from NSCLC patients and healthy volunteers, and 40 paired NSCLC tissues and neighboring normal tissues were acquired from NSCLC patients. After storage at room temperature for 2 h, the blood samples were centrifuged at 1200 g for 20 min to obtain the serum for subsequent studies. All tissue samples and serums were stored in liquid nitrogen until used. Informed consents were signed by the NSCLC patients and healthy volunteers.
Cell culture and transfection
Normal lung cell BEAS-2B and human NSCLC cell lines A549 and H292 were acquired from the American Type Culture Collection (Rockville, MD). Roswell Park Memorial Institute (RPMI)-1640 medium (Sigma, St. Louis, MO) supplemented with fetal bovine serum (FBS, 10%) (Sigma), streptomycin (100 g/mL; Sigma), and penicillin (100 U/mL; Sigma) was applied to culture BEAS-2B, A549, and H292 cells. Cells were maintained in a humidified atmosphere with 5% CO2 at 37°C.
The lentivirus-mediated circARHGAP10 overexpression vector (circARHGAP10) or negative control vector (pLVX) was obtained from GenePharma (Shanghai, China). Small interference RNA targeting circARHGAP10 (si-circARHGAP10) and si-FAM83F and their negative control (si-NC) were procured from GenePharma. MiR-638 mimic and inhibitor and their negative control (miRNA NC and inhibitor-NC) were purchased from GenePharma. The sequence of FAM83F was amplified and cloned into pcDNA vector (pc-NC) to acquire the overexpression vector for FAM83F (pc-FAM83F). Transfection of vectors or oligonucleotides was carried out through Lipofectamine 3000 reagent (Invitrogen, Carlsbad, CA).
Exosome isolation
After washing with phosphate buffer, A549 and H292 cells were cultivated in exosome-free medium (Sigma) for 48 h. Following this, the culture medium was collected and centrifuged to obtain a supernatant. Afterward, the supernatant was filtered through a 0.22 μm polyvinylidene difluoride filter (Millipore, Bedford, MA). Then, the serum or filter medium was mixed with exoquick exosome precipitation solution (System Biosciences, Mountain View, CA) and stored at 4°C for 24 h. Next, the exosome pellets obtained by centrifugation at 1500 g for 30 min were suspended in the exosome-free medium or phosphate buffer for subsequent analysis. Nanoparticle-tracking analysis was performed through ZetaView (Particle Metrix, Meerbusch, Germany) to assess the exosome concentration and size distribution.
Transmission electron microscopy
The transmission electron microscope (TEM) (JEM-2100) from JEOL (Tokyo, Japan) was employed to examine the morphology of extracted exosome. A drop of exosome suspension was placed on a copper net and incubated for 1 min. After drying at 65°C, the morphology of the exosomes was observed by JEM-2100 TEM at 80 kV.
Western blot analysis
The Western blot analysis was executed as previously described. 30 NSCLC tissues and neighboring normal tissues, as well as cells, were lysed using radioimmunoprecipitation assay (RIPA) lysis buffer (Invitrogen). ECL chemiluminescence detection reagents (GE Healthcare Life Sciences, Pittsburgh, PA) were applied to develop signals and visualize on X-ray film (Fujifilm, Tokyo, Japan). The primary antibodies were displayed as follows: anti-FAM83F (sc-102517; Santa Cruz Biotechnology, Santa Cruz, CA), anti-CD63 (sc-5275; Santa Cruz Biotechnology), anti-CD81 (sc-166029; Santa Cruz Biotechnology), anti-glucose transporter 1 (Glut1) (sc-377228; Santa Cruz Biotechnology), anti-lactate dehydrogenase (LDH) (sc-133123; Santa Cruz Biotechnology), and anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (sc-47724; Santa Cruz Biotechnology). GAPDH was regarded as a loading control.
Quantitative real-time polymerase chain reaction
Total RNA of the exosomes, serum, NSCLC tissues, and neighboring normal tissues and cells were extracted through TRIzol reagent (Invitrogen). Total RNA (2 μg) was digested with RNase R (3 U/μg; Epicentre Technologies, Madison, WI) at 37°C for 15 min for the assessment of the stability of circARHGAP10. The nuclear and cytoplasmic fractions of NSCLC cells were separated through the PARIS Kit (Life Technologies, Grand Island, NY) for analysis of the subcellular localization of circARHGAP10. Moloney Murine Leukemia Virus (M-MLV) First-Strand Kit (Life Technologies) or miRNA Reverse Transcription Kit (TaKaRa, Dalian, China) was applied to generate the first-strand complementary DNA (cDNA) for circARHGAP10, FAM83F, ARHGAP10, and miR-638.
Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using SYBR Premix Ex Taq (TaKaRa). The levels of circARHGAP10, FAM83F, ARHGAP10, and miR-638 were figured with the 2−ΔΔCt method, and GAPDH or U6 small nuclear RNA (snRNA) was used as an internal control. The primers used were displayed as below: circARHGAP10: 5′-TCTTTTTGAAAGAATGTACCAAGAG-3′ (F) and 5′-GGGATCATGTTGAACTTCTTTG-3′ (R); ARHGAP10: 5′-TGTGGAACCTATGCTGTCAT-3′ (F) and 5′-GACTTGCTCGTTTGTGGTC-3′ (R); FAM83F: 5′-ACAGAACGTAGAGCCCTTTGACA-3′ (F) and 5′-TCGAGCCACAGTGGAGGAGTA-3′ (R); GAPDH: 5′-GACTCCACTCACGGCAAATTCA-3′ (F) and 5′-TCGCTCCTGGAAGATGGTGAT-3′ (R) miR-638: 5′-ATCCAGTGCGTGTCGTG-3′ (F) and 5′-TGCTAGGGATCGCGGGCGGGTG-3′ (R); as well as U6 snRNA: 5′-GCTCGCTTCGGCAGCACA-3′ (F) and 5′-GAGGTATTCGCACCAGAGGA-3′ (R).
3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide assay
The 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay was performed to examine cell proliferation. After transfection for 48 h, A549 and H292 cells (2.0 × 103) were seeded in 96-well plates (Corning Costar, Corning, NY) and cultured for 24, 48, and 72 h. Afterward, MTT (20 μL; Sigma) was added to each well and incubated for 4 h, and dimethyl sulfoxide (200 μL) was supplemented to each well for the dissolution of the precipitate. The color reaction at 490 nm was analyzed by the Microplate Absorbance Reader (Thermo Fisher Scientific, Waltham, MA).
Cell colony formation assay
A549 and H292 cells were transfected with si-circARHGAP10 or si-NC. For colony formation assay, the cells (1 × 102 cells/10 cm dish) were seeded in six-well plates containing agar (Bio-Rad, Hercules, CA), and the medium was replaced every 2 d. After 10 d, the cells were fixed with methanol (3.7%) and stained with Crystal Violet (0.1%; KeyGen, Jiangsu, China). The colonies containing at least 50 cells were counted and photographed with the light microscope (Olympus, Tokyo, Japan).
Transwell assay
The migration capacity of transfected A549 and H292 cells was determined with the transwell chamber (8 μm; Corning Costar). In brief, RPMI-1640 medium with FBS (10%) was added to the lower part of the transwell chamber. Also, RPMI-1640 medium with transfected A549 and H292 cells (5 × 104) was replenished to the upper part of the transwell chamber. Next, the cells on the upper surface of the membrane were removed with a cotton swab and the cells on the lower surface of the membrane were fixed with methanol (100%) and stained with Crystal Violet (0.1%). The invasion ability of transfected A549 and H292 cells was evaluated through the same method as the migration assay, but the upper chamber was coated with a Matrigel matrix (BD Biosciences, San Jose, CA). The migrated and invasive cells were counted with a light microscope (Olympus).
Measurement of glucose consumption and lactate production
Transfected A549 and H292 cells were cultured in RPMI-1640 medium for 24 h. Next, the cell supernatant was collected and the glucose level and lactate production were detected using a Glucose Assay Kit (Sigma) or a Lactic Acid Assay Kit (Sigma) according to the manufacturer's instructions.
Dual-luciferase reporter assay
The binding sites between circARHGAP10 or FAM83F and miR-638 were predicted with circRNA Interactome or Targetscan Online Softwares. After that, the fragments of wild-type circARHGAP10 and 3′ Untranslated Regions (UTR) of FAM83F (containing predicted miR-638-binding sites), as well as mutant circARHGAP10 and 3′UTR of FAM83F were amplified and cloned into the pGL3-control vector (Promega, Madison, WI) to construct the luciferase reporter vectors. The 293T cells were cotransfected with miR-638 mimic or miRNA NC and a luciferase reporter vector by using Lipofectamine 3000 reagent. The luciferase intensity of luciferase reporter vector was determined through the Luciferase Reporter Assay Kit (Promega).
RNA immunoprecipitation assay
A Magna Nuclear RIP™ Kit (Millipore) was used for RNA immunoprecipitation (RIP) analysis. In short, RIPA buffers contain RNase inhibitor and protease inhibitor cocktail was used to lyse A549 and H292 cells. Then, RIP buffer containing magnetic beads coupled with immunoglobulin G (IgG) control or human anti-Ago2 was incubated with the cell lysate. Proteins were digested with protease to obtain immunoprecipitated RNA. In the end, the relative level of extracted RNA was examined by qRT-PCR.
Xenograft assay
Eight BALB/c nude mice (4–8 weeks old) were obtained from Shanghai Experimental Animal Center (Shanghai, China) for in vivo tumor xenograft assay. Briefly, lentivirus-mediated small hairpin RNA targeting circARHGAP10 (sh-circARHGAP10) or its negative control (sh-NC) was transfected into H292 cells. Then, transfected H292 cells were subcutaneously injected into the dorsal side of nude mice (4 mice/group). The tumor volume was measured with a digital caliper every 4 d from the seventh day and calculated with the equation: Volume = (length × width 2 )/2. On day 27, xenograft mice were euthanized to evaluate tumor tissue weight and other follow-up studies. The animal experiment was ratified by the Ethics Committee of the High-Tech Hospital, First Affiliated Hospital of Nanchang University.
Statistical analysis
SPSS 19.0 software (SPSS, Chicago, IL) and GraphPad Prism 6.0 (GraphPad, San Diego, CA) were executed for statistical analysis. The in vitro experiments of this study were performed at least three times. Data were exhibited as mean ± standard deviation. Differences between two groups were assessed through Student's t-test and one-way analysis of variance was applied to compare the differences among more groups. p < 0.05 was deemed statistically significant.
Results
circARHGAP10 was upregulated in NSCLC tissues, cells, and serum-derived exosomes
Exosomes can transport nucleotides, lipids, proteins, and other substances to recipient cells. 31 In this study, they first extracted exosomes from serum of NSCLC patients and healthy volunteers to probe into the underlying role of exosomal-circARHGAP10 in NSCLC. TEM revealed that the shape of serum exosomes in NSCLC patients and healthy volunteers was round, as displayed in Figure 1A. Furthermore, NTA analysis exhibited that the median size of NSCLC serum-derived exosomes was 109.8 nm (Fig. 1B), and Western blot analysis presented that exosomal marker proteins CD63 and CD81 were presented in the serum exosomes of patients with NSCLC and healthy volunteers (Fig. 1C). Results of qRT-PCR manifested that the expression of circARHGAP10 was higher in the serum of NSCLC patients compared with the healthy volunteers (Fig. 1D).
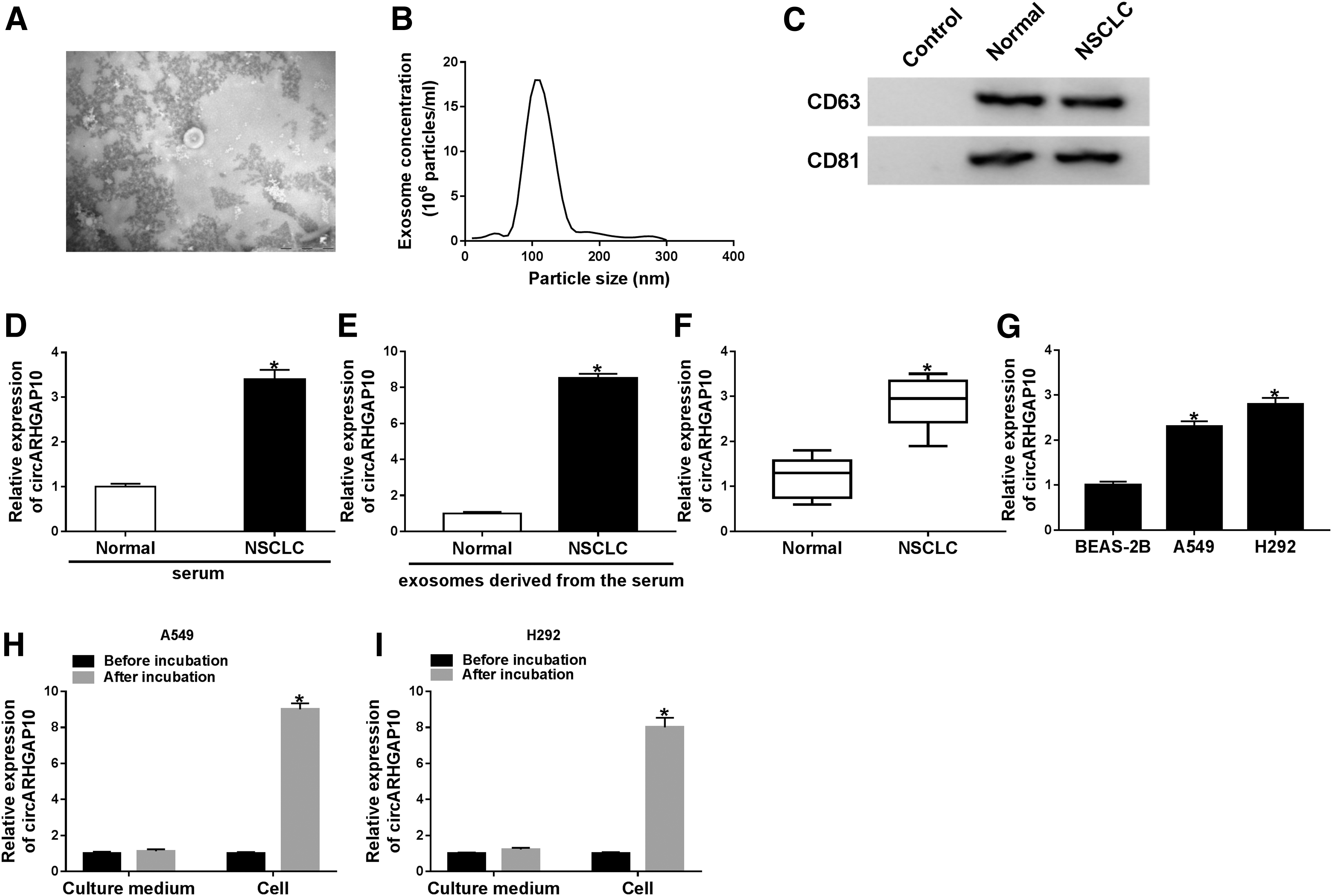
Expression levels of circARHGAP10 in NSCLC tissues, cells, and serum-derived exosomes.
Also, circARHGAP10 level in the serum exosomes of NSCLC patients was evidently elevated in contrast to the healthy volunteers (Fig. 1E). The expression levels of circARHGAP10 in NSCLC tissues and cells (A549 and H292) were abnormally enhanced when compared with the neighboring normal tissues or BEAS-2B cells (Fig. 1F, G). After incubation with NSCLC serum-derived exosomes, the expression of circARHGAP10 was conspicuously augmented in A549 and H292 cells compared with normal culture, implying that NSCLC serum-derived exosomes accelerated the expression of circARHGAP10 in NSCLC cells (Fig. 1H, I). These results demonstrated that NSCLC serum-derived exosome-mediated elevation of circARHGAP10 might be related to the progression of NSCLC.
circARHGAP10 facilitated proliferation, migration, invasion, and glycolysis of NSCLC cells
To further confirm the role of circARHGAP10 in NSCLC, they first assessed the resistance of circARHGAP10 for RNase R. The results suggested that circARHGAP10 possessed a stronger resistance to RNase R than ARHGAP10 in A549 and H292 cells, indicating this RNA existed in a circular form (Fig. 2A, B). Besides, circARHGAP10 expression level was drastically declined in A549 and H292 cells transfected with si-circARHGAP10, whereas the opposite result was observed in A549 and H292 cells transfected with circARHGAP10 (Fig. 2C, D). Also, qRT-PCR showed that circARHGAP10 was obviously localized within the cytoplasm in A549 and H292 cells (Fig. 2E, F).

Effects of circARHGAP10 on proliferation, migration, invasion, and glycolysis of NSCLC cells.

MTT assay demonstrated that circARHGAP10 enhancement expedited the proliferation of A549 cells, whereas circARHGAP10 inhibition repressed the proliferation of H292 cells (Fig. 2G, H), and the downregulation of circARHGAP10 constrained the colony formation of A549 and H292 cells (Supplementary Fig. S1). Transwell assay exhibited that migration and invasion were apparently promoted in circARHGAP10-elevated A549 cells. However, they observed remarkable repression of migration and invasion in H292 cells with reduced circARHGAP10 (Fig. 2I, J).
Additionally, glucose consumption and lactate production were dramatically boosted in circARHGAP10-elevated A549 cells, whereas inhibition of circARHGAP10 reduced glucose consumption and lactate production in H292 cells (Fig. 2K, L). In addition, the results of Western blot analysis presented that the expression levels of Glut1 and LDH were enhanced by circARHGAP10 overexpression in A549 cells, whereas Glut1 and LDH expression levels were decreased by circARHGAP10 inhibition in H292 cells (Fig. 2M). Taken together, these data manifested that circARHGAP10 could contribute to cell proliferation, migration, invasion, and glycolysis in NSCLC cells.
miR-638 acted as a target for circARHGAP10
To understand how circARHGAP10 affected cell proliferation, migration, invasion, and glycolysis in NSCLC, they employed circRNA Interactome Online Software to predict the target of circARHGAP10. As displayed in Figure 3A, miR-638 owned the binding sites for circARHGAP10. Then, they constructed the luciferase reporter vector containing circARHGAP10 or mutated circARHGAP10 to verify the relationship between circARHGAP10 and miR-638. Dual-luciferase reporter assay manifested that miR-638 mimic distinctly declined the luciferase activity of 293T cells transfected with the luciferase reporter vector containing wild-type circARHGAP10 sequence. However, miR-638 mimic did not have any influence on the luciferase reporter vector containing mutant circARHGAP10 sequence (Fig. 3B). Furthermore, results of RIP demonstrated that both circARHGAP10 and miR-638 were enriched in Ago2-containing beads compared with IgG immunoprecipitates in A549 and H292 cells (Fig. 3C, D). Also, circARHGAP10 overexpression repressed the expression of miR-638 in both A549 and H292 cells (Fig. 3E), and miR-638 was upregulated in A549 and H292 cells transfected with miR-638 mimic (Fig. 3F). Moreover, miR-638 expression levels were elevated in circARHGAP10-silenced A549 and H292 cells (Fig. 3G). The expression of miR-638 was reduced in A549 and H292 cells with miR-638 inhibitor (Fig. 3H).

circARHGAP10 acted as a sponge for miR-638.
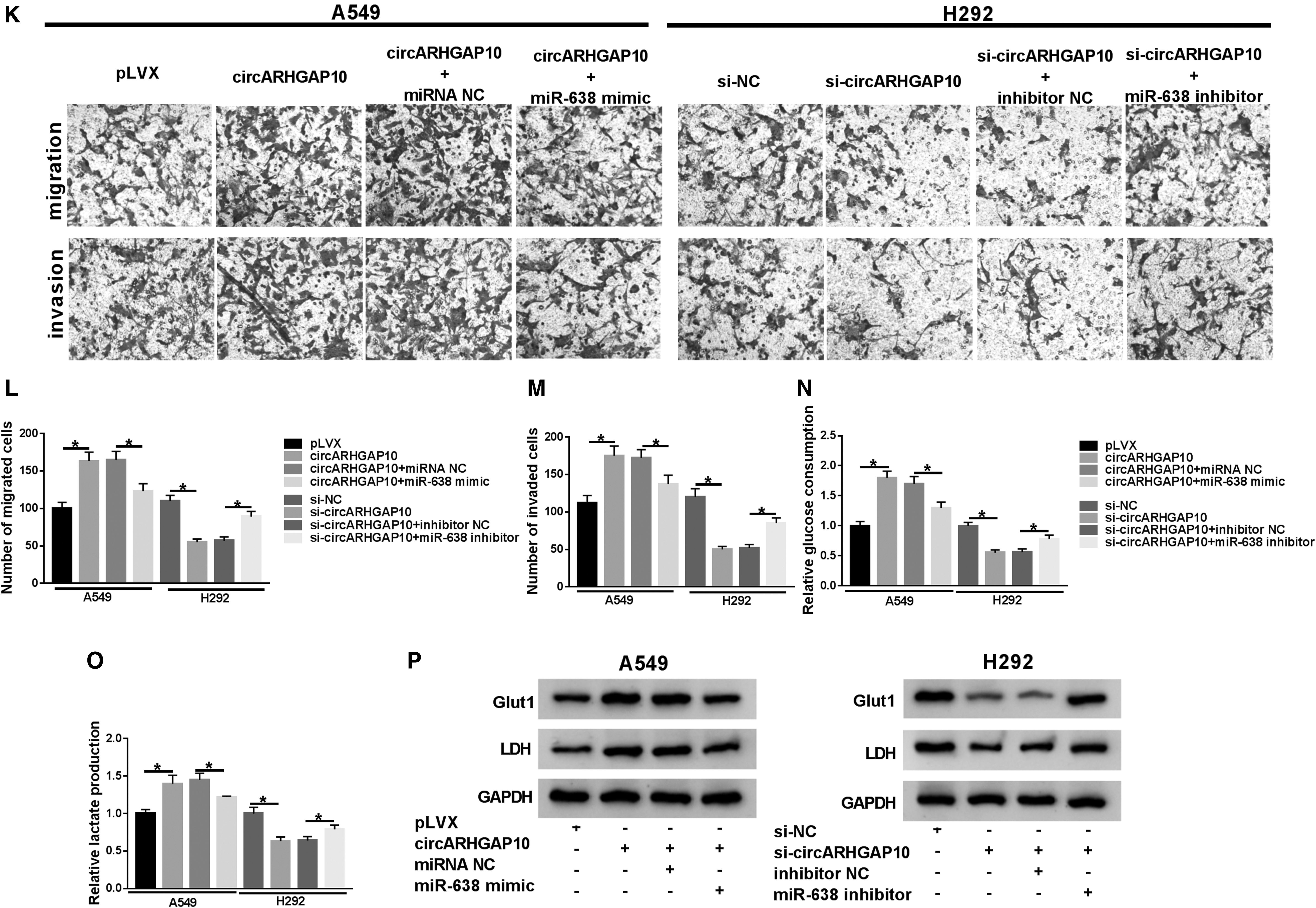
Subsequently, the effects of miR-638 on circARHGAP10-mediated proliferation, migration, invasion, and glycolysis of NSCLC cells were investigated. MTT assay exhibited that the promotion of proliferation of A549 cells induced by circARHGAP10 upregulation was reversed by miR-638 mimic, and the repressive effect of circARHGAP10 downregulation on proliferation of H292 cells was overturned by miR-638 inhibitor (Fig. 3I, J). Transwell assay also showed that enhanced miR-638 expression abolished the acceleration of migration and invasion of A549 cells caused by circARHGAP10 enhancement, and miR-638 inhibition restored the repressive effects of circARHGAP10 silencing on migration and invasion of H292 cells (Fig. 3K–M).
Besides, both the elevation of glucose consumption and lactate production of A549 induced by circARHGAP10 augmentation were recovered by the overexpression of miR-638. Additionally, the downregulation of miR-638 reversed circARHGAP10 repression-mediated reduction of glucose consumption and lactate production of H292 cells (Fig. 3N, O). In addition, the upregulation of Glut1 and LDH in circARHGAP10-elevated A549 cells were reversed by miR-638 overexpression, and the downregulation of Glut1 and LDH in H292 cells mediated by circARHGAP10 silencing were restored by the introduction of miR-638 inhibitor (Fig. 3P). Collectively, these results revealed that circARHGAP10 targeted miR-638 to mediate proliferation, migration, invasion, and glycolysis of NSCLC cells.
FAM83F was a target of miR-638
In view of the above results, they sought for miR-638 targets to gain a deeper understanding of the molecular mechanisms regulated by circARHGAP10. TargetScan Online Software presented that FAM83F possessed the binding sites with miR-638 (Fig. 4A). Then, they established the luciferase reporter vector containing 3′UTR of FAM83F or mutant 3′UTR of FAM83F sequences. Results of dual-luciferase reporter assay displayed that the luciferase activity of luciferase reporter vector containing 3′UTR of FAM83F in 293T cells was prominently decreased by miR-638 mimic, whereas the luciferase activity of luciferase reporter vector containing mutant 3′UTR of FAM83F was not distinctly different (Fig. 4B). Moreover, the mRNA and protein levels of FAM83F in A549 and H292 cells transfected with miR-638 mimic were lower than the miRNA NC group (Fig. 4C, D), whereas the mRNA and protein levels of FAM83F were higher in A549 and H292 cells transfected with miR-638 inhibitor (Fig. 4E, F).
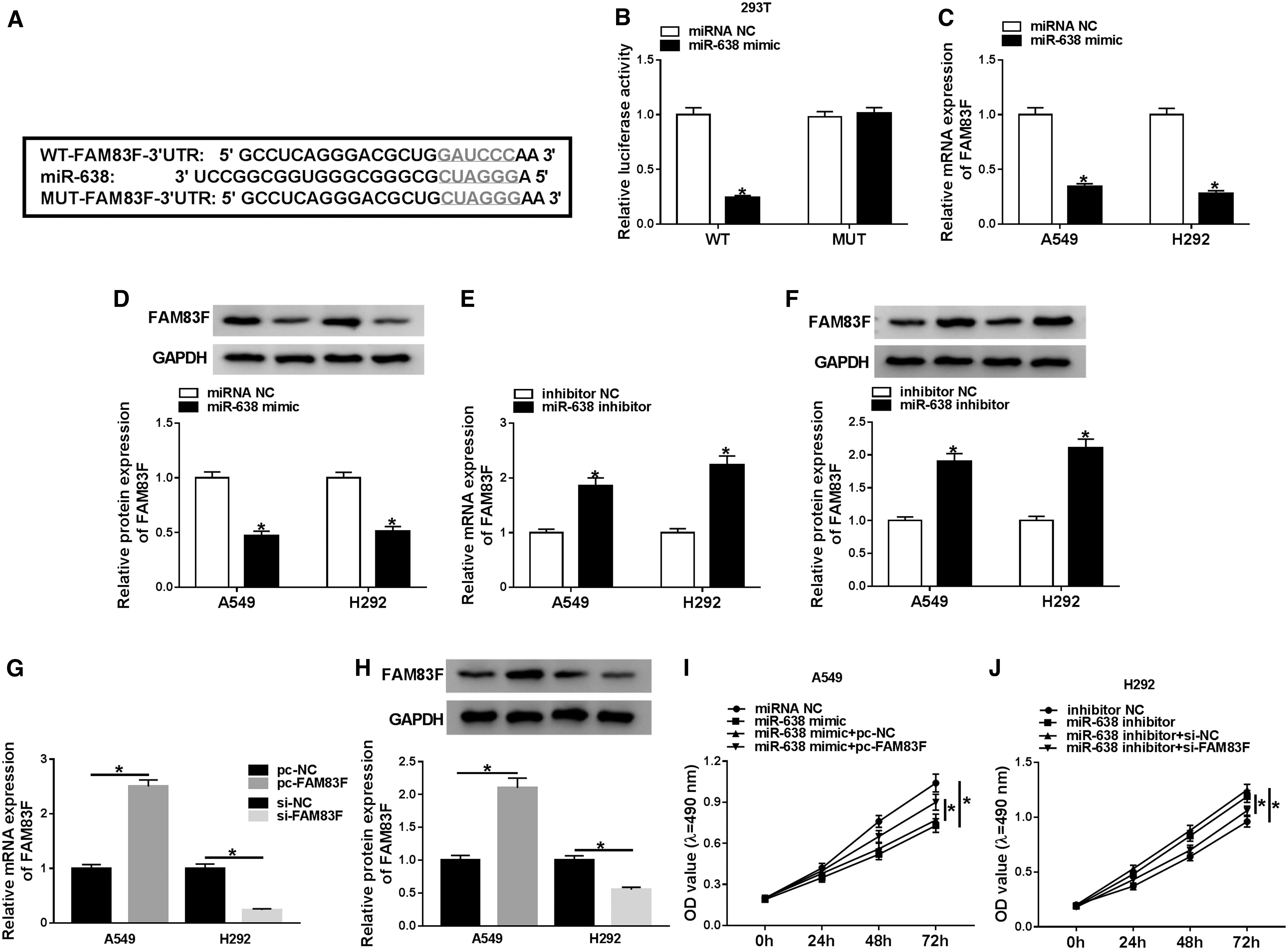
FAM83F acted as a target for miR-638.

In addition, FAM83F mRNA and protein levels were enhanced in A549 cells transfected with pc-FAM83F compared with the pc-NC group and were repressed in H292 cells transfected with si-FAM83F in contrast to the si-NC group (Fig. 4G, H). Furthermore, MTT assay indicated that miR-638 enhancement curbed proliferation of A549 cells, whereas this repression was abrogated by FAM83F upregulation (Fig. 4I). Oppositely, miR-638 silencing expedited the proliferation of H292 cells, whereas this promotion was overturned by FAM83F inhibition (Fig. 4J). Similarly, the elevation of FAM83F reversed the inhibition of migration and invasion of A549 cells induced by miR-638 upregulation; FAM83F depletion restored miR-638 silencing-mediated facilitation of migration and invasion of H292 cells (Fig. 4K–M).
Besides, the inhibitory effects of miR-638 mimic on glucose consumption and lactate production of A549 were overturned by FAM83F introduction, and the stimulative impacts of miR-638 knockdown on glucose consumption and lactate production of H292 cells were restored by FAM83F inhibition (Fig. 4N, O). Also, the elevation of FAM83F recovered the inhibitory effects of miR-638 overexpression on the expression levels of Glut1 and LDH in A549 cells, and FAM83F depletion abolished miR-638 repression-mediated influence on the expression of f Glut1 and LDH in H292 cells (Fig. 4P). Therefore, these data suggested that miR-638 played its role on proliferation, migration, invasion, and glycolysis of NSCLC cells through FAM83F.
circARHGAP10 modulated the expression of FAM83F by sponging miR-638
Considering that FAM83F served as a target for miR-638, the authors inferred that circARHGAP10 modulated the expression of FAM83F through miR-638 in NSCLC cells. Results of qRT-PCR and Western blot analysis manifested that circARHGAP10 elevation boosted the expression levels of FAM83F mRNA and protein in A549 cells, whereas this enhancement was abolished by the knockdown of FAM83F. Moreover, FAM83F overexpression reversed circARHGAP10 inhibition-mediated reduction of FAM83F mRNA and protein levels of H292 cells (Fig. 5A, B). In addition, the upregulation of miR-638 abrogated the augmentation of FAM83F mRNA and protein levels in A549 cells caused by circARHGAP10 introduction, but the downregulation of FAM83F mRNA and protein levels in H292 cells induced by circARHGAP10 depletion were restored by miR-638 knockdown (Fig. 5C, D). In sum, these data suggested that FAM83F was regulated by circARHGAP10 through sponging miR-638.
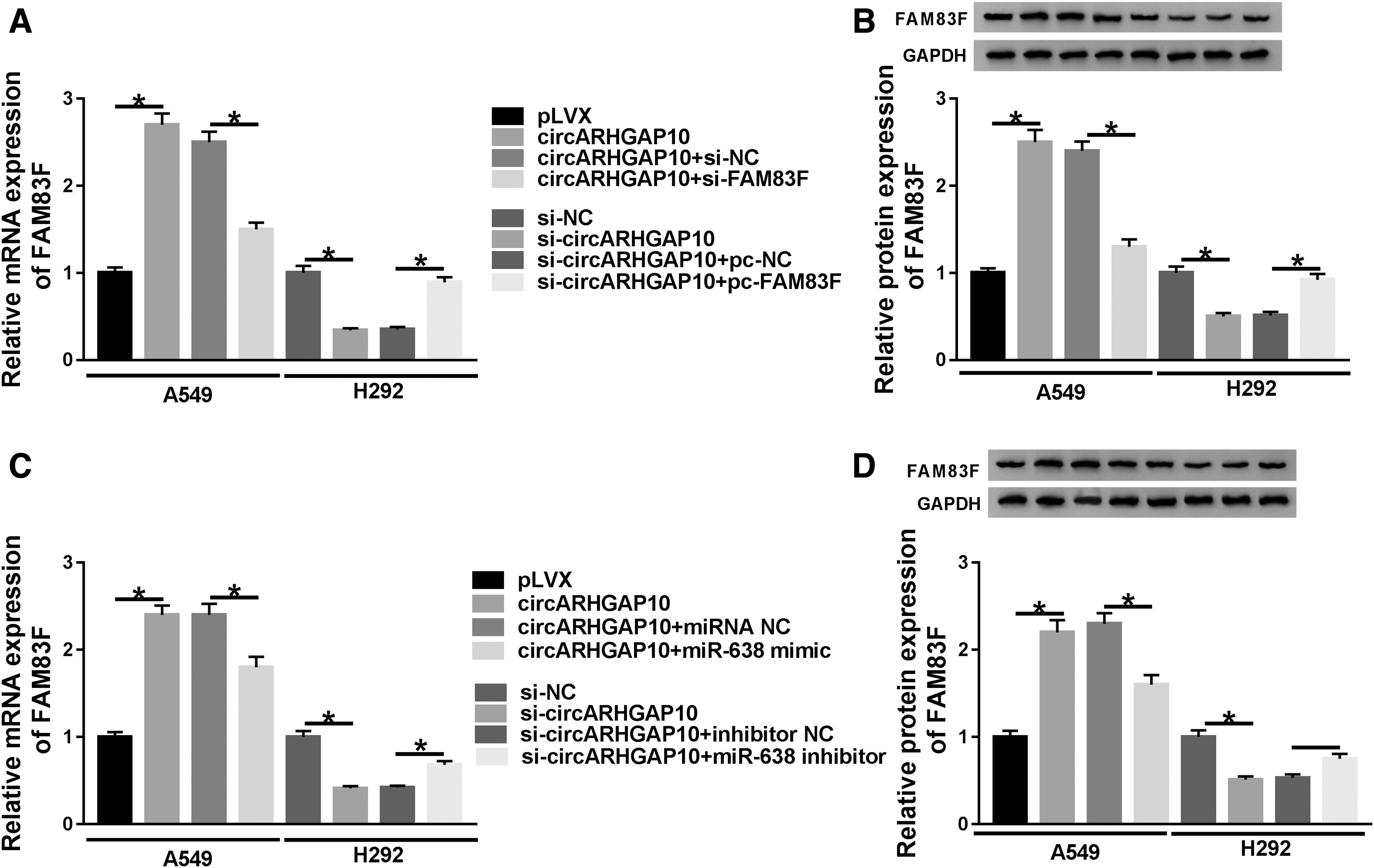
circARHGAP10 regulated FAM83F expression by targeting miR-638.
circARHGAP10 depletion repressed tumor growth in vivo
Given that circARHGAP10 impeded the proliferation, migration, invasion, and glycolysis of NSCLC cells in vitro, they further verified the role of circARHGAP10 in vivo through xenograft assay. H292 cells stably transfected with sh-circARHGAP10 or sh-NC were subcutaneously inoculated into the nude mice. They discovered a marked suppression of tumor volume in the sh-circARHGAP10 group compared with the sh-NC group (Fig. 6A). Moreover, the weight of tumor of the sh-circARHGAP10 group was lower than the control group (Fig. 6B). Furthermore, compared with the sh-NC group, circARHGAP10 was downregulated and miR-638 was upregulated in tumor tissues of sh-circARHGAP10 group (Fig. 6C, D). Besides, the expression levels of FAM83F mRNA and protein were declined in tumor tissues of sh-circARHGAP10 group in contrast to the control group (Fig. 6E, F). Collectively, these findings indicated that circARHGAP10 knockdown curbed the growth of tumor in vivo.
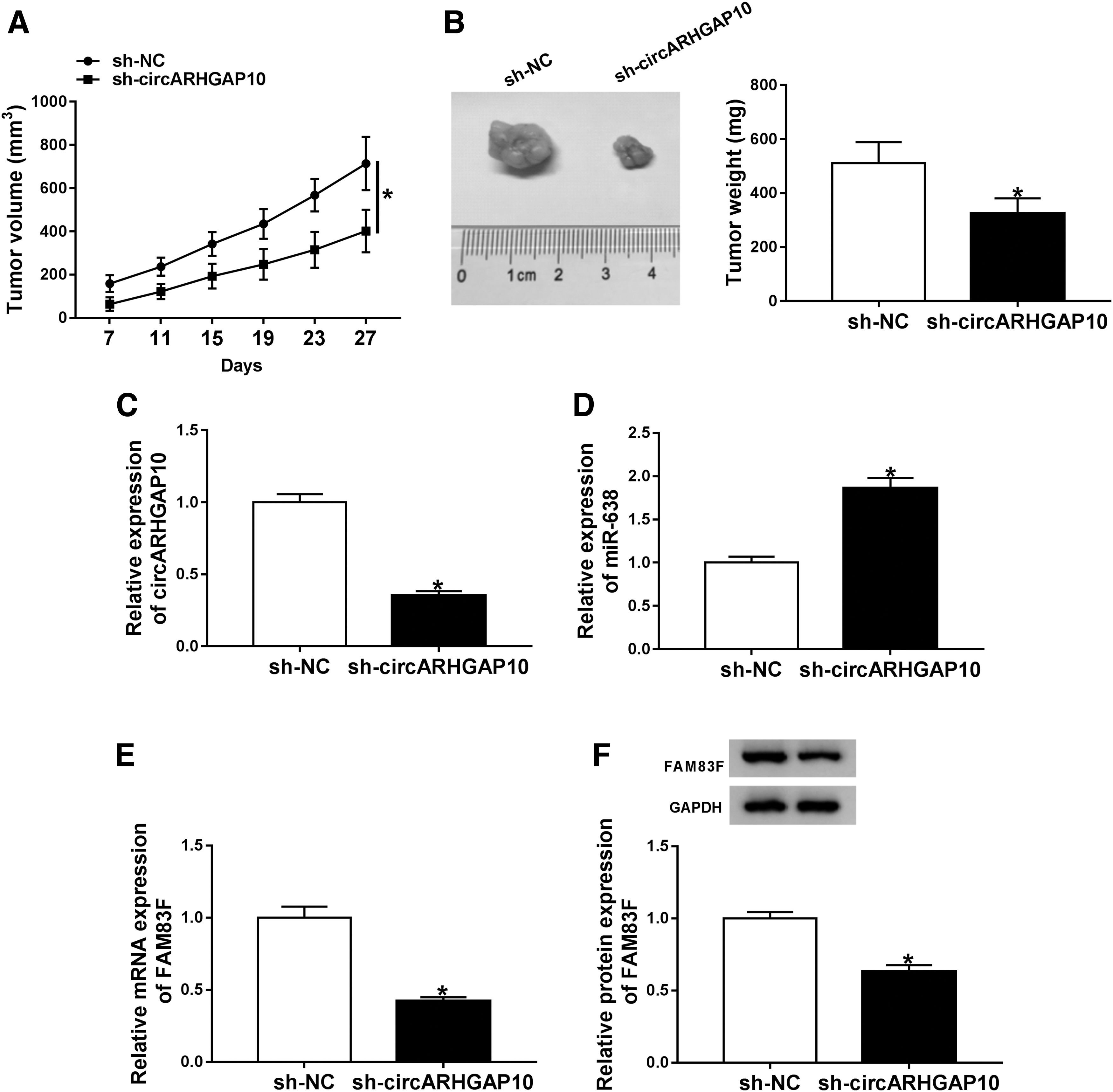
circARHGAP10 downregulation suppressed the growth of tumor in vivo.
Discussion
Originally, exosomes were deemed to be a mechanism for removing unnecessary proteins from cells. 32 However, there is increasing evidence which manifests that exosomes play a vital in the progression of tumors. 8 Metastasis is a complex cascade process that includes cancer cell survival, invasion, and attachment and colonization of host organs, and exosomes have been shown to affect this process. 33 Therefore, exploring the molecular mechanisms of exosomes can be used as a possible target for NSCLC liquid biopsy or as a therapeutic method.
Rising evidence demonstrated that circRNAs are connected with tumor microenvironment and metastasis of NSCLC. 34 –36 For instance, depletion of circRNA_100146 accelerated cell apoptosis and repressed cell invasion and proliferation in NSCLC cells. 35 Moreover, circRNA PTPRA impeded metastasis and epithelial/mesenchymal transitioning through targeting miR-96-5p in NSCLC. 36 Also, circRNA PVT1 silencing promoted cell apoptosis and suppressed cell proliferation in NSCLC cells through reducing expression through miR-497. 37
In this study, enhanced circARHGAP10 expression was founded in NSCLC tissues, cells, and serum-derived exosomes. Moreover, exosomes from NSCLC patient's serum boosted circARHGAP10 expression in NSCLC cells. Inhibition of circARHGAP10 suppressed cell proliferation, migration, invasion, and glycolysis of NSCLC cells in vitro, and inhibited tumor growth in vivo. Jin et al. claimed that circARHGAP10 suppression curbed glycometabolism through downregulation GLUT1 in vitro and in vivo, and repressed metastasis and proliferation through regulating GLUT1 expression through miR-150-5p in NSCLC. 17 These data revealed that serum-derived exosomes could mediate the progression of NSCLC through modulating circARHGAP10 expression.
Considering that circARHGAP10 acted as a sponge for miR-150-5p involved in the development of NSCLC, 17 they also uncovered miR-638 as a target for circARHGAP10 through dual-luciferase reporter assay and RIPA in NSCLC cells. Studies indicated that miR-638 was downregulated in a range of tumors. Li et al. suggested that miR-638 was participated in the progression of breast cancer through acting as a tumor suppressor. 20 Furthermore, miR-638 was also pointed out to induce cell apoptosis and repress cell cycle, colony formation, and proliferation by inactivation of the MEK1/2-ERK1/2 pathway by downregulating MECP2. 21 Xia et al. indicated that reduced miR-638 expression facilitated cell epithelial/mesenchymal transitioning, invasion, and proliferation in NSCLC. 38
In the current study, miR-638 was negatively regulated by circARHGAP10, and miR-638 overexpression reversed circARHGAP10 introduction-mediated facilitation of proliferation, migration, invasion, and glycolysis of NSCLC cells. In line with Xia et al., 38 miR-638 acted as a suppressor gene in NSCLC.
FAM83F served as an oncogene and was connected with progression in several tumors. 25 –27 Also, miR-940 repressed cell viability in NSCLC cells through downregulation. 29 Furthermore, miR-1827 repressed cell invasion, colony formation, and proliferation, and induced apoptosis through downregulating FAM83F and MYC in lung adenocarcinoma. 28 Notably, in this study, FAM83F was modulated by circARHGAP10 by targeting miR-638. Besides, FAM83F augmentation overturned the repressive effects of miR-638 enhancement on cell proliferation, migration, invasion, and glycolysis in NSCLC cells. The results from this study manifested that circARHGAP10 played a role through the miR-638/FAM83F axis in NSCLC.
In sum, inhibition of serum-derived exosome-mediated circARHGAP10 repressed the progression of NSCLC through reducing FAM83F expression by targeting miR-638. This research might serve as a theoretical basis for developing effective NSCLC diagnostic biomarkers and therapeutic targets.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.