
Abstract
Background:
Circular RNAs (circRNAs) have recently emerged as crucial regulatory molecules in prostate cancer (PCa), but few researches focus on the effects of circRNAs on PCa radiosensitivity. The issue will be addressed in this study using circRNA Cyclin B2 (circ_CCNB2) as an object.
Materials and Methods:
All RNA and protein levels were severally examined using quantitative real-time polymerase chain reaction and Western blot. Colony formation assay and flow cytometry were implemented for detecting cell colony capacity and apoptotic cells, respectively. Cellular migration and invasion abilities were evaluated by transwell assay. The combination between potential target molecules was analyzed by dual-luciferase reporter and RNA immunoprecipitation (RIP) assays. The effect of circ_CCNB2 on PCa radiosensitivity in vivo was explored using xenograft models in mice.
Results:
Circ_CCNB2 was upregulated in irradiation-resistant PCa tissues and cells. Circ_CCNB2 knockdown had promoted effect on the radiosensitivity of irradiation-resistant PCa cells by inhibiting autophagy. Besides, circ_CCNB2 could directly sponge miR-30b-5p, and the promotion of circ_CCNB2 knockdown on PCa radiosensitivity was achieved by elevating miR-30b-5p. MiR-30b-5p enhanced the radiosensitivity of irradiation-resistant PCa cells through repressing the expression of its target kinesin family member 18A (KIF18A). Furthermore, circ_CCNB2 regulated the KIF18A level through targeting miR-30b-5p. Circ_CCNB2 downregulation facilitated PCa radiosensitivity in vivo through inhibiting autophagy by miR-30b-5p/KIF18A.
Conclusions:
In this study, knockdown of circ_CCNB2 was shown to promote PCa radiosensitivity through autophagy repression by miR-30b-5p/KIF18A axis, developing a molecular resistance mechanism of PCa radiotherapy and a feasible strategy to increase radiosensitivity.
Introduction
Prostate cancer (PCa) occupies the second place in the most prevalent cancers of males worldwide. 1 Radiation therapy exhibits exciting breakthroughs in the metastasis of PCa and postoperative repairing. 2 –4 There is a relapse in a large portion of PCa patients who received the radiotherapy, which is possibly ascribed to the generation of radioresistance. 5 Having a great understanding regarding the radioresistance mechanism may be beneficial to confront the recurrent PCa and block its metastasis. Currently, several molecular targets have been developed as radiosensitizer to regulate the radiosensitivity of PCa. 6
Circular RNAs (circRNAs) are classified as a subgroup of noncoding RNAs (ncRNAs) with the prototypical property of covalently closed-loop structures, which endow them with a high degree of stability. 7 Feng et al. demonstrated that circ0005276 enhanced the proliferation and migration abilities of PCa cells through XIAP activation by the interaction with FUS. 8 Huang et al. showed that circABCC4 improved the evolvement of PCa through the regulation of miR-1182/FOXP4 9 and Dai et al. reported that circ_0141940 targeted miR-29a to promote PCa progression. 10 Also, dysregulated circRNAs were showed to associate with drug enzalutamide resistance. 11 However, the regulation of circRNAs in radiosensitivity of PCa has not been reported. Circ_CCNB2 was proved to facilitate drug sensitivity of renal cancer, 12 hence the authors surmise whether circ_CCNB2 is also involved in the regulation of radiosensitivity in PCa.
MicroRNAs (miRNAs and short ncRNAs) can influence the radiosensitivity of PCa cells by targeting the 3′-untranslated regions (UTRs) of messenger RNA (mRNA) to mediate various cellular behaviors. 13 For example, miR-205 increased radiosensitivity of PCa through retarding DNA damage repair by inhibiting PKCɛ and ZEB1 14 and impairing autophagy through targeting TP53INP1. 15 The radiosensitivity of PCa was promoted by miR-875-5p acting EGFR-ZEB1 axis. 16 The low expression of miR-30b-5p was related to PCa metastasis in high-fat diet mice. 17 However, the effect of miR-30b-5p on radiation response of PCa remains unknown.
Kinesin family member 18A (KIF18A) can participate in the maturation and mitosis of spermatids. 18 Luo et al. asserted that the KIF18A enhanced the proliferation and metastasis of hepatocellular carcinoma cells. 19 Zhang et al. clarified the high expression of KIF18A in PCa and the inhibition of PCa progression caused by KIF18A knockdown. 20 During this study, the relationship among KIF18A, miR-30b-5p, and circ_CCNB2 in regulating radiation sensitivity of PCa will be a key point to grasp.
Materials and Methods
Patient samples
In total, 25 pairs of PCa and normal adjacent tissues were derived from PCa patients during the prostatectomy at the Second Affiliated Hospital of Fujian Medical University. These patients were categorized into the radiosensitive group (n = 13) and radioresistant group (n = 12) according to the following classification criteria: patients with local complete response (tumor disappearance) or partial response (PR, tumor shrinkage at least 30%) were termed as radiosensitive patients, and those patients with a progressive disease (PD, tumor increased at least 20%) or stable disease (tumor response between PR and PD) were defined as radioresistant patients after radiotherapy for 3 months. All these specimens were promptly conserved in liquid nitrogen for RNA extraction. Sample collection was performed after obtaining the informed consent from all patients and this research was authorized by the Ethics Committee of the Second Affiliated Hospital of Fujian Medical University.
Cell culture and X-ray irradiation treatment
Human PCa cell lines DU145 and LNCaP (COBIOER, Nanjing, China) were cultured in medium consisting of Roswell Park Memorial Institute-1640 (RPMI-1640), 10% fetal bovine serum (FBS), and 1% penicillin-streptomycin (Gibco, Carlsbad, CA), in a humidified incubator that controlled the temperature at 37°C and CO2 percentage of 5%. For establishing the irradiation-resistant cells (DU145/IR and LNCaP/IR), DU145 and LNCaP cells were exposed to X-ray irradiation by a linear accelerator (Faxitron Bioptics, LLC, San Francisco, CA) as reported previously. 21
Quantitative real-time polymerase chain reaction
In this study, RNA was acquired by Trizol total RNA extraction kit (KeyGen, Nanjing, China), then complementary DNA (cDNA) was synthesized by PrimeScript™ RT Master Mix (Takara, Beijing, China) with 2 μg RNA as template. Afterward, quantitative real-time polymerase chain reaction (qRT-PCR) was conducted through the TB Green® Fast qPCR Mix (Takara) using specific primers in Table 1. The data analysis was performed by 2−ΔΔCt approach 22 with GAPDH (for circ_CCNB2 and KIF18A) and U6 (for miR-30b-5p) as the endogenous controls.
Primers Used for Quantitative Real-Time Polymerase Chain Reaction Analysis
KIF18A, kinesin family member 18A.
Western blot
Radio Immunoprecipitation Assay (RIPA) lysis buffer (KeyGen) was used to extract total proteins, and protein quantification was carried out by the Bradford Protein Assay Kit (Takara). Subsequently, the mixture of proteins (40 μg) and loading buffer (Takara) was loaded on 10% sodium dodecyl sulfate–polyacrylamide gel (Invitrogen, Carlsbad, CA) to conduct electrophoresis for 2 h, followed by transferring proteins onto polyvinylidene fluoride membranes (Millipore, Billerica, MA) and blockage in Western blot Blocking Buffer (KeyGen) for respective 1 h. After that, the membranes were incubated with the diluent of primary antibodies anti-light chain 3B (anti-LC3B; ab51520, 1:1000), anti-Beclin1 (ab62557, 1:1000), anti-p62 (ab109012, 1:1000), anti-KIF18A (ab251863, 1:1000), anti-E-cadherin (ab15148, 1:1000), anti-Vimentin (ab71144, 1:1000), and internal reference anti-GAPDH (ab9485, 1:3000) for 4 h and anti-rabbit immunoglobulin G (IgG) H&L (HRP) secondary antibody (ab205718, 1:5000) for 1 h at 25°C. All antibodies in the authors' Western blot assay were bought from Abcam (Cambridge, United Kingdom). Finally, the immune conjugated signals were examined using ECL Western Blotting Substrate Kit (Abcam) and density analysis was implemented through the ImageLab software version 4.1 (Bio-Rad Laboratories, Hercules, CA). 23
Cell transfection
The authors used Lipofectamine3000 Transfection Reagent (Invitrogen) to fulfill cell transfection assay in terms of the producer's specification. The oligonucleotides (GenePharma, Shanghai, China) used in this study included small interfering RNA (siRNA) targeting circ_CCNB2 (si-circ_CCNB2), short hairpin RNA (shRNA) against circ_CCNB2 (sh-circ_CCNB2), miR-30b-5p mimic, miR-30b-5p inhibitor, and their negative controls (si-NC, sh-NC, mimic NC, and inhibitor NC). Specially, shRNA transfection was carried out through the pSINsi vector (Takara) to stably express sh-circ_CCNB2 or sh-NC. Besides, the overexpression vectors pCE-RB-Mam-circ_CCNB2 (Over-circ_CCNB2) and pcDNA3.1-KIF18A (Over-KIF18A) were constructed by cloning the sequences of KIF18A and circ_CCNB2 into the expression vectors pCE-RB-Mam (RIBIBIO, Guangzhou, China) and pcDNA3.1 (Invitrogen).
Colony formation assay
Cell cloning formation capability was evaluated using colony formation assay. In brief, 1 × 103 cells were seeded in six-well plates to grow in the normal culture condition for 2 weeks. After the colonies were fastened and dyed in 4% paraformaldehyde and crystal violet (Sigma, St. Louis, MO), the colonies were photographed and counted under a microscope.
Apoptosis assay
Annexin V-fluorescein isothiocyanate (Annexin V-FITC)/propidium iodide (PI) apoptosis detection kit (KeyGen) was employed for assessing cell apoptosis. Around 2 × 105 harvested cells were resuspended in 500 μL binding buffer and the obtained cell suspension was mixed with 5 μL Annexin V-FITC, followed by the addition of 5 μL PI for lucifugal incubation at room temperature. Ten minutes later, the apoptotic cells were observed under the flow cytometer (BD Biosciences, San Diego, CA), in which Annexin V+/PI− represented the apoptotic cells in the early phase and Annexin V+/PI+ for late phase. The formula: apoptotic cells/total cells × 100% was applied for the calculation of the apoptosis rate.
Migration and invasion assays
For migration detection, 4 × 103 cells in the serum-free medium were seeded into the upper chamber of the transwell 24-well chamber (Corning, Inc., Corning, NY), and the lower chamber was filled with medium with 10% FBS to attract cells. After 24 h, a sterile cotton swab was used for removing the unmigrated cells on the upper side of the membrane, while migrated cells on the reverse side of the membrane were fixated in 4% paraformaldehyde (Sigma) and stained using crystal violet (Sigma). Ultimately, the picture and data of migrated cells were obtained through a microscope. For invasion detection, the lower surface of the transwell upper chamber needed to be coated with matrigel (Corning, Inc.) first and the other operation steps were performed as migration detection above.
Dual-luciferase reporter assay
The amplified sequences of circ_CCNB2 and KIF18A 3′UTR, including wild type (WT) with the binding domain of miR-30b-5p and mutant type (MUT) with the mutated binding domain of miR-30b-5p, were inserted into the pGL3 vector (Promega, Madison, WI). After acquiring the positive recombinational luciferase plasmids (circ_CCNB2 WT/MUT and KIF18A 3′UTR-WT/MUT), 293T cells (COBIOER) were transfected with above respective plasmids and miR-30b-5p mimic or mimic NC to cultivate in DMEM complemented with 10% FBS and 1% penicillin-streptomycin (Gibco) for 48 h. Then the dual-luciferase reporter system (Promega) was used for examining the firefly and Renilla luciferase activities of cell lysates conforming to the operation manual offered by the producer. The normalization of firefly luciferase intensity was achieved by Renilla luciferase activity.
RNA immunoprecipitation assay
Magna RNA immunoprecipitation (RIP) kit (Millipore) was conducted to validate whether circ_CCNB2 could combine with miR-328-3p. Fifty microliters of cell lysate in RIP lysis buffer was extracted as the positive control Input. Whereafter, magnetic beads precovered with anti-Argonaute2 (anti-Ago2;ab32381; Abcam) were used for incubating other 50 μL cell lysate with anti-Immunoglobulin G (anti-IgG; ab205718; Abcam) as the negative control. Then circ_CCNB2 and miR-30b-5p expression levels were measured using qRT-PCR following the isolation of RNA from magnetic beads.
Xenograft tumor assay
BALB/c nude mice (7-week-old, n = 12) purchased from Vital River Laboratory Animal Technology (Beijing, China) were divided into two groups (sh-NC and sh-circ_CCNB2, six mice/group) at random. Then 500 μL cell suspension (about 2 × 105 cells) of DU145 cells that expressed sh-NC or sh-circ_CCNB2 was subcutaneously injected into the unilateral back of mice to establish the xenograft model. Twelve days later, mice accepted irradiation treatment with 10 Gy daily. A digital caliper was applied for monitoring the tumor size and tumor volume was recorded every 5 d according to the computational formula: length × width 2 /2. Thirty-seven days later, all mice were euthanized by CO2 asphyxia method in accordance with the guideline of American Veterinary Medical Association (AVMA), and then tumor weight was measured using an electronic scale after the tumor excision from mice. Then the expression of circ_CCNB2, miR-30b-5p, and KIF18A as well as the protein levels of autophagy-related indexes (LC3II/LC3I, Beclin1, and p62) were analyzed by qRT-PCR and Western blot. This animal experiment was approved by the Animal Ethics Committee of the Second Affiliated Hospital of Fujian Medical University.
Statistical analysis
Statistical analyses in this study were implemented using SPSS 22.0 with three independent repetitions of all assays and the mean ± standard deviation of data exhibition. And graph drawing was performed by GraphPad Prism 7. The liner connection was analyzed by Spearman's correlation coefficient. Student's t-test and one-way analysis of variance followed by Tukey's test were applied for the difference analysis. p < 0.05 indicated that the statistical difference was significant.
Results
Circ_CCNB2 was overexpressed in irradiation-resistant PCa tissues and cells
After the qRT-PCR analysis in 25 PCa tissue samples, the authors found the upregulation of circ_CCNB2 in contrast to normal adjacent tissues (Fig. 1A). And among 25 samples, 13 irradiation-resistant tissues displayed the increased trend of circ_CCNB2 expression compared with 12 irradiation-sensitive tissues (Fig. 1B). In addition, the circ_CCNB2 level in DU145/IR and LNCaP/IR cells was much higher than that in DU145 and LNCaP cells (Fig. 1C). By comparison to sensitive tissues and cells, the protein level of p62 (an antiautophagy marker) was inhibited, while Beclin1 and LC3II/LC3I (pro-autophagy indexes) were enhanced in irradiation-resistant PCa tissues (Fig. 1D), as well as in DU145/IR and LNCaP/IR cells (Fig. 1E). The overexpression of circ_CCNB2 might be associated with autophagy in irradiation-resistant PCa.
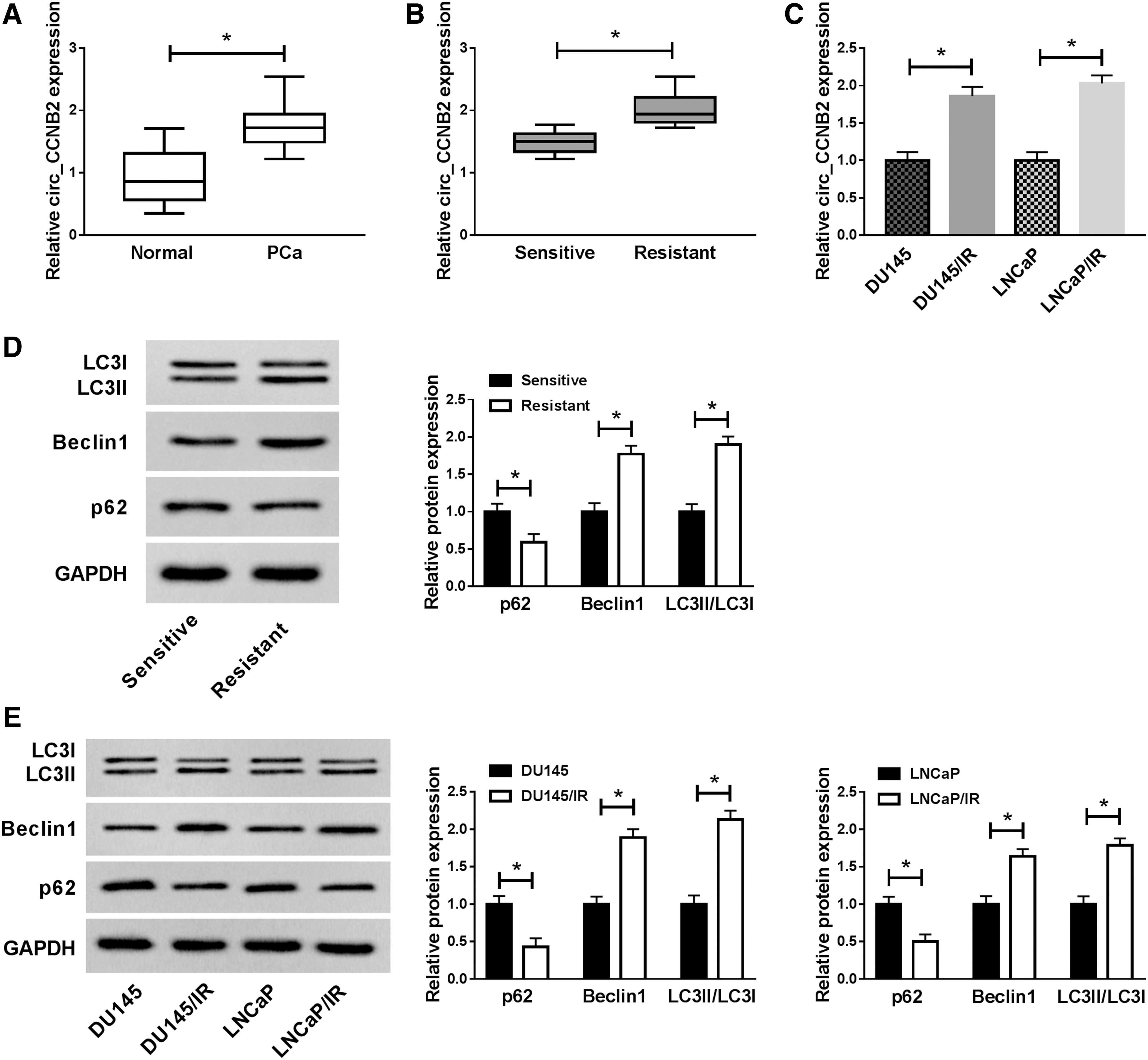
Circ_CCNB2 was overexpressed in irradiation-resistant PCa tissues and cells.
Knockdown of circ_CCNB2 enhanced the radiosensitivity by inhibiting autophagy in irradiation-resistant PCa cells
To further investigate the relationship between circ_CCNB2 and autophagy in regulating radiosensitivity of PCa, siRNA targeting circ_CCNB2 was transfected into DU145/IR and LNCaP/IR cells. The qRT-PCR indicated that si-circ_CCNB2 transfection successfully interfered the circ_CCNB2 expression relative to si-NC transfection (Fig. 2A). Colony formation assay showed that colonies of si-circ_CCNB2 group were repressed compared with si-NC group (Fig. 2B). Also, the authors found that the introduction of si-circ_CCNB2 generated a stimulative effect on cell apoptosis (Fig. 2C) contrasted to si-NC group in DU145/IR and LNCaP/IR cells. Transwell assay manifested that the number of migrated and invaded cells was decreased, as the results of circ_CCNB2 knockdown (Fig. 2D). These data explained that circ_CCNB2 knockdown promoted the radiosensitivity of PCa cells to inhibit PCa progression. Subsequently, they observed that p62 was increased, while Beclin1 and LC3II/LC3I were declined after circ_CCNB2 was knocked down, exhibiting that si-circ_CCNB2 evoked the autophagy inhibition in DU145/IR and LNCaP/IR cells (Fig. 2E). Furthermore, the authors used an autophagy inhibitor 3-MA (Sigma) to analyze the relationship between autophagy and circ_CCNB2. The results demonstrated that 3-MA rescued the circ_CCNB2 overexpression-mediated radiosensitivity inhibition in irradiation-resistant PCa cells as the illustration of Supplementary Figure S1, hinting that circ_CCNB2 effect is actually mediated by autophagy promotion. Therefore, they concluded that knockdown of circ_CCNB2 impaired autophagy to enhance radiosensitivity of irradiation-resistant PCa cells.
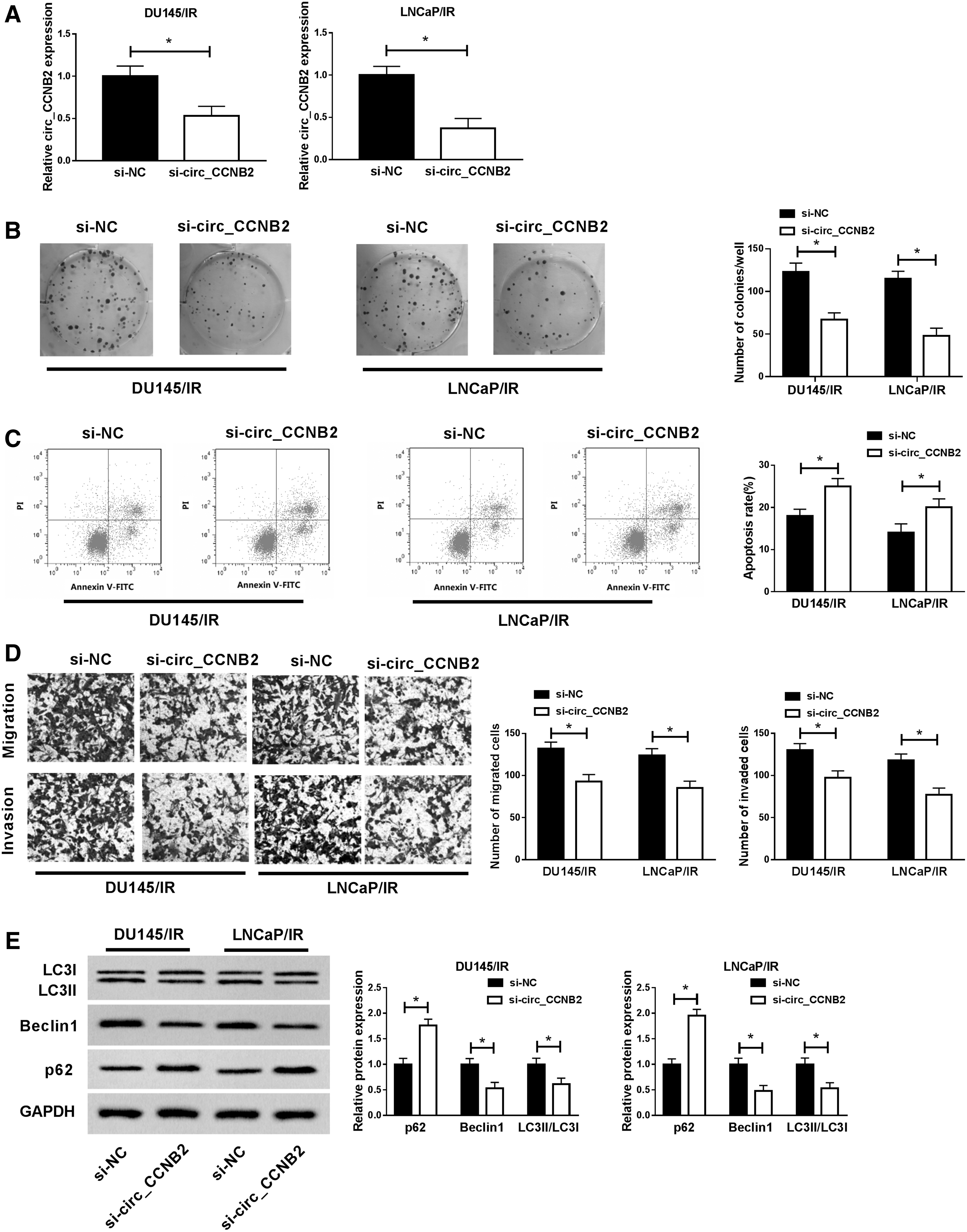
Knockdown of circ_CCNB2 enhanced radiosensitivity by inhibiting autophagy in irradiation-resistant PCa cells. Respective transfection of si-circ_CCNB2 and si-NC was performed for 48 h in DU145/IR and LNCaP/IR cells.
MiR-30b-5p was a target of circ_CCNB2
Researchers have identified that circRNAs can target miRNAs. 24,25 The authors noticed that miR-30b-5p expression was lower in PCa tissues than that in normal adjacent tissues and this downregulation phenomenon was also presented in irradiation-resistant PCa tissues and cells (DU145/IR and LNCaP/IR) by comparison with irradiation-sensitive tissues and cells (DU145 and LNCaP) (Fig. 3A, B). And circ_CCNB2 expression was negatively correlated with miR-30b-5p (r = −0.4759, p = 0.0162) in PCa tissues (Fig. 3C). The bioinformatics analysis of Starbase 3.0 exhibited the binding region between circ_CCNB2 and miR-30b-5p, then dual-luciferase reporter assay made clear that miR-30b-5p overexpression resulted in the inhibition of luciferase activity of circ_CCNB2 WT plasmid in 293T cells in place of circ_CCNB2 MUT plasmid (Fig. 3D). Moreover, the enrichment of circ_CCNB2 and miR-30b-5p in anti-Ago2 group was much higher than that in anti-IgG group, close to the positive control Input, insinuating the combination between circ_CCNB2 and miR-30b-5p (Fig. 3E). Altogether, miR-30b-5p was a target of circ_CCNB2.

MiR-30b-5p was a target of circ_CCNB2.
Inhibition of miR-30b-5p reversed the effects of si-circ_CCNB2 on irradiation-resistant PCa cells
In DU145/IR and LNCaP/IR cells transfected with si-circ_CCNB2, miR-30b-5p level was signally enhanced, making a comparison with the si-NC group (Fig. 4A). Due to the peculiarity of circ_CCNB2 as a miR-30b-5p sponge, the rescue experiment was used to analyze whether miR-30b-5p was related to the function of circ_CCNB2 on PCa radiosensitivity. Also, miR-30b-5p inhibition reverted the si-circ_CCNB2-induced suppressive effect on colony capacity (Fig. 4B) and promotive effect on apoptosis (Fig. 4C). As a consequence of the introduction of miR-30b-5p inhibitor, the migration and invasion inhibition caused by si-circ_CCNB2 were also alleviated (Fig. 4D, E). Furthermore, the rising of p62 protein expression and the lessening of Beclin1, LC3II/LC3I levels caused by si-circ_CCNB2 were all restored after the presence of miR-30b-5p inhibitor (Fig. 4F). Above data indicated that miR-30b-5p inhibitor could recover the si-circ_CCNB2-induced effects on irradiation-resistant PCa cells.
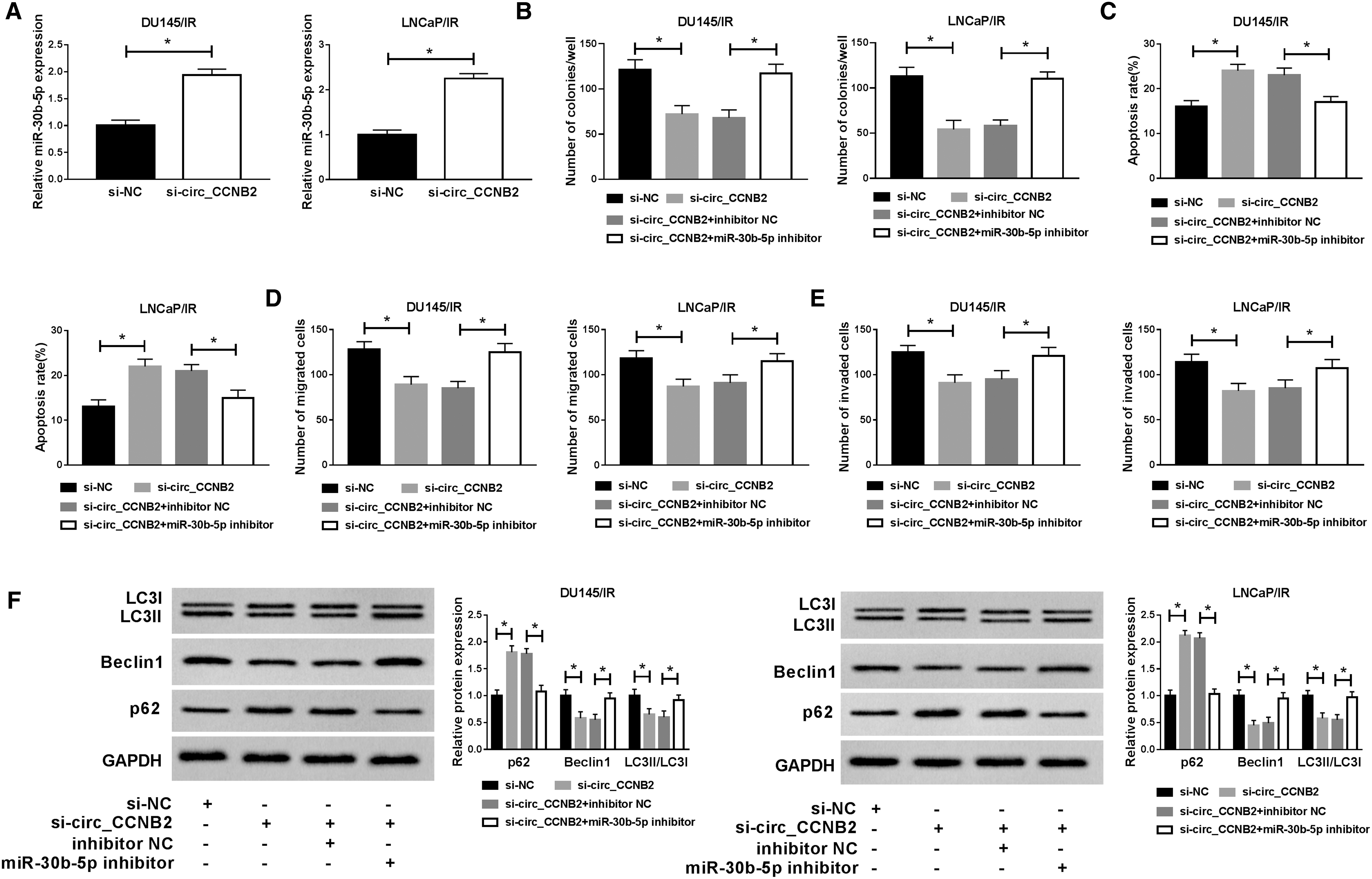
Inhibition of miR-30b-5p reversed the effects of si-circ_CCNB2 on irradiation-resistant PCa cells.
MiR-30b-5p directly targeted KIF18A
Through the analysis of qRT-PCR and Western blot, the KIF18A mRNA (Fig. 5A) and protein (Fig. 5B) expression levels were found to be upregulated in PCa tissues and irradiation-resistant PCa tissues, compared to the normal adjacent tissues and irradiation-sensitive PCa tissues. Additionally, this ascending tendency of KIF18A mRNA and protein expression was displayed in DU145/IR and LNCaP/IR cells with DU145 and LNCaP cells as controls (Fig. 5C, D). 3′UTR of mRNA is recognized as an acting factor of miRNA. 26 The analysis result of Spearman's correlation coefficient showed a negative relationship (r = −0.5975 and p = 0.0016) between miR-30b-5p and KIF18A in PCa tissues (Fig. 5E), implying that miR-30b-5p might target KIF18A. As expected, miR-30b-5p had the target combinative sites in the 3′UTR of KIF18A in Starbase 3.0 and dual-luciferase reporter assay confirmed their binding in 293T cells (Fig. 5F). Thus, KIF18A was a target of miR-30b-5p.
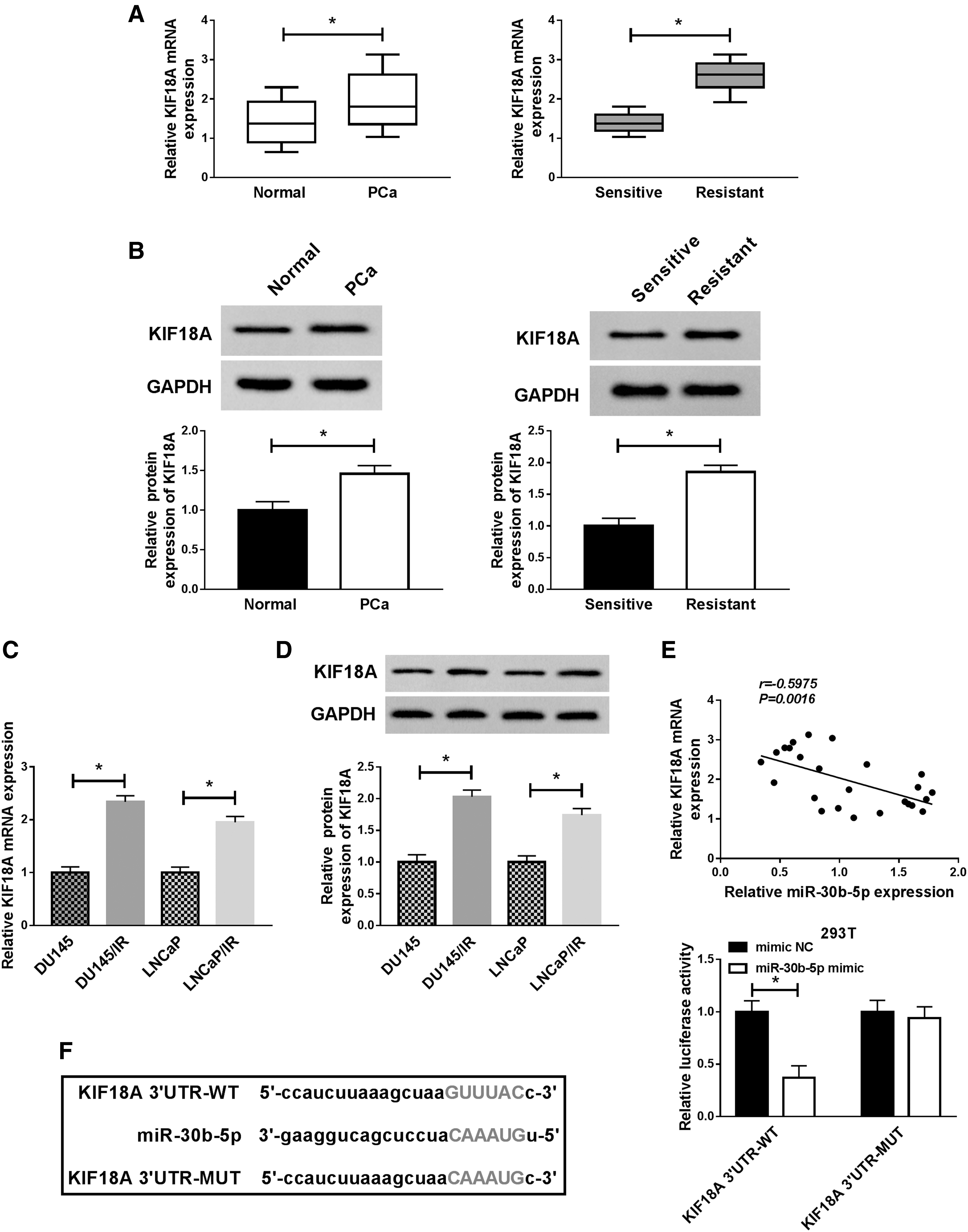
MiR-30b-5p directly targeted KIF18A.
MiR-30b-5p promoted the radiosensitivity of irradiation-resistant PCa cells through repressing autophagy by inhibiting KIF18A
The effect of miR-30b-5p on KIF18A expression was researched. As Figure 6A and B described, miR-30b-5p mimic transfection caused the downregulation of KIF18A mRNA and protein levels in DU145/IR and LNCaP/IR cells by contrast with the mimic NC group. Furthermore, the authors discovered that cell colony capacity was refrained (Fig. 6C) and the apoptosis rate was heightened (Fig. 6D) after transfection of miR-30-5p mimic, along with the inhibition of cell migration and invasion (Fig. 6E, F), but it is interesting that all these effects were offset by the overexpression of KIF18A. Besides, the elevation of miR-30b-5p expression triggered the autophagy suppression judging from the upregulated p62 and decreased Beclin1 or LC3II/LC3I, whereas abnormal high expression of KIF18A neutralized these changes (Fig. 6G). All in all, miR-30b-5p targeted KIF18A to facilitate the radiosensitivity of irradiation-resistant PCa cells through hindering autophagy.
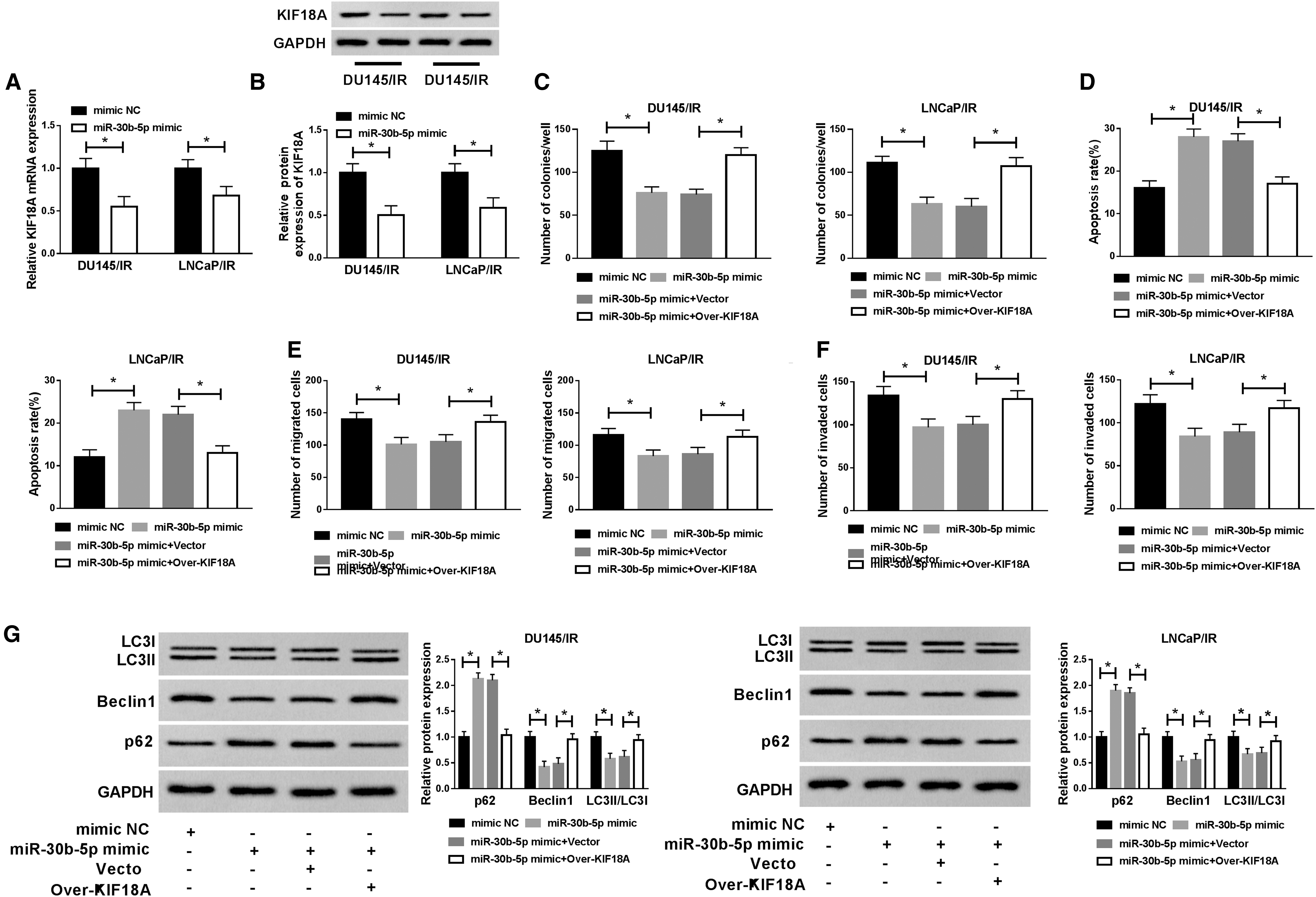
MiR-30b-5p promoted the radiosensitivity of irradiation-resistant PCa cells through repressing autophagy by targeting KIF18A.
Circ_CCNB2 positively regulated the expression of KIF18A by targeting miR-30b-5p
Due to the target relationship between circ_CCNB2 and miR-30b-5p, as well as miR-30b-5p and KIF18A, the authors designed a rescue experiment to analyze the relationship between circ-CCNB2 and KIF18A. As the illustration of Figure 7A, KIF18A mRNA level was overtly reduced after circ_CCNB2 knockdown, while co-transfection of si-circ_CCNB2 and miR-30b-5p inhibitor relived this inhibition contrasted with si-circ_CCNB2+inhibitor NC group. Similarly, miR-30b-5p inhibitor also ameliorated the si-circ_CCNB2-induced inhibitory impact on KIF18A protein level (Fig. 7B). Hence, the authors draw a conclusion that circ_CCNB2 could positively modulate the KIF18A expression by targeting miR-30b-5p.

Circ_CCNB2 positively regulated the expression of KIF18A by targeting miR-30b-5p.
Circ_CCNB2 knockdown contributed to the radiosensitivity of PCa in vivo through autophagy impairment by the miR-30b-5p/KIF18A axis
Xenograft tumor assay was performed for studying the function of circ_CCNB2 in PCa in vivo. As Figure 8A presented, tumor volume of sh-circ_CCNB2 group was notably decreased after X-ray irradiation 12 d postinjection, contrasted to sh-NC group. And the tumor weight was also inhibited after circ_CCNB2 downregulation in vivo (Fig. 8B). In excised tumors, circ_CCNB2 and KIF18A (mRNA and protein) levels were clearly diminished, while miR-30b-5p was boosted after the knockdown of circ_CCNB2 (Fig. 8C, D). In addition, Western blot indicated that E-cadherin (antimetastasis marker) was upregulated, while Vimentin (prometastasis marker) was deceased after circ_CCNB2 was downregulated in mice, demonstrating that circ_CCNB2 knockdown also inhibited PCa metastasis in vivo (Fig. 8E). Simultaneously, the authors found that p62 protein expression was increased but Beclin1 and LC3II/LC3I had the inverse trend in tumors of sh-circ_CCNB2 group in contrast with sh-NC group (Fig. 8E). Collectively, circ_CCNB2 knockdown expedited the radiosensitivity of PCa in vivo through the impairment of autophagy by the miR-30b-5p/KIF18A axis.
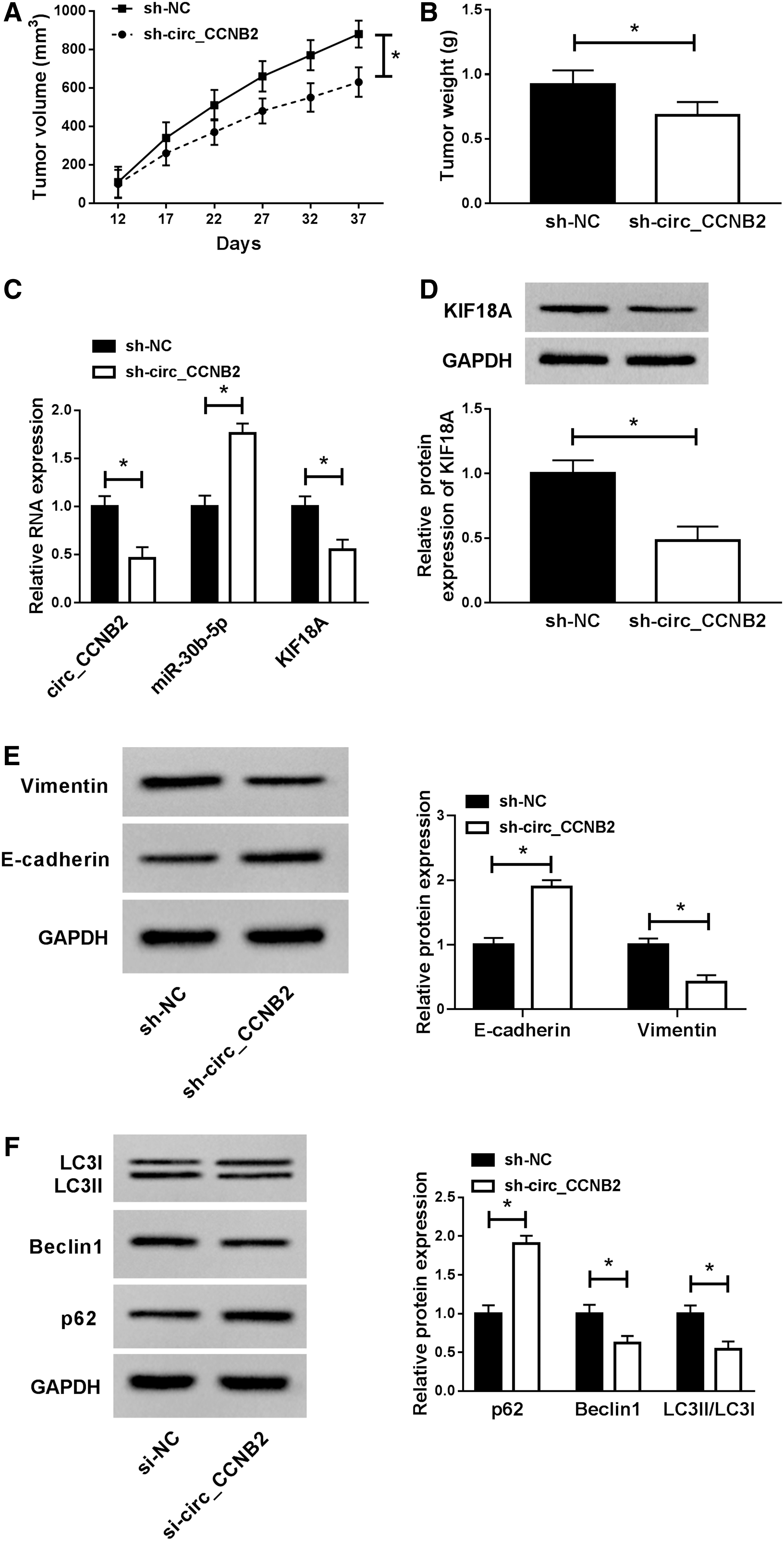
Circ_CCNB2 knockdown contributed to the radiosensitivity of PCa in vivo through autophagy impairment by the miR-30b-5p/KIF18A axis.
Discussion
The appearance of numerous molecules contributes to withstand the radioresistance generation of PCa. 27,28 This study suggested that circ_CCNB2 knockdown could heighten the radiosensitivity of PCa through autophagy regulation by acting on the miR-30b-5p/KIF18A axis, which proved that circ_CCNB2 could regard as a molecular indicator of radioresistance in PCa radiation treatment.
As regard the regulation of circRNAs in radiosensitivity of human cancers, Chen et al. announced that circ_000543 knockdown reinforced the radiation sensitivity of nasopharyngeal carcinoma through regulating PDGFRB expression by targeting miR-9 29 ; Yang et al. also showed that the inhibition of circRNA ZNF292 exerted the repressive effect on radioresistance of hypoxic hepatoma cells. 30 Also, the influence of circATP8B4 on improving radioresistance has been in the elucidation of glioma. 31 The function research of these circRNAs is started from their aberrant upregulation. Herein, the authors first discovered the overexpression of circ_CCNB2 in irradiation-resistant PCa tissues and cells. After circ_CCNB2 was knocked down, colony formation and metastasis abilities of irradiation-resistant PCa cells were inhibited, while apoptosis was promoted, implicating that circ_CCNB2 knockdown enhanced the radiosensitivity of PCa cells to retard the tumor progression.
Autophagy is a cellular process regulating cell signaling and maintaining homeostasis, and the regulation of autophagy is closely related to the radiosensitivity of cancer cells. 3,32 For instance, chloroquine (an autophagy repressor) inhibited autophagy to increase the radiosensitivity of bladder cancer cells 33 and blocking autophagy made osteosarcoma cells more sensitive to radiation in vitro. 34 Liu et al. found that curcumin motivated the radiosensitivity of PCa cells by the suppression of miR-143/miR-143-mediated autophagy. 35 Currently, the authors found the promotion of autophagy after the related protein detection in irradiation-resistant PCa tissues and cells, indicating that autophagy might induce the radioresistance formation of PCa. Then they found autophagy inhibitor 3-MA returned the tumor-promotive effect of circ_CCNB2 overexpression on irradiation-resistant PCa cells, which gave the evidence that the role of circ_CCNB2 in regulating radiosensitivity of PCa was attributable to autophagy mediation, at least in part.
In the molecular level, the functional mechanisms of circRNAs are usually dependent upon the circRNA-miRNA-mRNA network in different kinds of cancers, including PCa. 36 –38 For circRNA-miRNA, the authors identified miR-30b-5p as a miRNA target for circ_CCNB2. MiR-30b-5p was underexpressed in PCa tissues and cells with radiation resistance. And the effect of circ_CCNB2 on regulating radiosensitivity was achieved by targeting miR-30b-5p. For miRNA-mRNA, KIF18A was a downstream target of miR-30b-5p. KIF18A is an oncogene to facilitate cell growth and metastasis in lung adenocarcinoma. 39 –41 Herein, KIF18A could eliminate the repressive effect of miR-30b-5p on cell growth, metastasis, and autophagy, which suggested that KIF18A also played a tumor-promoting role in irradiation-resistant PCa cells. Moreover, miR-30b-5p targeted KIF18A to promote the radiosensitivity of PCa by blocking autophagy. Wang et al. exhibited that the modulation of circ_014511 on radiosensitivity in bone marrow mesenchymal stem cells was owed to the miR-29b-2-5p/P53 axis. 42 Conformably, circ_CCNB2 could bind to miR-30b-5p to affect the expression of KIF18A, exerting the regulation of radiation sensitivity of PCa in this report. Furthermore, the authors' experiments in vivo notarized that circ_CCNB2 downregulation could restore the radiosensitivity of PCa through the miR-30b-5p/KIF18A-mediated autophagy impairment.
Conclusions
Altogether, the authors' results showed that knockdown of circ_CCNB2 enhanced the PCa radiosensitivity through autophagy obstruction by acting on the miR-30b-5p/KIF18A axis. The modulatory mechanism of circ_CCNB2/miR-30b-5p/KIF18A concerning the sensitivity of PCa to radiation was disclosed in public. Circ_CCNB2 has the potential to predict the generation of radioresistance in PCa radiotherapy. In the therapeutic level, the suppression of circ_CCNB2 may have radiation sensitization for improving the therapeutic effect of radiotherapy on those recurrent PCa patients.
Footnotes
Availability of Data and Materials
The analyzed data sets generated during this study are available from the corresponding author on reasonable request.
Ethics Approval and Consent to Participate
This study was approved by the ethical review committee of the Second Affiliated Hospital of Fujian Medical University.
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.