
Abstract
Background:
Non-small cell lung cancer (NSCLC) is the most prevalent cancer in the world. Chemotherapy resistance is a major obstacle to NSCLC therapy. This study explored the role and molecular mechanism of circular RNA 0011292 (circ_0011292) in tumorigenesis and chemoresistance of NSCLC.
Methods:
The levels of circ_0011292, miR-379-5p, and tripartite motif-containing protein 65 (TRIM65) were measured by quantitative real-time polymerase chain reaction or Western blot assay. Cell proliferation was assessed by Cell Counting Kit-8 (CCK-8) assay. Cell apoptosis was monitored by flow cytometry. Cell migration and invasion were detected by transwell assay. The levels of apoptosis-related and epithelial-mesenchymal transition-related proteins were examined by Western blot. The half-inhibition concentration (IC50) of paclitaxel (PTX) was evaluated by CCK-8 assay. Xenograft model was established to analyze the effect of circ_0011292 on PTX resistance of NSCLC in vivo. The interaction among circ_0011292, miR-379-5p, and TRIM65 was verified by dual-luciferase reporter assay and RNA immunoprecipitation assay.
Results:
Circ_0011292 and TRIM65 were upregulated, while miR-379-5p was downregulated in NSCLC tissues and cells. Circ_0011292 knockdown hindered NSCLC progression and enhanced PTX sensitivity of NSCLC. Circ_0011292 silencing reduced PTX resistance in vivo. Besides, miR-379-5p potentiated PTX sensitivity by targeting TRIM65. Also, circ_0011292 increased PTX resistance by sponging miR-379-5p.
Conclusion:
Circ_0011292 facilitated tumorigenesis and PTX resistance in NSCLC by regulating the miR-379-5p/TRIM65 axis, suggesting that circ_0011292 was a promising therapeutic target for NSCLC chemotherapy.
Introduction
Lung cancer ranks first among cancer deaths in the world, accounting for ∼18.4%. 1 About 85% of lung carcinomas are identified as non-small cell lung cancer (NSCLC), of which are mainly adenocarcinoma and squamous cell carcinoma. 2 Significant advances have been made in NSCLC treatment, such as the use of small molecule tyrosine kinase inhibitors and immunotherapy. 3 However, the high metastasis and recurrence rates of NSCLC result in a 5-year survival rate as low as 16%. 4 Besides, chemotherapy resistance is also an essential factor leading to poor prognosis in NSCLC patients. 5 Therefore, finding the molecular mechanism of chemotherapy resistance or sensitivity is imperative for improving the prognosis of NSCLC.
Circular RNAs (circRNAs) are noncoding RNAs with a covalent closed-loop structure, which are different from linear RNAs with 5′ caps and 3′ tails. 6 Increasing evidence suggested that circRNAs are strongly associated with the pathogenesis of various cancers. 7 For example, circCLK3 sponged microRNA (miRNA)-320a to accelerate cervical cancer tumorigenesis by upregulating FoxM1. 8 Circ_0001955 contributed to the progression of hepatocellular carcinoma by serving as a sponge of miRNA-516a-5p. 9 CircPTPRA suppressed bladder cancer growth by regulating the miRNA-636/KLF9 axis. 10 In addition, studies have indicated that circRNAs exert a vital regulatory effect on the epithelial-mesenchymal transition (EMT) of NSCLC. 11 Previous research showed that circ_0011292 derived from tubulointerstitial nephritis antigen like 1 (TINAGL1) was upregulated in Taxol-resistant A549 cells. 12 Nevertheless, the role and mechanism of circ_0011292 in NSCLC remain unknown.
miRNAs are short noncoding RNAs with a length of 18–24 nucleotides. Accumulating evidence verified that the aberrant expression of miRNAs was closely related to tumor occurrence and progression. 13 Furthermore, multiple miRNAs are promising biomarkers for the diagnosis and treatment of NSCLC. 14 Also, circRNAs can function as a molecular sponge for miRNAs to influence tumor biogenesis. 15 However, the sponge effect of circ_0011292 has not been studied.
Tripartite motif-containing protein 65 (TRIM65) is considered a potential cancer-promoting factor. 16 In addition, TRIM65 was first identified to be associated with white matter lesions. 17 Wei et al. found that TRIM65 strengthened the invasiveness of bladder urothelial carcinoma cells by activating the ubiquitination of ANXA2 and inducing EMT. 18 TRIM65 expression was prominently increased in NSCLC patients, and TRIM65 knockdown impeded NSCLC growth by activating tumor suppressor p53. 19
Hence, the authors first validated that circ_0011292 expression was overtly elevated in NSCLC tissues and cells. Next, they investigated the role and potential mechanism of circ_0011292 in the development and paclitaxel sensitivity of NSCLC.
Materials and Methods
Clinical specimens
Forty-three NSCLC tissues and adjacent normal tissues were obtained from NSCLC patients undergoing tumor resection at Weifang People's Hospital. This research was authorized by the Ethics Committee of Weifang People's Hospital. All participants were informed of the research purpose and signed written informed consent.
Cell culture
The fetal lung fibroblast cell line MRC5 and NSCLC cell lines (A549 and H1299) were bought from the American Type Culture Collection (ATCC, Manassas, VA). All cells were incubated in RPMI-1640 medium supplemented with 10% fetal bovine serum (Gibco, Carlsbad, CA) at 37°C with 5% CO2. A549 and H1299 cells were treated with increasing doses of paclitaxel (PTX) (Solarbio, Beijing, China) to establish PTX-resistant NSCLC cell lines (A549/PTX and H1299/PTX). Then, PTX-resistant cells were exposed to 0.5 mM PTX to maintain a resistant phenotype.
Cell transfection
Small interfering RNA (siRNA) targeting circ_0011292 (si-circ_0011292#1 and si-circ_0011292#2), the siRNA control (si-NC), miR-379-5p mimics (miR-379-5p), the mimics control (miR-NC), circ_0011292 overexpression vector (circ_0011292), TRIM65 overexpression vector (TRIM65), the empty overexpression vector (pcDNA), miR-379-5p inhibitor (anti-miR-379-5p), and the control (anti-miR-NC) were commercially obtained from GenePharma (Shanghai, China). Lipofectamine 3000 (Invitrogen, Carlsbad, CA) was utilized to transfect oligonucleotides and vectors.
Quantitative real-time polymerase chain reaction
After extracting RNA using Trizol reagent (Invitrogen), complementary DNA (cDNA) was synthesized using M-MLV RT Kit (AiYou Biosciences, Guangzhou, China) or miRNA 1st Strand cDNA Synthesis Kit (Vazyme, Nanjing, China). Then, quantitative real-time polymerase chain reaction (qRT-PCR) was performed by AceQ qPCR SYBR Green Master Mix (Vazyme). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and U6 were utilized as internal controls. For RNase R treatment, 2 μg RNA was incubated with or without RNase R (3U/μg) (Sigma, St. Louis, MO) for 30 min at 37°C. The primers were as follows: circ_0011292-F: 5′-ACATCTACCAGGTCACTCCTGT-3′, circ_0011292-R: 5′-ATGGTTCTTGGTCACACTGCCA-3′; TINAGL1-F: 5′-ACCAGGTCACTCCTGTCTACC-3′, TINAGL1-R: 5′-TGCCTCCCTTGTATAGGAAGAA-3′; miR-379-5p-F: 5′-GCGCTGGTAGACTATGGAA-3′, miR-379-5p-R: 5′-GTGCAGGGTCCGAGGT-3′; TRIM65-F: 5′-CGGAATTCATGGCCGCGCAGCTG-3′, TRIM65-R: 5′-CGGGATCCTCAGCTGAGCACCTCTTCC-3′; GAPDH-F: 5′-ACAACTTTGGTATCGTGGAAGG-3′, GAPDH-R: 5′-GCCATCACGCCACAGTTTC-3′; and U6-F: 5′-CTCGCTTCGGCAGCACA-3′, U6-R: 5′-AACGCTTCACGAATTTGCGT-3′.
Cell Counting Kit-8 assay
The treated cells (3 × 103) were plated into 96-well plates and cultured for 24, 48, 72, or 96 h, respectively. In addition, cells (3 × 103) seeded into 96-well plates were treated with increasing doses of PTX (0.05–12.8 μM). Next, cells were interacted with 10 μL Cell Counting Kit-8 (CCK-8) solution (Solarbio) for 4 h. Finally, a Multimode Reader (BioTek, Burlington, VT) was used to assess cell viability by measuring the optical density at 450 nm. PTX resistance was expressed as the half inhibition concentration (IC50) of PTX.
Flow cytometry
The treated cells were seeded into six-well plates. Then, AnnexinV-fluorescein isothiocyanate (AnnexinV-FITC)/Propidium Iodide (PI) Apoptosis Detection kit (Invitrogen) was used to detect cell apoptosis. Subsequently, the apoptosis rate was monitored by BD FACSCalibur flow cytometer (BD Biosciences, Franklin Lakes, NJ).
Transwell assay
In cell migration assay, cells were placed in the upper chamber. After 24 h of culture, the upper cells were swabbed with a cotton swab, and the lower migrated cells were stained with crystal violet. Then, five fields were randomly selected under the microscope to count the migrated cells. In cell invasion assay, the procedures were the same as the migration assay, except that transwell chambers were coated with Matrigel (BD Biosciences).
Western blot assay
After lysing cells with RIPA buffer (Solarbio), the extracted protein was separated by polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA). Then, the membranes were incubated with primary antibodies against B-cell lymphoma-2 (Bcl-2) (ab196495; Abcam, Cambridge, UK), cleaved-caspase 3 (c-caspase 3) (ab2302; Abcam), caspase 3 (ab4051; Abcam), E-cadherin (ab15148; Abcam), N-cadherin (ab76057; Abcam), vimentin (ab137321; Abcam), proliferating cell nuclear antigen (PCNA) (ab18197, Abcam), TRIM65 (ab165884; Abcam), or GAPDH (ab9485; Abcam) after blocking with 5% skim milk. Subsequently, the membranes were interacted with secondary antibody (ab7090; Abcam). The signal intensity was measured by the enhanced chemiluminescence system (Millipore).
Xenograft experiment
Lentiviral vectors carrying circ_0011292 short hairpin RNA (sh-circ_0011292) or negative control (sh-NC) were synthesized from Genechem (Shanghai, China). BALB/c nude mice (5-week old) were randomly divided into two groups (n = 6 per group). A549/PTX cells (5 × 106) transfected with sh-circ_0011292 or sh-NC were subcutaneously injected into the right back of nude mice. Subsequently, the mice were given phosphate buffered saline (PBS) or 3 mg/kg PTX (Solarbio) every 3 days after 6 days of injection. Tumor volume was monitored every 3 days. After 24 days of inoculation, the mice were killed and the xenografts were weighed. The animal experiment was approved by the Animal Research Committee of Weifang People's Hospital.
Dual-luciferase reporter assay
A549/PTX and H1299/PTX cells were plated in 24-well plates. The sequences of circ_0011292 or TRIM65 3′UTR containing wild-type or mutant binding sites of miR-379-5p were inserted into pmirGLO vector (Promega, Madison, WI), named as circ_0011292-wt, circ_0011292-mut, TRIM65 3′UTR-wt, and TRIM65 3′UTR-mut reporter. The corresponding luciferase reporter and miR-379-5p mimics or miR-NC were co-transfected into A549/PTX and H1299/PTX cells. Finally, the luciferase intensity was tested by Dual-Lucy Assay Kit (Solarbio).
RNA immunoprecipitation (RIP) assay
RIP assay was carried out using Magna RIP kit (Millipore). First of all, A549/PTX and H1299/PTX cells were lysed by RIP lysis buffer. Then, cells were incubated with magnetic beads conjugated with Ago2 antibody or IgG antibody. Finally, the levels of co-precipitated RNA were measured by qRT-PCR.
Statistical analysis
All data were shown as mean ± standard deviation using GraphPad Prism 7.0 software (GraphPad, San Diego, CA). Student's t-test was utilized to compare differences between two groups, and one-way analysis of variance was used to detect the differences among multiple groups. p < 0.05 was considered statistically significant.
Results
Circ_0011292 was upregulated in NSCLC tissues and PTX-resistant cells
To study the role of circ_0011292 in NSCLC, the authors first examined the expression difference of circ_0011292 in NSCLC tissues and adjacent normal tissues. The qRT-PCR results exhibited that circ_0011292 level in NSCLC tissues was remarkably higher than that in matched normal tissues (Fig. 1A). Next, they further tested the expression of circ_0011292 in parental cells and PTX-resistant cells. The results suggested that circ_0011292 level in A549 and H1299 cells was markedly higher than that in MRC5 cells and strikingly lower than that in A549/PTX and H1299/PTX cells (Fig. 1B). Besides, qRT-PCR revealed that circ_0011292 was resistant to RNase R relative to its linear isoform TINAGL1 (Fig. 1C, D). These data indicated that circ_0011292 might play a vital role in PTX resistance.
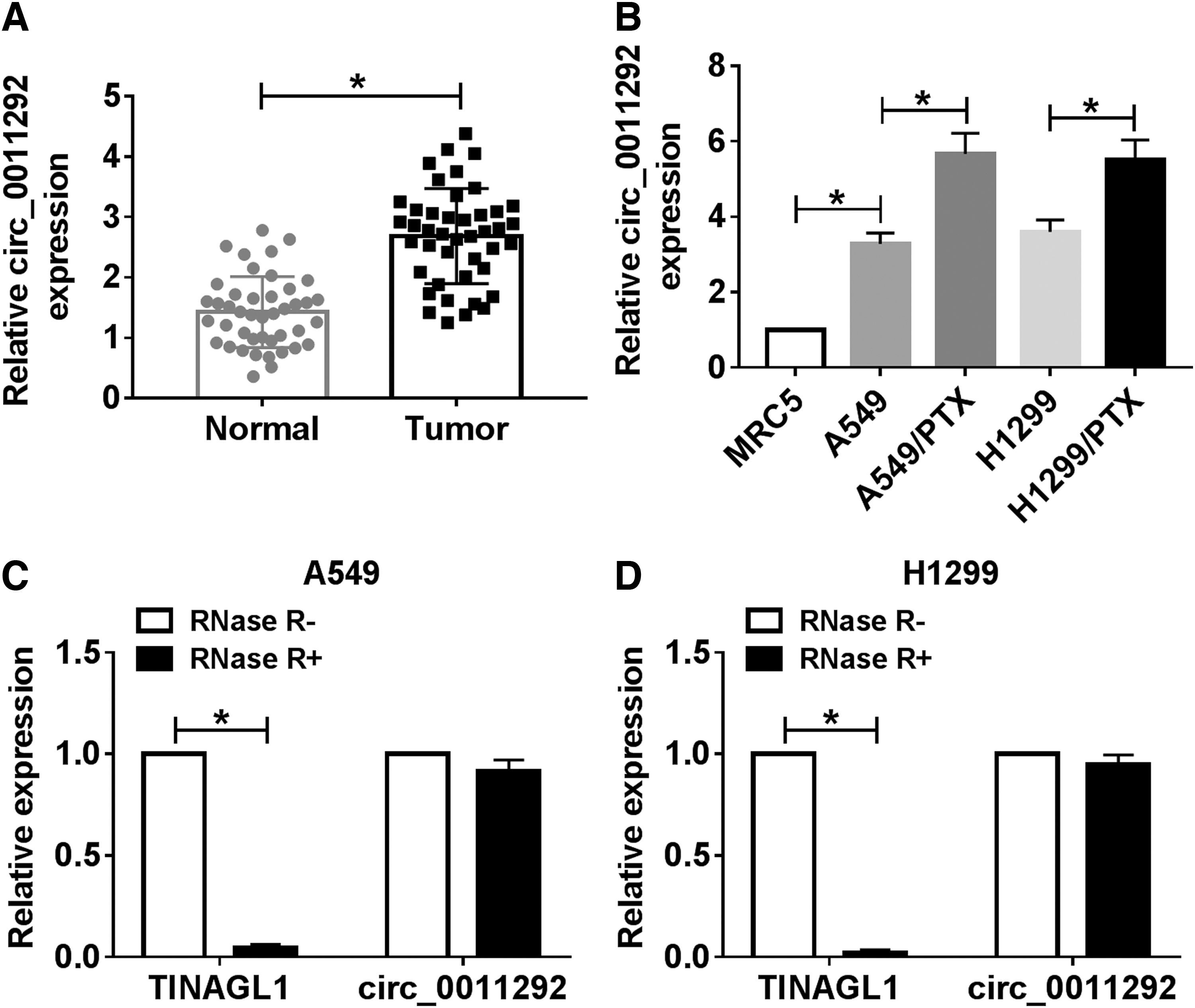
Circ_0011292 was upregulated in NSCLC tissues and PTX-resistant cells.
Knockdown of circ_0011292 hindered proliferation, migration, and invasion and promoted apoptosis in NSCLC cells
To investigate the function of circ_0011292 in the progression of NSCLC, A549 and H1299 cells were transduced with si-NC, si-circ_0011292#1, or si-circ_0011292#2. First of all, the knockdown efficiency was detected by qRT-PCR, and si-circ_0011292#1 with the highest efficiency was selected for subsequent research (Fig. 2A, B). CCK-8 assay showed that the viability of A549 and H1299 cells was drastically repressed after transfection with si-circ_0011292#1 (Fig. 2C, D). Flow cytometry suggested that inhibition of circ_0011292 prominently increased the apoptosis rate of A549 and H1299 cells (Fig. 2E). Moreover, transwell assay exhibited that knockdown of circ_0011292 inhibited NSCLC cell migration and invasion compared with the si-NC group (Fig. 2F, G). To verify the effect of circ_0011292 on the apoptosis and EMT of NSCLC cells, the expression of a series of related proteins was detected by Western blot. The results revealed that compared with the si-NC group, transfection of si-circ_0011292#1 resulted in a drastic decrease in Bcl-2, N-cadherin and vimentin levels, and a marked increase in c-caspase 3 and E-cadherin levels (Fig. 2H, I). These data evidenced that circ_0011292 silencing hindered NSCLC progression.
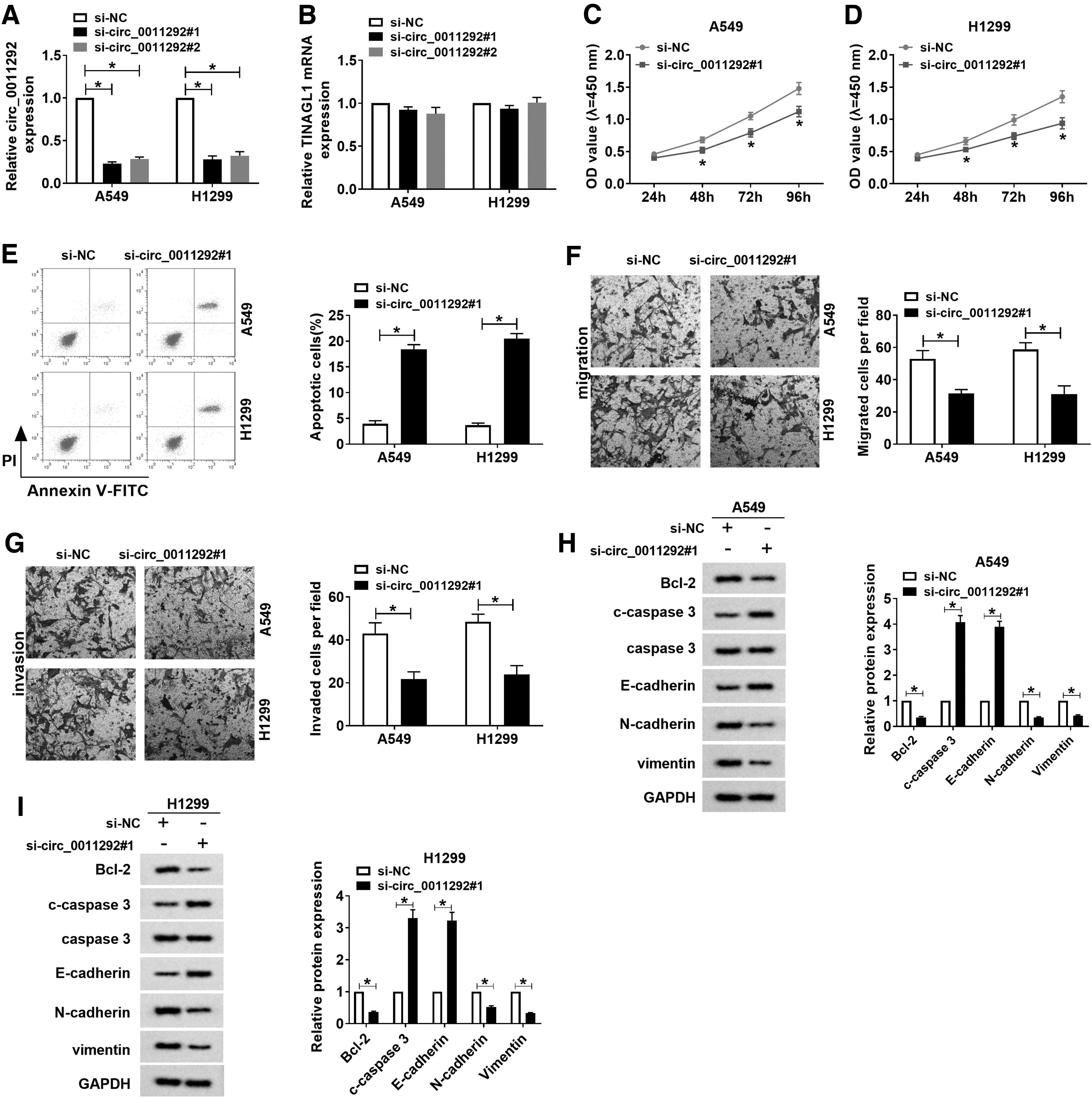
Knockdown of circ_0011292 hindered proliferation, migration, and invasion and promoted apoptosis in NSCLC cells.
Circ_0011292 depletion increased PTX sensitivity in PTX-resistant NSCLC cells
First, PTX-resistant cell lines were established. CCK-8 results showed that IC50 of PTX was overtly increased in A549/PTX and H1299/PTX cells compared to parental cells (Fig. 3A, B). Next, A549/PTX and H1299/PTX cells were introduced with si-NC or si-circ_0011292#1 to study the effect of circ_0011292 knockdown on PTX sensitivity. The results revealed that circ_0011292 silencing decreased the PTX resistance of A549/PTX and H1299/PTX cells (Fig. 3C). In addition, PTX treatment prominently increased the apoptosis rate of PTX-resistant cells compared with the control group, and the effect was strengthened after transfection with si-circ_0011292#1 (Fig. 3D). Consistently, PTX stimulation led to a significant decrease in Bcl-2 expression and an evident increase in c-caspase 3 expression, and these effects were aggrandized by downregulating circ_0011292 (Fig. 3E, F). Collectively, these data unveiled that knockdown of circ_0011292 enhanced PTX sensitivity in PTX-resistant NSCLC cells.
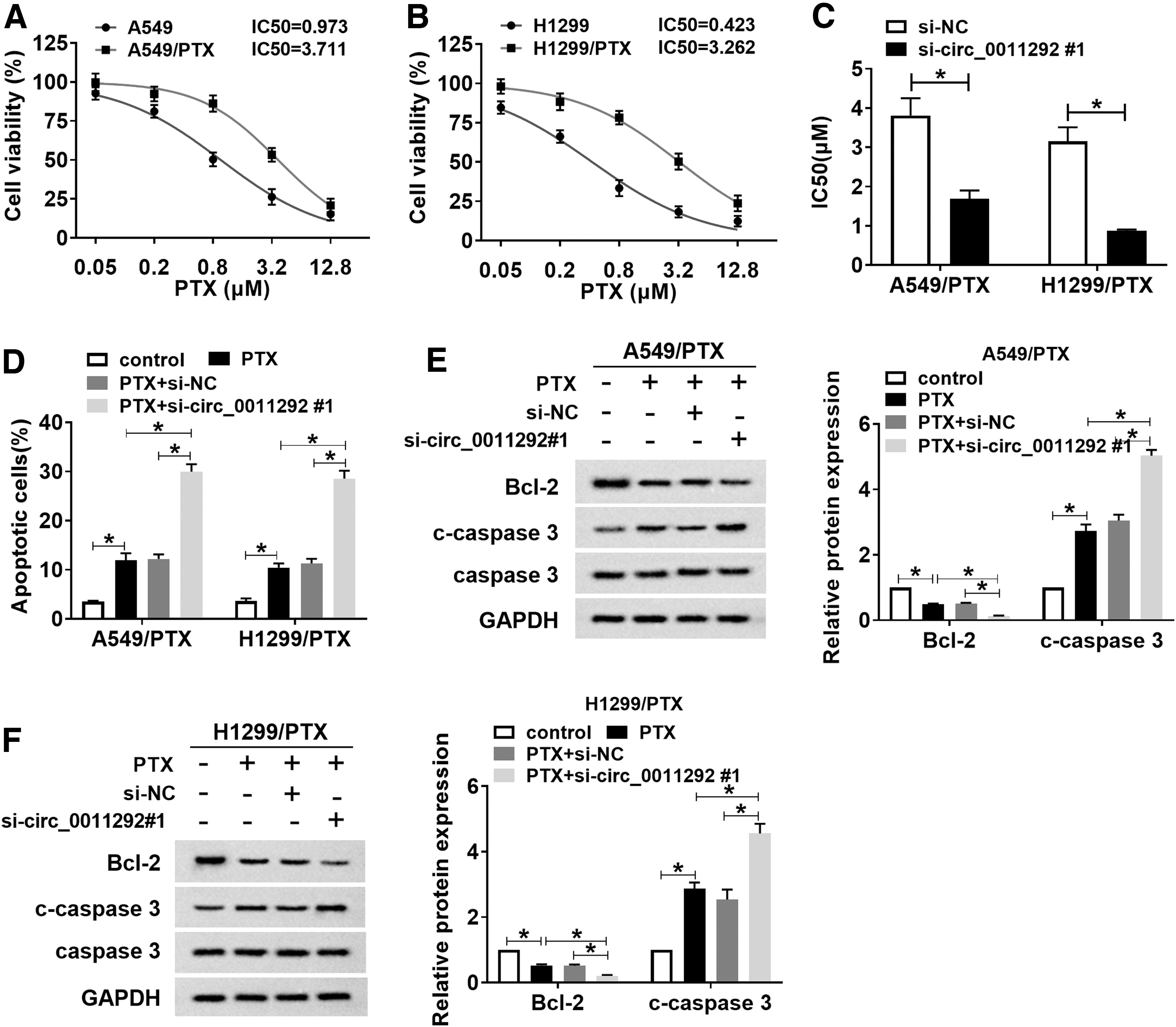
Circ_0011292 depletion increased PTX sensitivity in PTX-resistant NSCLC cells.
Circ_0011292 silencing enhanced PTX sensitivity of NSCLC in vivo
To elucidate the effect of circ_0011292 on PTX resistance in vivo, A549/PTX cells transfected with sh-NC or sh-circ_0011292 were subcutaneously injected into nude mice. After 6 days of injection, mice were intraperitoneally given PBS or 3 mg/kg PTX every 3 days. The results exhibited that PTX treatment or circ_0011292 inhibition could remarkably reduce tumor volume and weight, while simultaneous PTX treatment and circ_0011292 knockdown enhanced this inhibition effect (Fig. 4A, B). In addition, PTX stimulation decreased the protein level of proliferation marker PCNA, and the change was strengthened by transfection of sh-circ_0011292 (Fig. 4C). These data indicated that circ_0011292 knockdown potentiated PTX sensitivity of NSCLC in vivo.
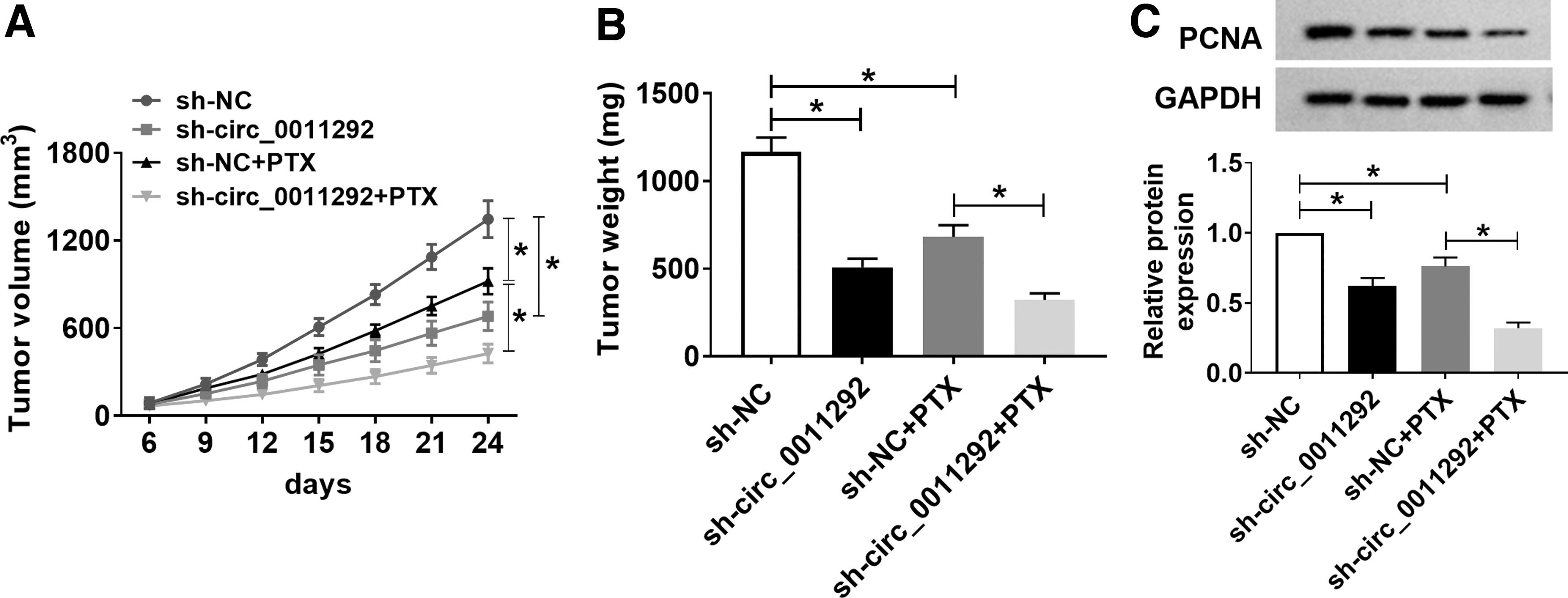
Circ_0011292 silencing enhanced PTX sensitivity of NSCLC in vivo. A549/PTX cells transfected with sh-NC or sh-circ_0011292 were subcutaneously injected into the mice. After 6 days, mice were injected with PBS or 3 mg/kg PTX every 3 days.
Circ_0011292 was a sponge of miR-379-5p
The authors predicted that miR-379-5p was a target of circ_0011292 through the starBase online database (Fig. 5A). To further verify the interaction between circ_0011292 and miR-379-5p, the dual-luciferase reporter assay was performed. The results suggested that miR-379-5p mimics strikingly decreased the luciferase activity of circ_0011292-wt reporter (Fig. 5B, C). Furthermore, RIP analysis showed that circ_0011292 and miR-379-5p were overtly enriched in the Ago2 group compared to the lgG group (Fig. 5D, E). Moreover, miR-379-5p level in A549 and H1299 cells was lower than that in MRC5 cells and higher than that in PTX-resistant cells (Fig. 5F). Transfection with circ_0011292 observably elevated circ_0011292 expression, but did not affect TINAGL1 expression (Fig. 5G, H). Besides, circ_0011292 upregulation reduced the expression of miR-379-5p, while circ_0011292 depletion increased the expression of miR-379-5p (Fig. 5I, J). Compared with adjacent normal tissues, miR-379-5p level was drastically decreased in NSCLC tissues (Fig. 5K). In NSCLC tissues, miR-379-5p expression was negatively correlated with circ_0011292 expression (Fig. 5L). These data concluded that circ_0011292 directly targeted miR-379-5p in NSCLC.

Circ_0011292 was a sponge of miR-379-5p.
TRIM65 was a target of miR-379-5p
Next, the starBase online database further predicted that miR-379-5p and TRIM65 3′UTR have complementary structure (Fig. 6A). To further validate the relationship between miR-379-5p and TRIM65, dual-luciferase reporter assay was carried out and showed a significant reduction in the luciferase activity of A549/PTX and H1299/PTX cells co-transfected with TRIM65 3′UTR-wt and miR-379-5p mimics (Fig. 6B, C). Moreover, RIP assay revealed that miR-379-5p and TRIM65 were co-precipitated by Ago2 antibody (Fig. 6D, E). In addition, the mRNA and protein levels of TRIM65 in A549 and H1299 cells were increased compared to MRC5 cells and reduced compared to PTX-resistant cells (Fig. 6F, G). QRT-PCR exhibited that miR-379-5p was prominently upregulated in PTX-resistant cells transfected with miR-379-5p mimics (Fig. 6H). Overexpression of miR-379-5p inhibited the mRNA and protein expression of TRIM65 (Fig. 6I, J). Also, the knockdown efficiency of miR-379-5p was detected in PTX-resistant cells transduced with anti-miR-NC or anti-miR-379-5p (Fig. 6K), and inhibition of miR-379-5p elevated the mRNA and protein levels of TRIM65 (Fig. 6L, M). Compared to normal tissues, the mRNA and protein levels of TRIM65 were distinctly increased in NSCLC tissues (Fig. 6N, O). Moreover, miR-379-5p and TRIM65 were negatively correlated in NSCLC tissues (Fig. 6P). These data indicated that TRIM65 was a target of miR-379-5p in NSCLC.

TRIM65 was a target of miR-379-5p.
MiR-379-5p regulated NSCLC progression and PTX resistance by targeting TRIM65
To investigate the relationship between miR-379-5p and TRIM65 in tumor progression and chemoresistance, A549/PTX and H1299/PTX were introduced with miR-NC, miR-379-5p, miR-379-5p+pcDNA, or miR-379-5p+TRIM65. The authors first discovered that TRIM65 transfection reversed the decrease in TRIM65 expression caused by miR-379-5p upregulation (Fig. 7A–C). Moreover, transfection with miR-379-5p conspicuously repressed cell proliferation, PTX resistance, and cell migration and invasion, while the effects were abrogated by upregulating TRIM65 (Fig. 7D–H). Also, miR-379-5p upregulation suppressed EMT by increasing E-cadherin level and reducing N-cadherin and vimentin levels, which was abolished after introduction with TRIM65 (Fig. 7I, J). Furthermore, PTX treatment induced cell apoptosis, and transfection of TRIM65 partially eliminated the proapoptosis effect triggered by miR-379-5p mimics (Fig. 7K, L). Simultaneously, the expression levels of apoptosis-associated proteins (Bcl-2 and c-caspase 3) confirmed the effect of miR-379-5p and TRIM65 on cell apoptosis (Fig. 7M, N). To sum up, these data manifested that miR-379-5p increased PTX sensitivity by targeting TRIM65 in PTX-resistant cells.

MiR-379-5p regulated NSCLC progression and PTX resistance by targeting TRIM65. A549/PTX and H1299/PTX cells were transduced with miR-NC, miR-379-5p, miR-379-5p+pcDNA, or miR-379-5p+TRIM65, respectively.
Circ_0011292 modulated NSCLC progression and PTX resistance by sponging miR-379-5p
The authors hypothesized that circ_0011292 sponged miR-379-5p to regulate NSCLC progression and PTX sensitivity. Therefore, they performed rescue experiments in A549/PTX and H1299/PTX transfected with si-NC, si-circ_0011292#1, si-circ_0011292#1+anti-miR-NC, or si-circ_0011292#1+anti-miR-379-5p. The results of qRT-PCR suggested that introduction of anti-miR-379-5p reversed the increase in miR-379-5p expression induced by circ_0011292 knockdown (Fig. 8A). Besides, circ_0011292 silencing remarkably impeded cell proliferation, PTX resistance, and cell migration and invasion, whereas co-transfection with si-circ_0011292#1 and anti-miR-379-5p alleviated the effects (Fig. 8B–F). In addition, inhibition of circ_0011292 caused a striking increase in E-cadherin level and significant decrease in N-cadherin and vimentin levels, which were reversed after transfection of anti-miR-379-5p (Fig. 8G, H). Moreover, inhibition of miR-379-5p partially alleviated the proapoptosis effect caused by circ_0011292 silencing (Fig. 8I, J). miR-379-5p downregulation partially reversed the decrease in Bcl-2 level and increase in c-caspase 3 level caused by circ_0011292 knockdown (Fig. 8K, L) while suppression of miR-379-5p reversed the inhibition of circ_0011292 depletion on TRIM65 mRNA and protein expression (Fig. 8M–O). In NSCLC tissues, circ_0011292 level was positively correlated with TRIM65 mRNA level (Fig. 8P). Collectively, these data reveal that circ_0011292 knockdown enhanced PTX sensitivity by sponging miR-379-5p in PTX-resistant cells.
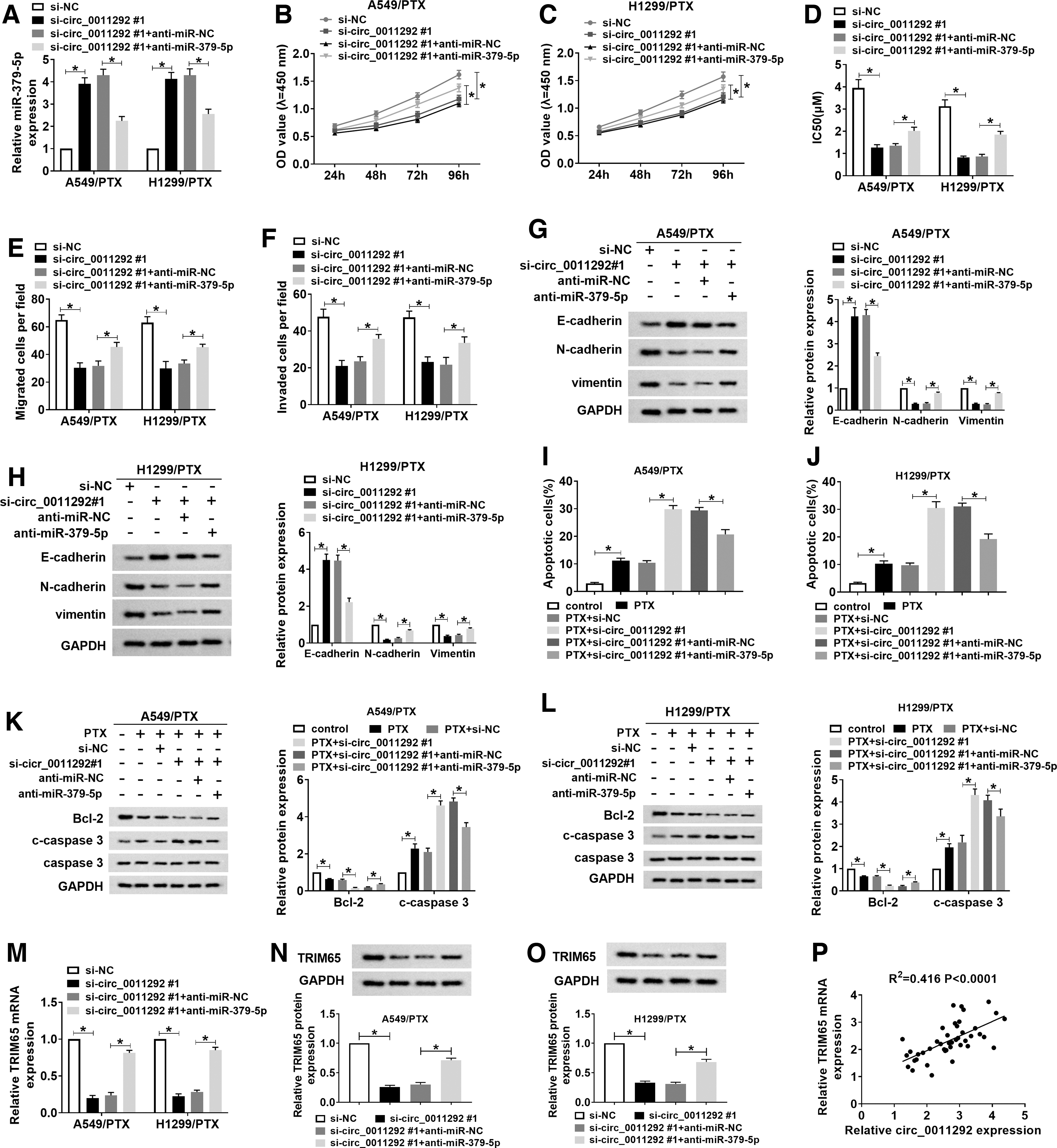
Circ_0011292 modulated NSCLC progression and PTX resistance by sponging miR-379-5p.
Discussion
Chemotherapy, including PTX, is a promising treatment for patients with advanced NSCLC who cannot undergo resection. 20 Drug resistance has become a significant obstacle limiting the efficacy of chemotherapy. 21 Moreover, EMT is related to enhanced stem cell properties and drug resistance, in addition to enhancing the mobility of tumor cells. 22 In addition, circRNA disorders are associated with chemoresistance in a variety of tumors. 23 For instance, circ-PVT1 improved PTX resistance by modulating the miR-124-3p/ZEB1 axis. 24 Therefore, to discover new biomarkers for overcoming chemoresistance, the authors investigated the role of circ_0011292 in PTX sensitivity of NSCLC.
Recent studies have suggested that circRNAs play several crucial roles in cellular physiology, including acting as competing endogenous RNAs (ceRNAs). 25,26 Moreover, the ceRNA mechanism has been studied a lot in the progression of NSCLC. For example, circ_0020123 increased the malignancy of NSCLC by serving as a ceRNA for miR-144 to elevate ZEB1 and EZH2 expression. 27 CircVANGL1 exerted a carcinogenic effect on NSCLC by functioning as ceRNA to regulate miR-195/Bcl-2 axis. 28 Circ_0020123 contributed to NSCLC tumorigenesis by sponging miR-488-3p and upregulating ADAM9. 29 Besides, some circRNAs have been demonstrated to be involved in drug resistance of NSCLC. 30 Li et al. revealed that circ_0002483 potentiated Taxol sensitivity in NSCLC by sponging miR-182-5p. 31 Dong et al. discovered that circ_0076305 enhanced cisplatin resistance in NSCLC by binding to miR-124-3p and increasing STAT3 expression. 32 In this research, the authors unveiled that circ_0011292 was predominantly raised in NSCLC tissues and PTX-resistant cells. More importantly, silencing of circ_0011292 impeded NSCLC progression and aggrandized PTX sensitivity.
Moreover, they selected the tumor suppressor miR-379-5p as a candidate through bioinformatics to study the mechanism of circ_0001721 in NSCLC. Chen et al. disclosed that miR-379-5p restrained hepatocellular carcinoma metastasis through deactivation of FAK/AKT pathway. 33 Wu et al. suggested that miR-379-5p curbed bladder cancer progression by binding to MDM2. 34 Lv et al. indicated that miR-379-5p functioned as a target of circ-PITX1 to block glioblastoma development by downregulating MAP3K2. 35 Furthermore, miR-379 improved cisplatin sensitivity in NSCLC by targeting EIF4G2. 36 In this study, the miR-379-5p level was drastically reduced in NSCLC tissues and PTX-resistant cells. Interestingly, circ_0011292 enhanced PTX resistance in NSCLC by sponging miR-379-5p in PTX-resistant NSCLC cells.
Recently, many members of the TRIM family have been verified to be significant regulators of tumorigenesis, including TRIM65. 37 In colorectal cancer, TRIM65 expedited tumor metastasis through degradation of ARHGAP35. 38 In lymphoma malignancies, TRIM65 triggered tumorigenesis by regulating ERK1/2 pathway. 39 In ovarian cancer, TRIM65 facilitated tumor metastasis by upregulating EMT-related protein vimentin. 40 Besides, TRIM65 level in cisplatin-resistant cells was remarkably increased in comparison with the parental NSCLC cells, and TRIM65 depletion suppressed cisplatin resistance in cisplatin-resistant NSCLC cells by modulating miR-138-5p/ATG7 axis. 41 Similar to previous study, the authors manifested that TRIM65 was dramatically upregulated in NSCLC tissues and cells. Importantly, miR-379-5p increased PTX sensitivity by targeting TRIM65 in PTX-resistant NSCLC cells.
Conclusion
In summary, circ_0011292 promoted tumor progression and enhanced PTX resistance in NSCLC by sponging miR-379-5p to upregulate TRIM65. These findings provided new biomarkers for NSCLC to overcome chemotherapy resistance.
Footnotes
Authors' Contributions
Conception and design: C.G.; development of methodology: H.W.; acquisition of data: H.J.; analysis and interpretation of data: L.Q.; writing, review, and revision of article: X.W.; all coauthors have reviewed and approved the article before submission.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.