
Abstract
Background:
Hepatocellular carcinoma (HCC) is the most common form of liver cancer. Circular RNAs (circRNAs) play a vital role in cancer development and progression. This study investigated the role and potential mechanism of circRNA filamin binding LIM protein 1 (circFBLIM1) in HCC.
Methods:
Exosomes were identified by transmission electron microscopy (TEM), nanoparticle tracking analysis (NTA), and Western blot assay. The levels of circFBLIM1, miR-338, and low-density lipoprotein receptor-related protein 6 (LRP6) were measured by quantitative real-time polymerase chain reaction or Western blot. Glycolysis was analyzed by detecting glucose consumption, lactate production, ATP level, extracellular acidification rate (ECAR), and oxygen consumption rate (OCR). Cell viability was evaluated by 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) assay. Cell apoptosis was detected by flow cytometry. Xenograft assay was performed to analyze tumor growth in vivo. The interaction among circFBLIM1, miR-338, and LRP6 was confirmed by dual-luciferase reporter assay and RNA immunoprecipitation (RIP) assay. This study was approved by the Institutional Review Board of the First Affiliated Hospital of Guizhou University of Traditional Chinese Medicine.
Results:
CircFBLIM1 was highly expressed in HCC serum exosomes and HCC cells. Inhibition of circFBLIM1 confined HCC glycolysis and progression. CircFBLIM1 knockdown blocked tumorigenesis in vivo. CircFBLIM1 was a sponge of miR-338 and promoted HCC progression and glycolysis by regulating miR-338. Moreover, miR-338 suppressed HCC progression and glycolysis via targeting LRP6. Mechanistically, circFBLIM1 functioned as an miR-338 sponge to upregulate LRP6.
Conclusion:
CircFBLIM1 facilitated HCC progression and glycolysis via modulating the miR-338/LRP6 axis, which may provide promising therapeutic targets for HCC.
Introduction
Liver cancer ranks fourth in tumor mortality, with hepatocellular carcinoma (HCC) accounting for 75%–85% of primary liver cancer. 1,2 HCC is principally caused by hepatitis B or C virus (HBV or HCV) infection. 3 Due to the high rate of metastasis and recurrence, the 5-year survival rate of HCC patients remains as low as 18%. 4 Therefore, novel treatment strategies are urgently needed to improve the progression of liver cancer.
The Warburg effect is defined as the cancer cells rely on aerobic glycolysis to obtain the energy needed for cellular processes. 5 Exosomes are extracellular vesicles with a diameter between 30 and 150 nm. 6 In addition, exosomes contain many active biomolecules, such as lipids, proteins, and nucleic acids, 7 which are involved in intercellular communication by transferring exosomes. 8 Recently, studies have confirmed that circular RNAs are strikingly enriched and stably expressed in exosomes. 9
Circular RNAs (circRNAs) are a subgroup of noncoding RNAs with covalent closed-loop structures. 10 A growing number of studies have unveiled that dysregulated circRNAs are closely related to tumor pathology and prognosis, including HCC. 11 For example, circHIAT1 curbed the growth of HCC cells via sponging microRNA-3171 to upregulate PTEN. 12 CircMYLK facilitated HCC development by modulating the microRNA-362-3p/Rab23 axis. 13 Circ_0070269 impeded HCC progression by downregulating microRNA-182 and elevating NPTX1 expression. 14 Human reference genome (GRCh37/hg19) showed that hsa_circ_0010090 located at chr1: 16084668–16113084 is derived from filamin binding LIM protein 1 (FBLIM1) and is called “circFBLIM1.” In addition, previous research has verified that circFBLIM1 expedited HCC progression by sponging microRNA-346. 15 However, the exact mechanism of circFBLIM1 in HCC needs further investigation.
MicroRNAs (miRNAs) are short endogenous noncoding RNAs with a length of 19–25 nucleotides, which exert a regulatory effect on tumor progression. 16 Growing evidence has manifested that miRNAs participate in HCC development and thus become biomarkers for monitoring carcinogenesis. 17 Previous research disclosed that miR-338 expression was drastically diminished in HCC tissues by bead-based miRNA expression profiling. 18 However, the association between circFBLIM1 and miR-338 remains unclear.
Low-density lipoprotein receptor-related protein 6 (LRP6), as the Wnt coreceptor, exerts a crucial role in the Wnt/β-catenin pathway. 19 LRP6 gene mutations are associated with severe human diseases, including Alzheimer's disease, osteoporosis, and cancer. 20 In HCC, LRP6 contributes to tumor progression via activating the Wnt/β-catenin pathway. 21
Therefore, the authors explored the function and potential mechanisms of exosomal circFBLIM1 in HCC progression and glycolysis.
Materials and Methods
Serum samples
Thirty HCC patients and 25 age-matched healthy volunteers were recruited from The First Affiliated Hospital of Guizhou University of Traditional Chinese Medicine. Five milliliters of venous blood was collected from each participant, and the serum was obtained by centrifugation. All participants signed written informed consent. The research was ratified by the Ethics Committee of The First Affiliated Hospital of Guizhou University of Traditional Chinese Medicine.
Cell culture
Human normal hepatocyte cells THLE-2 and HCC cells SNU-387 were commercially obtained from American Type Culture Collection (ATCC, Manassas, VA). The HCC cell line Huh7 was purchased from Honsun Biological Technology Co., Ltd. (Shanghai, China). All cells were cultured in RPMI-1640 medium (Gibco, Los Angeles, CA) supplemented with 10% fetal bovine serum (FBS; Gibco) at 37°C.
Exosome isolation and identification
Serum exosomes were extracted following the procedure described previously. 22 In brief, serum was centrifuged at 3000 g for 10 min to precipitate cells and then centrifuged at 10,000 g for 30 min to remove cell debris. Then, the collected supernatant was ultracentrifuged at 110,000 g for 60 min, and the exosomes were resuspended in PBS, followed by ultracentrifugation for 60 min to obtain pure exosomes. Next, the morphology and size of exosomes were observed using a Philips CM120 transmission electron microscope (TEM) (Philips Research, Eindhoven, Netherlands) at 80 kV. The size distribution of exosomes was analyzed by nanoparticle tracking analysis (NTA). The expression of the exosome marker was examined by Western blot assay.
Cell transfection
CircFBLIM1 overexpression vector (circFBLIM1) and the empty vector (pLVX), silencing RNA (siRNA) against circFBLIM1 (si-circFBLIM1) and the siRNA control (si-NC), miR-338 mimic and the mimic control (miRNA NC), miR-338 inhibitor and the inhibitor control (inhibitor NC), LRP6 overexpression vector (pc-LRP6) and the empty vector (pc-NC) were obtained from GeneChem (Shanghai, China). The oligonucleotides and plasmids were transfected into SNU-387 and Huh7 cells using Lipofectamine 3000 (Invitrogen, Carlsbad, CA).
Quantitative real-time polymerase chain reaction
After isolating RNA with TRIzol (Invitrogen), RNA was reverse transcribed using the HiScript II One Step RT-PCR Kit (Vazyme, Nanjing, China) or miScript II RT Kit (Qiagen, Frankfurt, Germany). Subsequently, quantitative PCR was performed using AceQ qPCR SYBR Green Master Mix (Vazyme). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or U6 was taken as an internal control. Primers are shown below: circFBLIM1 (F: 5′-CGGATCCCAGAGCCAAAGTAA-3′, R: 5′-GTTCTTCATGGACCCCGAGT-3′.), miR-338 (F: 5′-TGCGGTCCAGCATCAGTGAT-3′, R: 5′-CCAGTGCAG GGTCCGAGGT-3′.), LRP6 (F: 5′-ACGATTGTAGTTGGAGGCTTG-3′, R: 5′-ATGGCTTCTTCGCTGACATCA-3′.), GAPDH (F: 5′-GGGAAACTGTGGCGTGAT-3′, R: 5′-GAGTGGGTGTCGCTGTTGA-3′.), and U6 (F: 5′-CTCGCTTCGGCAGCACA-3′, R: 5′-AACGCTTCACGAATTTGCGT-3′.).
Western blot assay
Protein was lysed with RIPA buffer (Solarbio, Beijing, China). After separating the same amount of protein by polyacrylamide gel electrophoresis, the separated protein was transferred to a polyvinylidene fluoride membrane (Millipore, Billerica, MA). Next, the membranes were probed with primary antibodies against CD63 (ab216130; Abcam, Cambridge, United Kingdom), LRP6 (ab134146; Abcam), or GAPDH (ab9485; Abcam) after reacting with 5% skim milk for 2 h. Subsequently, the membranes interacted with the secondary antibody (ab7090; Abcam). Last, the bands were emerged by the enhanced chemiluminescence system (Solarbio).
Measurement of glucose consumption, lactate production, and ATP level
Glucose consumption and lactate production were detected by the Glucose Assay Kit and Lactic Acid Kit (Abcam). ATP level was determined using the ATP Colorimetric Assay Kit (AmyJet Scientific, Wuhan, China).
Detection of extracellular acidification rate and oxygen consumption rate
Extracellular acidification rate (ECAR) and oxygen consumption rate (OCR) were detected using the Seahorse XFe 96 Extracellular Flux Analyzer (Seahorse Bioscience, North Billerica, MA). Briefly, cells (2 × 104) were seeded into a Seahorse XF 96 cell culture microplate. For ECAR, glucose, oligomycin, and 2-deoxyglucose (2-DG) were added in sequence at the indicated time points; and for OCR, oligomycin, p-trifluoromethoxy carbonyl cyanide phenylhydrazone (FCCP), and rotenone were added in turn. Last, data were analyzed using the Seahorse XF-96 Wave software.
Cell viability assay
Cells (3 × 103) were seeded into 96-well plates. Next, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) solution (Solarbio) was added to each well after incubation for 0, 24, 48, and 72 h. Then, cells were treated with dimethyl sulfoxide (DMSO; Solarbio) for 15 min. Finally, cell viability was assessed using Synergy Neo2 Hybrid Multimode Reader (BioTek, Burlington, VT).
Cell apoptosis assay
Transfected cells were incubated with an Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) kit (BestBio, Shanghai, China). Then, the apoptotic cells were monitored using an Attune NxT Flow Cytometer (Thermo Fisher Scientific, Waltham, MA).
Xenograft tumor experiment
Huh7 cells were stably transfected with the lentivirus carrying shRNA against circFBLIM1 (sh-circFBLIM1) or the negative control (sh-NC) (Genelily BioTech, Shanghai, China). Then, seven BALB/c nude mice (5 weeks old) were randomly selected from each group, and transfected cells (5 × 106) were subcutaneously injected into the right abdomen of the mice. Then, tumor volume was examined every 4 d. Twenty-seven days after injection, the mice were sacrificed, and the xenograft was weighed. The animal experiment was ratified by the Animal Research Committee of The First Affiliated Hospital of Guizhou University of Traditional Chinese Medicine.
Dual-luciferase reporter assay
The sequences of circFBLIM1 or LRP6–3′UTR containing wild-type or mutant binding sites of miR-338 were inserted into the pmirGLO vector (Promega, Madison, WI). The corresponding luciferase reporter and miR-338 mimic or miRNA NC were cotransfected into 293T cells. Finally, the luciferase activity was detected by the Dual-Lucy Assay Kit (Solarbio).
RNA immunoprecipitation assay
RNA immunoprecipitation (RIP) analysis was performed using the EZ-Magna RIP kit (Millipore). First, RIP lysis buffer was used to lyse SNU-387 and Huh7 cells introduced with miR-338 mimic or miRNA NC. Then, cell lysates were incubated with magnetic beads conjugated with the AGO2 antibody or IgG antibody.
Statistical analysis
Data are displayed as mean ± standard deviation by using GraphPad Prism 7 software (GraphPad, Inc., La Jolla, CA). Differences were compared using Student's t-test or one-way analysis of variance. When p < 0.05, it was considered statistically significant.
Results
CircFBLIM1 was upregulated in serum-derived exosomes and HCC cells
First, exosomes were isolated from the serum of HCC patients and healthy volunteers. The authors observed the characteristics of these vesicles with a TEM (Fig. 1A). The size distribution of particles was detected by NTA, and the results exhibited that the particle size range was 50–175 nm (Fig. 1B). Also, Western blot assay confirmed the presence of exosome marker CD63 in extracted exosomes (Fig. 1C). In addition, circFBLIM1 expression in serum exosomes of HCC patients was strikingly increased compared with healthy volunteers (Fig. 1D). Relative to the normal hepatocyte cell line THLE-2, the level of circFBLIM1 was drastically elevated in SNU-387 and Huh7 cells (Fig. 1E). Then, SNU-387 and Huh7 cells were incubated with serum exosomes, and the results revealed that circFBLIM1 level was overtly increased in the cells after incubation (Fig. 1F, G). These results implied that exosomal circFBLIM1 might play a vital role in HCC.
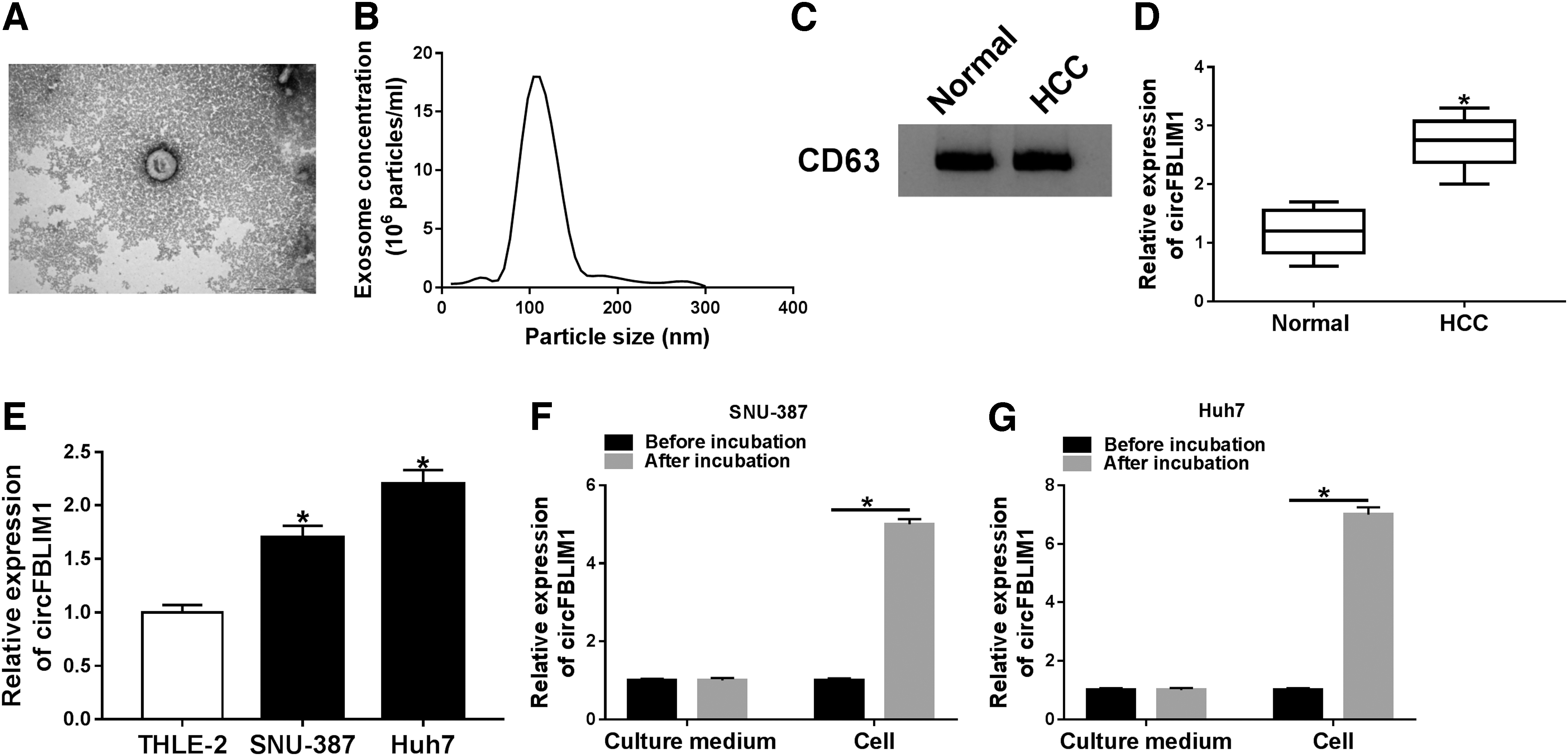
CircFBLIM1 was upregulated in serum-derived exosomes and HCC cells.
CircFBLIM1 expression was associated with glycolysis
To determine whether circFBLIM1 is related to glycolysis, the authors examined two glycolysis-related organic acids (G-6-P and L-Lactate) in the serum of HCC patients. The results suggested that the levels of G-6-P and L-Lactate were remarkably increased in serum of HCC patients compared with the normal group (Fig. 2A, B). Besides, transfection with circFBLIM1 prominently increased circFBLIM1 expression, but did not affect the FBLIM1 mRNA level (Fig. 2C, D). Conversely, circFBLIM1 expression was downregulated after introduction with si-circFBLIM1 (Fig. 2E, F). CircFBLIM1 overexpression in SNU-387 and Huh7 cells markedly accelerated pH decline in the medium (Fig. 2G, H), whereas silencing of circFBLIM1 in SNU-387 and Huh7 cells prominently inhibited pH decline (Fig. 2I, J). As shown in Supplementary Figure S1, circFBLIM1 expression was positively correlated with G-6-P or L-Lactate level in HCC. These data discovered that highly expressed circFBLIM1 was involved in glycolysis.

CircFBLIM1 expression was associated with glycolysis.
CircFBLIM1 induced glycolysis in HCC cells
To further elucidate the role of circFBLIM1 in glycolysis, a series of glycolysis-related indicators were determined in SNU-387 and Huh7 cells transfected with pLVX, circFBLIM1, si-NC, or si-circFBLIM1. First, overexpression of circFBLIM1 increased glucose consumption, while silencing of circFBLIM1 inhibited glucose consumption compared with the control group (Fig. 3A, B). Also, circFBLIM1 upregulation expedited lactate production, whereas circFBLIM1 downregulation curbed lactate production (Fig. 3C, D). ATP level was increased after transfection with circFBLIM1, while ATP level was decreased when circFBLIM1 was silenced (Fig. 3E, F). In addition, ECAR reflects the overall glycolytic flux, and OCR reflects mitochondrial respiration. The results suggested that overexpression of circFBLIM1 resulted in a noticeable increase in ECAR and a marked decrease in OCR (Fig. 3G, I), while silenced circFBLIM1 significantly reduced ECAR and elevated OCR (Fig. 3H, J). These data evidenced that circFBLIM1 facilitated glycolysis in HCC cells.
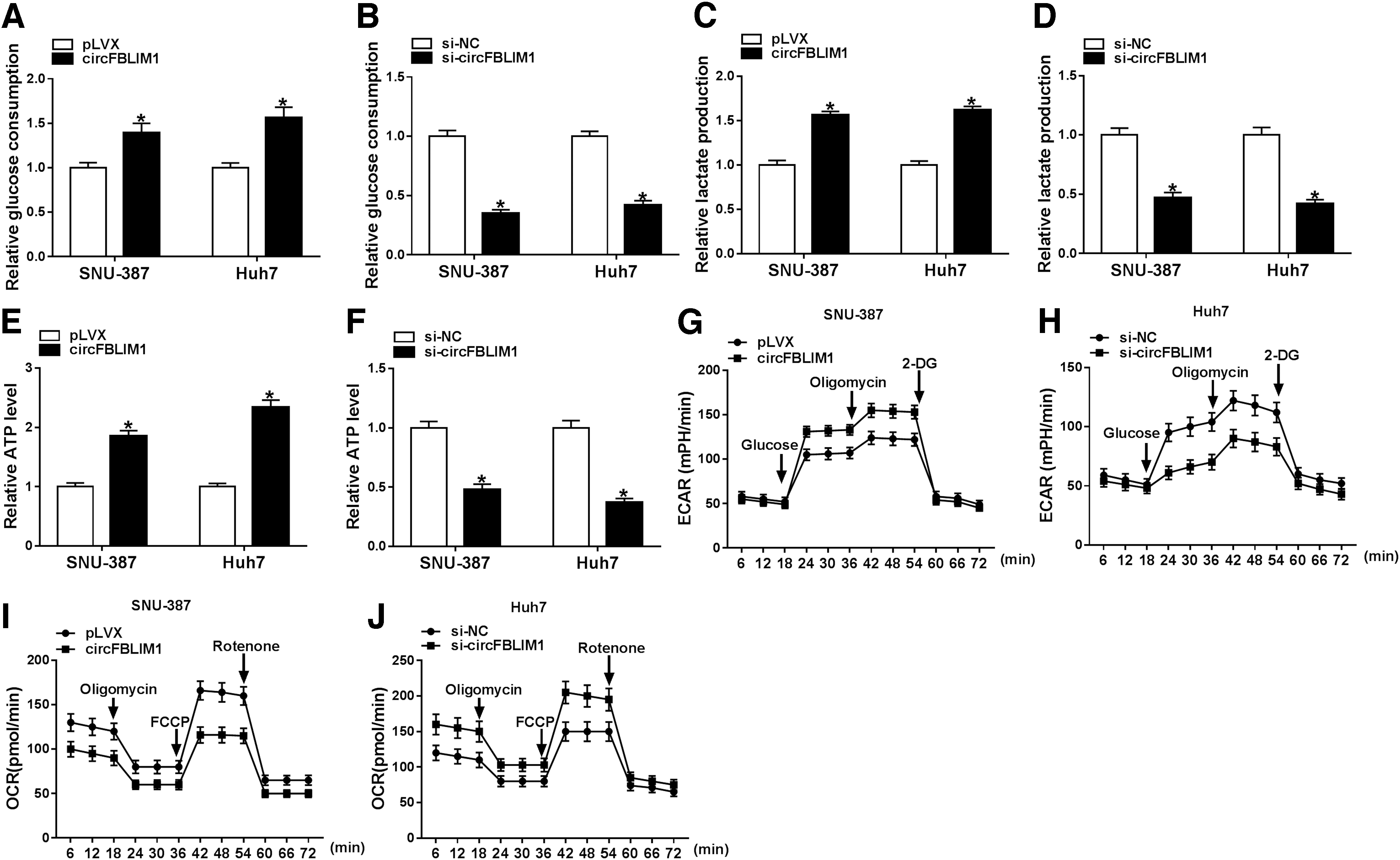
CircFBLIM1 induced glycolysis in HCC cells. SNU-387 and Huh7 cells were introduced with pLVX, circFBLIM1, si-NC, or si-circFBLIM1, respectively.
CircFBLIM1 knockdown hindered HCC cell progression and tumor growth in vivo
To explore the role of circFBLIM1 in tumor progression, MTT analysis and flow cytometry were performed in SNU-387 and Huh7 cells transfected with si-NC or si-circFBLIM1. The results showed that the viability of SNU-387 and Huh7 cells was strikingly decreased in the si-circFBLIM1 group compared with the si-NC group (Fig. 4A, B). Also, knockdown of circFBLIM1 remarkably accelerated the apoptosis rate of SNU-387 and Huh7 cells (Fig. 4C, D). The authors also constructed a xenograft mouse model to analyze tumor growth in vivo, and the results revealed that tumor volume and weight were overtly reduced in the sh-circFBLIM1 group relative to the sh-NC group (Fig. 4E, F). In addition, circFBLIM1 expression was distinctly decreased in the sh-circFBLIM1 group compared with the sh-NC group (Fig. 4G). These data indicated that depletion of circFBLIM1 blocked HCC cell progression and tumor growth in vivo.

CircFBLIM1 knockdown hindered HCC cell progression and tumor growth in vivo.
CircFBLIM1 regulated the progression and glycolysis of HCC by sponging miR-338
According to the starBase website, circFBLIM1 may be a molecular sponge of miR-338 (Fig. 5A). This prediction was confirmed by the dual-luciferase reporter assay, which miR-338 mimic remarkably reduced the luciferase activity of WT-circFBLIM1 reporter (Fig. 5B). Furthermore, RIP assay exhibited that circFBLIM1 and miR-338 were enriched by the AGO2 antibody in SNU-387 and Huh7 cells overexpressing miR-338 (Fig. 5C, D). To elucidate the role of miR-338 as a target of circFBLIM1 in HCC, SNU-387 and Huh7 cells were transfected with si-NC, si-circFBLIM1, si-circFBLIM1+inhibitor NC, or si-circFBLIM1+miR-338 inhibitor. First, cotransfection of si-circFBLIM1 and miR-338 inhibitor abrogated the increased expression of miR-338 caused by circFBLIM1 silencing (Fig. 5E). MTT assay indicated that knockdown of circFBLIM1 blocked the viability of SNU-387 and Huh7 cells, whereas the effect was reversed by downregulating miR-338 (Fig. 5F, G). Flow cytometry showed that transfection with the miR-338 inhibitor abolished the promotion of circFBLIM1 depletion on cell apoptosis (Fig. 5H). In addition, circFBLIM1 silencing conspicuously decreased glucose consumption, lactate production, and ATP level, which were reversed after transfection with the miR-338 inhibitor (Fig. 5I–K). Also, circFBLIM1 knockdown resulted in a marked reduction in ECAR and a notable increase in OCR, whereas inhibition of miR-338 overturned the effects (Fig. 5L–O). These data manifested that circFBLIM1 sponged miR-338 to modulate HCC progression and glycolysis.
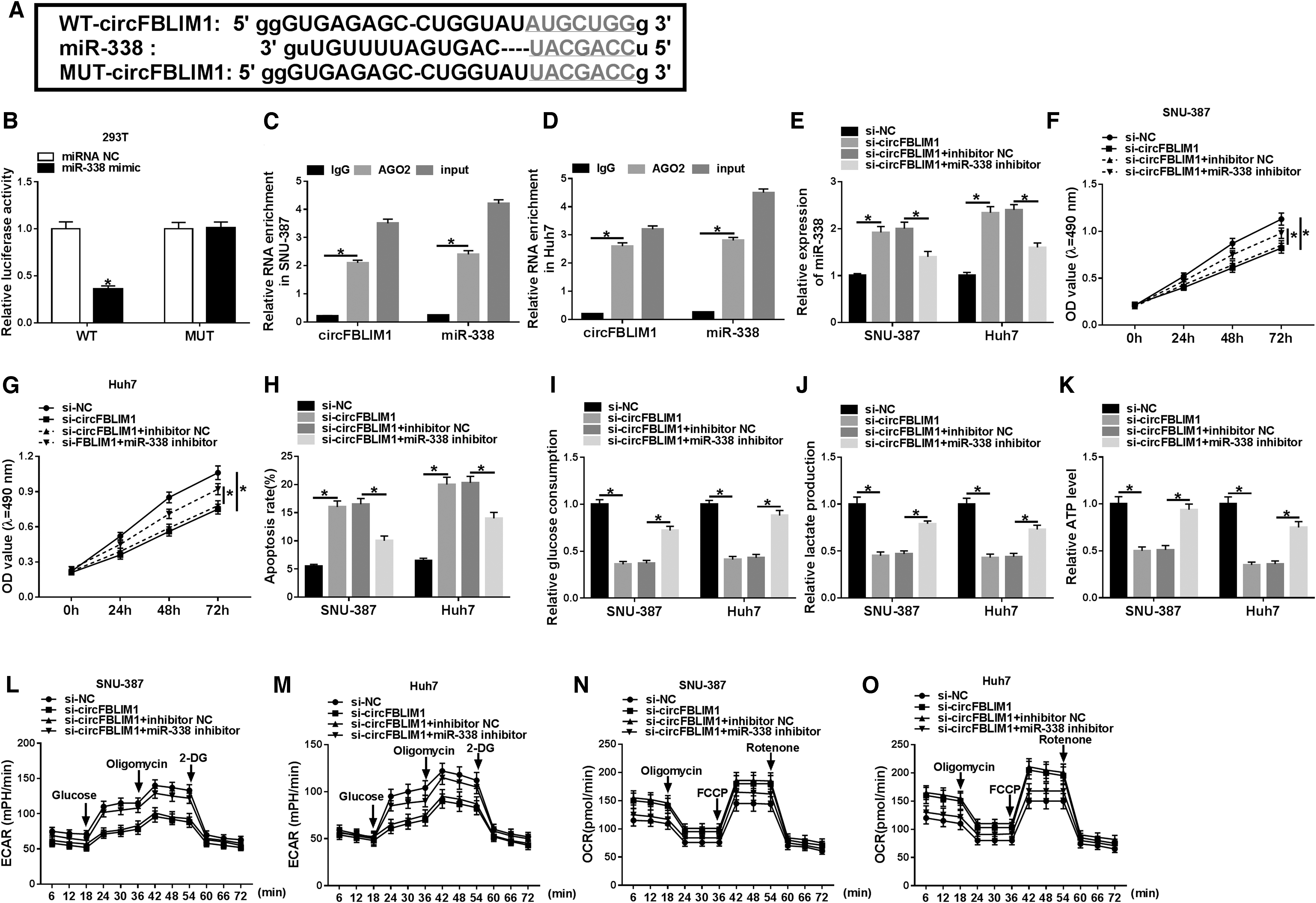
CircFBLIM1 regulated the progression and glycolysis of HCC by sponging miR-338.
MiR-338 modulated the progression and glycolysis of HCC via targeting LRP6
Online database starBase predicted that LRP6 might be a target gene of miR-338 (Fig. 6A). Dual-luciferase reporter assay suggested that miR-338 mimic observably reduced the luciferase activity of WT-LRP6–3′UTR reporter (Fig. 6B). The authors also performed RIP assay to verify whether miR-338 bound to LRP6, and the results exhibited that miR-338 and LRP6 were enriched by the AGO2 antibody in miR-338-overexpressed SNU-387 and Huh7 cells (Fig. 6C, D). Moreover, transfection with miR-338 mimic distinctly increased miR-338 expression relative to the control group (Fig. 6E). To investigate the effect of miR-338 and LRP6 on HCC development, SNU-387 and Huh7 cells were introduced with miRNA NC, miR-338 mimic, miR-338 mimic+pc-NC, or miR-338 mimic+pc-LRP6. The results of quantitative real-time polymerase chain reaction (qRT-PCR) and Western blot assay presented that upregulation of LRP6 undermined the decreased expression of LRP6 induced by miR-338 overexpression (Fig. 6F, G). Besides, cotransfection of miR-338 mimic and pc-LRP6 attenuated the inhibitory effect of miR-338 mimic on cell viability (Fig. 6H, I). Increased apoptosis rate caused by miR-338 upregulation was reversed by elevating LRP6 expression (Fig. 6J). In addition, upregulation of LRP6 abated the inhibitory effect of miR-338 overexpression on glucose consumption, lactate production, and ATP level (Fig. 6K–M). Transfection of miR-338 mimic led to a remarkable decrease in ECAR and a sharp increase in OCR, while these effects were abrogated by upregulating LRP6 (Fig. 6N–Q). These data concluded that miR-338 targeted LRP6 to regulate HCC progression and glycolysis.
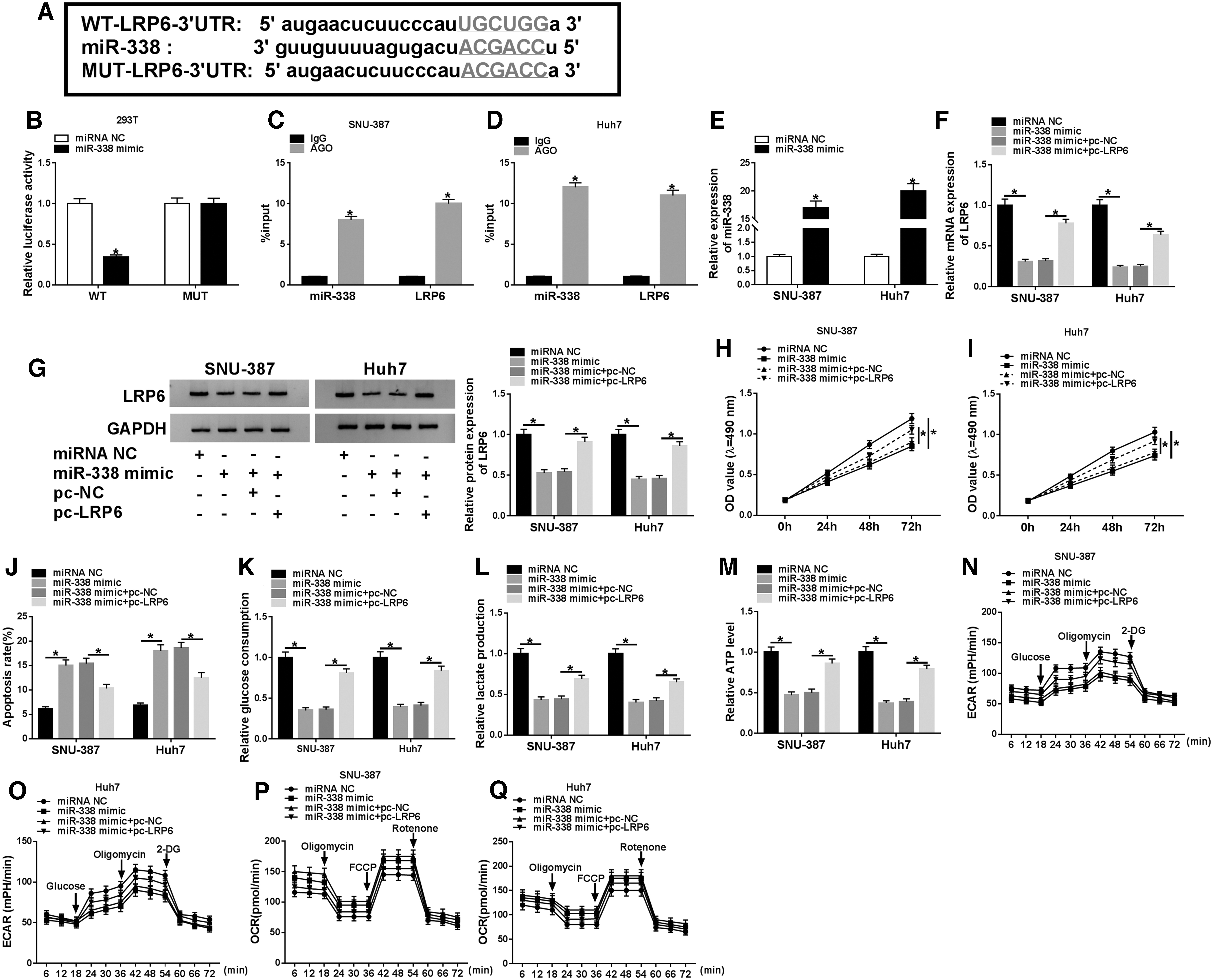
MiR-338 modulated the progression and glycolysis of HCC via targeting LRP6.
CircFBLIM1 increased LRP6 expression by sponging miR-338
To clarify the relationship among circFBLIM1, miR-338, and LRP6, LRP6 expression was detected by qRT-PCR and Western blot in SNU-387 and Huh7 cells transfected with si-NC, si-circFBLIM1, si-circFBLIM1+pc-NC, si-circFBLIM1+pc-LRP6, si-circFBLIM1+inhibitor NC, or si-circFBLIM1+miR-338 inhibitor, respectively. The results exhibited that circFBLIM1 silencing drastically reduced the mRNA and protein levels of LRP6, whereas LRP6 expression was restored after transfection with pc-LRP6 (Fig. 7A, B). In addition, suppression of miR-338 reversed the inhibition of circFBLIM1 knockdown on LRP6 mRNA and protein levels (Fig. 7C, D). These data evidenced that circFBLIM1 regulated LRP6 expression via sponging miR-338 in HCC cells.

CircFBLIM1 increased LRP6 expression by sponging miR-338.
Discussion
Glycolysis is a hallmark feature of cancer; especially cancer cells preferentially use glycolysis to produce glucose-dependent ATP. 23 Lactate is an important metabolite in cancer metabolic reprogramming, and its large accumulation results in extracellular pH in the tumor microenvironment being acidified, ranging from 6.0 to 6.5. 24 Besides, tumor-derived exosomes are strongly associated with the tumor microenvironment. 25 In this study, circFBLIM1 expression was overtly elevated in HCC serum exosomes and HCC cells. More importantly, circFBLIM1 carried by serum exosomes could be transferred to HCC cells.
Increasing evidence has corroborated that circRNAs participate in tumor biological processes via acting as competing endogenous RNAs (ceRNAs). 26 Besides, emerging studies suggested that circRNAs compete with miRNAs to modulate the expression of target genes, thus affecting HCC progression. For example, Yu et al. revealed that circRNA-104718 triggered HCC development by sponging microRNA-218-5p to elevate TXNDC5 expression. 27 Liu et al. disclosed that circRNA-5692 hindered HCC progression by serving as a ceRNA through the miR-328-5p/DAB2IP axis. 28 In addition, adipocyte-secreted circRNAs facilitated HCC growth via inhibiting miR-34a and upregulating USP7. 29 Previous research presented that higher metastatic HCC cells transferred the metastatic potential to low-metastatic or nonmetastatic HCC cells through exosomal circPTGR1, thereby improving the metastatic ability of HCC cells. 30 In this study, the authors verified that silenced circFBLIM1 suppressed HCC glycolysis and progression.
To elucidate the potential basis of circFBLIM1 in HCC, they screened miRNAs that play an anticancer role in HCC, and demonstrated that miR-338 could bind to circFBLIM1 based on bioinformatics. Numerous studies have indicated that miR-338 was a suppressing factor in a variety of tumors. For instance, Zhang et al. discovered that inhibition of microRNA-338-3p accelerated angiogenesis in HCC. 31 Wang et al. showed that microRNA-338-3p targeted FOXP4 to repress HCC cell proliferation. 32 Also, recent research exhibited that microRNA-338 could implicate in regulating glycolysis of HCC cells. 33 Besides, the authors first confirmed that circFBLIM1 induced the progression and glycolysis of HCC via sponging miR-338.
LRP6 is a fundamental coreceptor for WNT and is usually involved in the activation of Wnt/β-catenin pathway in epithelial cancer. 34 In triple-negative breast cancer, LRP6 expedited tumor metastasis by modulating S100A4 through the Wnt/β-catenin pathway. 35 In addition, accumulating evidence has validated that miRNAs bound to target gene 3′UTR to regulate gene expression. 36 For instance, LRP6 bound to microRNA-454 to regulate angiogenesis and metastasis in pancreatic ductal adenocarcinoma. 37 Besides, miR-126-3p targeted LRP6 and PIK3R2 to curb HCC angiogenesis and metastasis. 38 This study indicated that LRP6 upregulation reversed the inhibitory effect of miR-338 mimic on HCC progression and glycolysis.
Taken together, these findings verified that exosomal circFBLIM1 contributed to the progression and glycolysis of HCC by sponging miR-338 and upregulating LRP6. The circFBLIM1/miR-338/LRP6 axis may provide a new therapeutic approach for HCC treatment.
Footnotes
Disclosure Statement
The authors declare that they have no financial conflicts of interest.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.