
Abstract
Background:
Long noncoding RNAs (lncRNAs) have been reported to be important regulators in cancer. In this study, we aimed to discover the functions of lncRNA TP53TG1 in glioma.
Methods:
The expression of lncRNA TP53TG1, microRNA-524-5p (miR-524-5p) and RAB5A, a member RAS oncogene family (RAB5A), were examined by quantitative real-time polymerase chain reaction. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay and flow cytometry were applied to analyze the proliferation and apoptosis of glioma cells. Colony formation assay was used to detect the colony formation ability and radioresistance of glioma cells. Western blot assay was performed to detect the expression of autophagy-associated proteins and RAB5A. StarBase software was utilized to predict the combination between miR-524-5p and TP53TG1 or RAB5A, and dual-luciferase reporter assay and RNA immunoprecipitation assay were used to verify the above predictions. Animal experiment using immunodeficient nude mice was conducted to detect the role of TP53TG1 in vivo.
Results:
Radiation stimulation (6 Gy) upregulated the abundance of TP53TG1. TP53TG1 potentiated radioresistance and progression of glioma by promoting the autophagy. miR-524-5p was verified as a direct downstream regulation of TP53TG1. miR-524-5p depletion attenuated the influence of TP53TG1 interference on the functions of glioma cells. RAB5A was a direct target of miR-524-5p as well. The inhibitory effect of miR-524-5p on the malignancy of glioma cells was overturned by overexpression of RAB5A. RAB5A was regulated by TP53TG1/miR-524-5p signaling in glioma cells. TP53TG1 silencing impeded the progression of glioma in vivo.
Conclusion:
lncRNA TP53TG1 accelerated the proliferation, colony formation, autophagy, and radioresistance, and restrained the apoptosis of glioma cells through miR-524-5p/RAB5A axis.
Introduction
Glioma accounts for ∼44% of all central nervous system tumors. 1 Among glioma patients, glioblastoma (GBM) patients exhibit the worst prognosis. 2 Radiotherapy is the main complementary therapy for glioma patients having undergone surgery. 3,4 However, the radioresistance of glioma cells limited the therapeutic effect of radiotherapy. 5,6 Therefore, it is urgent to find crucial molecules that regulate the radioresistance of glioma cells.
Long noncoding RNAs (lncRNAs) were regulators of gene expression in the progression of cancers. 7 lncRNAs could serve as microRNAs (miRNAs) sponges to function. 8 –12 lncRNA TP53TG1 was associated with autophagy of glioma cells. 13 Besides, Chen et al. reported that lncRNA TP53TG1 was abnormally upregulated in glioma tissues and cells, and TP53TG1 could accelerate the proliferation and metastasis of glioma cells under glucose deprivation. 14 Kabacik et al. proved that radiation exposure could elevate the expression of lncRNA TP53TG1. 15 However, the correlation among TP53TG1, autophagy, and radioresistance in glioma has never been reported.
Emerging articles have reported that miRNAs were involved in the occurrence and progression of cancers. 16 –18 Xu et al. demonstrated that miR-524 blocked the development of glioma through NCF2. 19 Nevertheless, the regulatory mechanism between miR-524 and the radioresistance of glioma cells remains to be elucidated.
RAB5A, member RAS oncogene family (RAB5A), is a member of Rab GTPase family, which is involved in the regulation of cellular biological functions including vesicle transport. 20,21 RAB5A has emerged as an important regulator in multiple cancers. For instance, Zhang et al. reported that RAB5A facilitated the metastasis of oral cancer cells through ERK/MMP pathway. 22 Yu et al. proved that the abundance of RAB5A was negatively related to the prognosis of patients with colorectal cancer. 23 Geng et al. found that RAB5A accelerated the proliferation and motility of hepatocellular carcinoma cells through FAK pathway. 24 Few studies focused on the role of RAB5A in glioma. Fu et al. found that Malat1 promoted the autophagy and proliferation of glioma cells through upregulating STMN1, RAB5A, and ATG4D by sponging miR-101. 25 Herein, we aimed to illustrate the molecular mechanism behind RAB5A in the progression of glioma, and the interaction between miR-524-5p and TP53TG1 or RAB5A was explored in the following experiments.
Materials and Methods
Patients
Normal brain tissues (n = 51) and glioma tissues (n = 51) were collected from volunteers and glioma patients in Wuhan Puren Hospital. The tissues were obtained during surgery and were utilized to examine the enrichment of TP53TG1, miR-524-5p, and RAB5A. Written informed consent was provided from each participant before surgery. This study had obtained the permission of the Medical Ethics Committee of Wuhan Puren Hospital. The correlation between the expression of TP53TG1, miR-524-5p, and RAB5A, and clinical characteristics of glioma patients are given in Tables 1 –3, respectively.
Correlation Between TP53TG1 Expression and Clinical Characteristics in Glioma Patients (n = 51)
Chi-square test.
p < 0.05.
Correlation Between miR-524-5p Expression and Clinical Characteristics in Glioma Patients (n = 51)
Chi-square test.
p < 0.05.
Correlation Between RAB5A Expression and Clinical Characteristics in Glioma Patients (n = 51)
Chi-square test.
p < 0.05.
Cell culture and radiation
Normal human astrocytes NHA and two glioma cell lines, including LN229 and T98G were purchased from BeNa Culture Collection (Beijing, China). Cells were grown in Dulbecco's modified Eagle medium (Gibco, Carlsbad, CA) added with 10% fetal bovine serum (Gibco), 10% penicillin (100 U/mL), 10% streptomycin (100 μg/mL), and glutamine (2 mM) in a humidified incubator at 37°C and 5% CO2.
Glioma cells were exposed to different doses of X-ray with a linear accelerator (RadSource, Suwanee, GA) at a dose rate of 3.2 Gy/min.
Quantitative real-time polymerase chain reaction
Total RNA was isolated from tissues or cells with TRIzol reagent (Invitrogen, Carlsbad, CA). The reverse transcription of lncRNA TP53TG1, RAB5A, and miR-524-5p was conducted using M-MLV Reverse Transcriptase (Promega, Madison, WI) and One-step miRNA RT Kit (Haigene, Harbin, China). Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using SYBR-Green Master Mix (Takara, Osaka, Japan) and the following primers: lncRNA TP53TG1 (forward, 5′-ACGAAGGTACCCAACCCTCT-3′; reverse, 5′-GGTGTAAGTGTTCGCCTGGT-3′), miR-524-5p (forward, 5′-AAGGGAAGCACTTTCTCTTGTC-3′; reverse, 5′-TCAAACCGTAACACTCCAAAGG-3′), RAB5A (forward, 5′-CAAGAACGATACCATAGCCTAGCAC-3′; reverse, 5′-CTTGCCTCTGAAGTTCTTTAACCC-3′), U6 (forward, 5′-GCACCTTAGGCTGAACA-3′; reverse, 5′-AGCTTATGCCGAGCTCTTGT-3′), and β-actin (forward, 5′-AGCCTCGCCTTTGCCGA-3′; reverse, 5′-CTGGTGCCTGGGGCG-3′). β-actin and U6 served as internal references in this study. The relative expression was analyzed using 2−ΔΔCt method.
Cell transfection
The knockdown of TP53TG1 was conducted with TP53TG1-specific small interfering RNA (si-TP53TG1) or short hairpin RNA (sh-TP53TG1), and si-negative control (si-NC) and sh-NC were used as controls. The overexpression and knockdown of miR-524-5p were conducted using miR-524-5p mimic (miR-524-5p) and miR-524-5p inhibitor (anti-miR-524-5p), and miR-NC and anti-miR-NC were used as controls. RAB5A overexpression was performed using RAB5A overexpression plasmid (RAB5A), and vector was used as the control. Transfection was conducted using Lipofectamine 3000 (Invitrogen).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
Glioma cells were seeded into 96-well plates. After transfection for 0, 24, 48 or 72 h, 20 μL 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sangon, Shanghai, China) was added to the wells of 96-well plates, and glioma cells were incubated with MTT reagent for 4 h in the dark. About 150 μL dimethyl sulfoxide (Invitrogen) was added to dissolve the reaction products for 15 min after discarding the supernatant. Microplate reader (Bio-Rad, Hercules, CA) was used to measure the absorbance (490 nm).
Colony formation assay
For colony formation capacity detection, glioma cells were plated in six-well plates in triplicate (400 cells/well). The cells were incubated for 2 weeks, and the culture media was refreshed per 3 d. The colonies were stained with crystal violet. The colonies including 50 cells or more cells were counted.
For radioresistance detection, transfected glioma cells were seeded into six-well plates in triplicate (400 cells/well), and the cells were exposed to indicated doses using a linear accelerator (RadSource). The other procedures were same as described previously.
Flow cytometry for apoptosis detection
The apoptosis of glioma cells was detected by Annexin V apoptosis detection kit (R&D Systems, Abingdon, United Kingdom). After transfection for 72 h, glioma cells were resuspended in binding buffer, and then the cells were stained using fluorescein isothiocyanate-combined Annexin V and propidine iodide. The apoptosis rate was analyzed on FC-500 flow cytometer (Beckman Coulter, Pasadena, CA) within 1 h of cell harvest.
Western blot assay
Glioma cells were washed and harvested with cold phosphate-buffered saline. Proteins were extracted using radioimmunoprecipitation lysis buffer (Beyotime, Shanghai, China) added with protease inhibitor. The denaturation of protein samples was conducted at 95°C for 5 min. Equal amounts of protein samples were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel and transferred to polyvinylidene fluoride membrane. After being blocked, the membrane was incubated with the following primary antibodies overnight with gentle shaking at 4°C: anti-microtubule-associated protein 1 light chain 3-II/I (anti-LC3-II/I; ab51520; Abcam, Cambridge, MA), anti-p62 (ab155686; Abcam), anti-Beclin-1 (ab210498; Abcam), anti-RAB5A (ab18211; Abcam), and anti-glyceraldehyde-3-phosphate dehydrogenase (anti-GAPDH, ab181602; Abcam). After washing, the membrane was incubated with secondary antibody (ab205718; Abcam). The protein signal was visualized using enhanced chemiluminescence (Thermo Fisher Scientific, Waltham, MA).
Dual-luciferase reporter assay
Luciferase reporter vectors, termed TP53TG1 wild type (TP53TG1 WT), TP53TG1 mutant type (TP53TG1 MUT), RAB5A 3′-untranslated region-WT (RAB5A 3′-UTR-WT), and RAB5A 3′-UTR-MUT, including the wild-type or mutant type binding sequence with miR-524-5p, were constructed. Glioma cells were cotransfected with miR-NC or miR-524-5p and the above reporter vectors for 48 h. The dual-luciferase reporter assay kit (Promega) was used to analyze the luciferase activity.
RNA immunoprecipitation assay
Glioma cells were disrupted using RNA immunoprecipitation (RIP) buffer (Millipore, Billerica, MA). The cell lysate was incubated with sepharose beads precoated with anti-Argonaute-2 (Ago2; Millipore) or anti-immunoglobulin G (Millipore). The immunoprecipitated RNA was examined by qRT-PCR.
Murine xenograft assay
Nine immunodeficient nude mice were purchased from Beijing Laboratory Animal Center (Beijing, China). This study was permitted by the Research Ethics Committee of Wuhan Puren Hospital. About 2 × 106 LN229 cells stably expressing sh-NC or sh-TP53TG1 were subcutaneously injected to the mice (n = 3). When the volume of tumors reached ∼300 mm3, nude mice in sh-NC + radiation group and sh-TP53TG1 + radiation group were irradiated with a dose of 10 Gy. The size of tumors was measured using vernier caliper with the formula of volume = length × width 2 × π/6. The weight of tumors was detected at 5 weeks after inoculation. The expression of TP53TG1, miR-524-5p, and RAB5A was examined using qRT-PCR and Western blot assay.
Statistical analysis
Data were analyzed with GraphPad Prism 7 software and displayed as mean ± standard deviation. Student's t-test was used to compare the differences between two groups, and the differences among multiple groups were analyzed using one-way analysis of variance followed by Tukey's test. A value of p < 0.05 indicated a significant comparison.
Results
lncRNA TP53TG1 is aberrantly upregulated in glioma tissues and cells
An abnormal upregulation in the expression of TP53TG1 was found in glioma tissues and cells (LN229 and T98G) relative to corresponding nontumor tissues and normal human astrocytes NHA (Fig. 1A, B). As mentioned in Figure 1C and D, radiation treatment increased the level of TP53TG1 in glioma cells in a dose-dependent manner. Collectively, TP53TG1 might play a crucial role in the progression and radioresistance of glioma cells.

lncRNA TP53TG1 is aberrantly upregulated in glioma tissues and cells.
TP53TG1 plays an oncogenic role in glioma
We conducted loss-of-function experiments using TP53TG1 si-RNA to explore the biological functions of TP53TG1 in glioma. Before the functional experiments, we initially assessed the knockdown efficiency of si-TP53TG1 in glioma cells. As given in Figure 2A and B, there was a conspicuous decrease in the enrichment of TP53TG1 in si-TP53TG1 group in glioma cells. The optical density value at 490 nm was lower in glioma cells with the transfection of si-TP53TG1 (Fig. 2C, D), suggesting that TP53TG1 intervention prevented the proliferation of glioma cells. The colonies were lesser in si-TP53TG1 group than those in the si-NC group (Fig. 2E, F), demonstrating that TP53TG1 interference inhibited the colony formation ability of glioma cells. The results of flow cytometry showed that the transfection of si-TP53TG1 promoted the apoptosis of glioma cells (Fig. 2G, H). The levels of LC3-II/I and Beclin-1 were decreased in si-TP53TG1 group in comparison with those in si-NC group, and the expression of p62 showed a reverse phenomenon to LC3-II/I or Beclin-1 (Fig. 2I, J). To detect the influence of TP53TG1 depletion on the radioresistance of glioma cells, glioma cells transfected with si-NC or si-TP53TG1 were exposed to different doses of radiation. As indicated in Figure 2K and L, TP53TG1 interference downregulated the survival fraction of glioma cells exposed to radiation. Taken together, TP53TG1 accelerated the radioresistance of glioma cells by promoting the autophagy, and it also promoted the proliferation and colony formation and blocked the apoptosis of glioma cells.
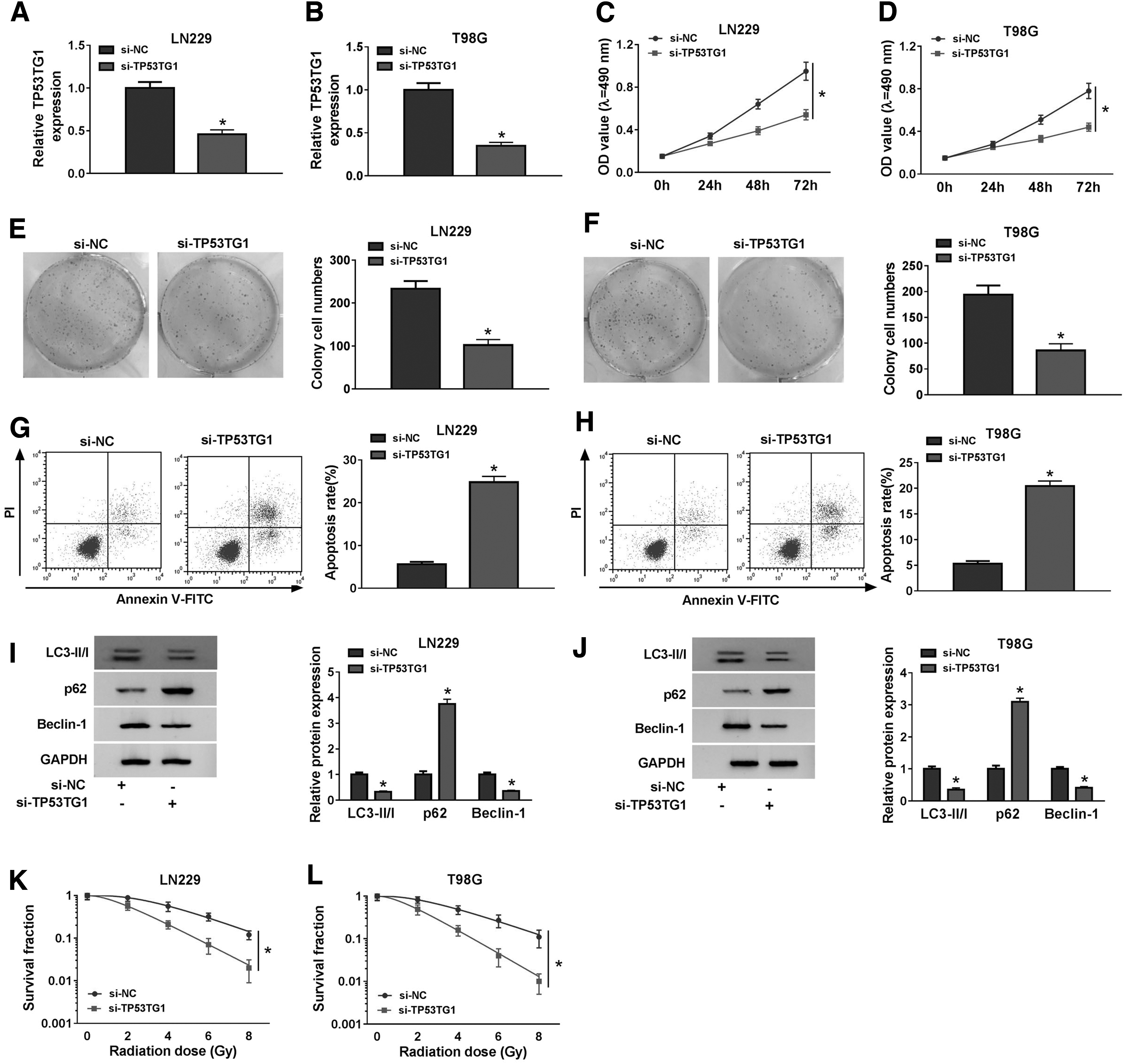
TP53TG1 plays an oncogenic role in glioma. Glioma cells were transfected with si-NC or si-TP53TG1.
miR-524-5p is a direct target of TP53TG1 in glioma cells
To investigate whether TP53TG1 served as miRNA sponge to function, we used StarBase database to explore TP53TG1-miRNA interactions. miR-524-5p was predicted as one of the targets of TP53TG1 (Fig. 3A). The transfection of miR-524-5p could conspicuously decrease the luciferase activity in TP53TG1 WT group relative to miR-NC and TP53TG1 WT group, and the transfection with miR-NC or miR-524-5p did not affect the luciferase activity in TP53TG1 MUT group (Fig. 3B, C), demonstrating that miR-524-5p could directly bind to TP53TG1 in glioma cells. To further verify the interaction, RIP assay was carried out. As mentioned in Figure 3D, TP53TG1 and miR-524-5p were both enriched in RNA-induced silencing complex, which was pulled-down using Ago2 antibody. The regulatory relationship between miR-524-5p and TP53TG1 in glioma cells was then studied. As given in Figure 3E and F, miR-524-5p was significantly upregulated in glioma cells with the transfection of si-TP53TG1. The abundance of miR-524-5p was notably reduced in glioma tissues relative to paired normal tissues (Fig. 3G). Meanwhile, the expression of miR-524-5p was also decreased in glioma cells in contrast to that in NHA cells (Fig. 3H). The radiation of 6 Gy notably reduced the level of miR-524-5p in glioma cells (Fig. 3I, J). In summary, TP53TG1 directly bound to miR-524-5p, and it negatively modulated the level of miR-524-5p in glioma cells.
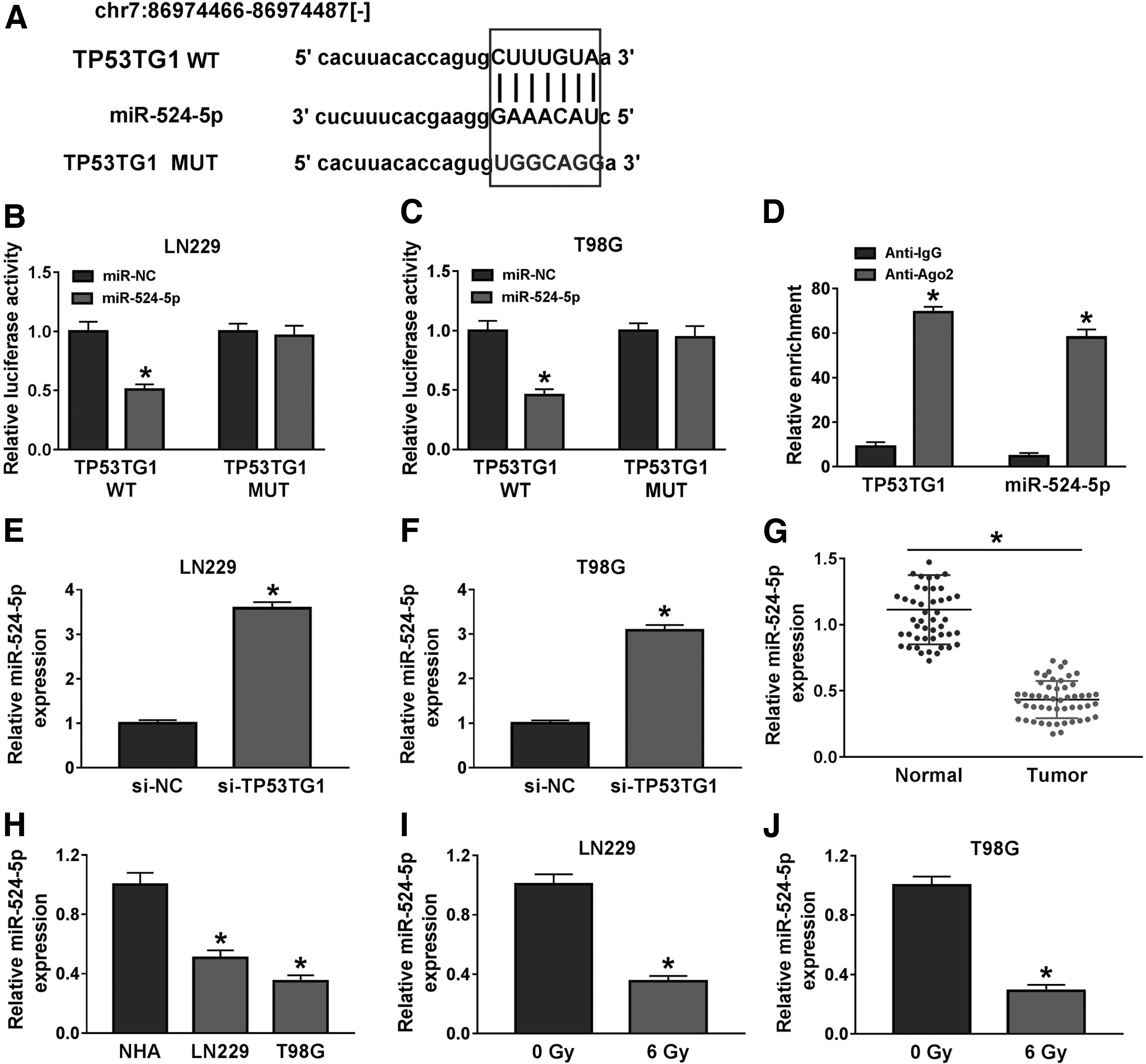
miR-524-5p is a direct target of TP53TG1 in glioma cells.
TP53TG1 contributes to the radioresistance of glioma cells through promoting the autophagy by sponging miR-524-5p
The enrichment of miR-524-5p was elevated with the depletion of TP53TG1, and the addition of anti-miR-524-5p decreased the expression of miR-524-5p in glioma cells (Fig. 4A, B). As exhibited in Figure 4C–H, the transfection of anti-miR-524-5p diminished the inhibitory effects aroused by TP53TG1 intervention on the proliferation and colony formation and the promoting influence on the apoptosis of glioma cells. Furthermore, TP53TG1 interference-mediated autophagy inhibition was overturned by the cotransfection of si-TP53TG1 and anti-miR-524-5p in glioma cells (Fig. 4I, J). In addition, the survival fraction of glioma cells was recovered in si-TP53TG1 and anti-miR-524-5p cotransfected group, which was hampered by the silencing of TP53TG1 (Fig. 4K, L). Taken together, TP53TG1 facilitated the malignancy of glioma through sponging miR-524-5p.
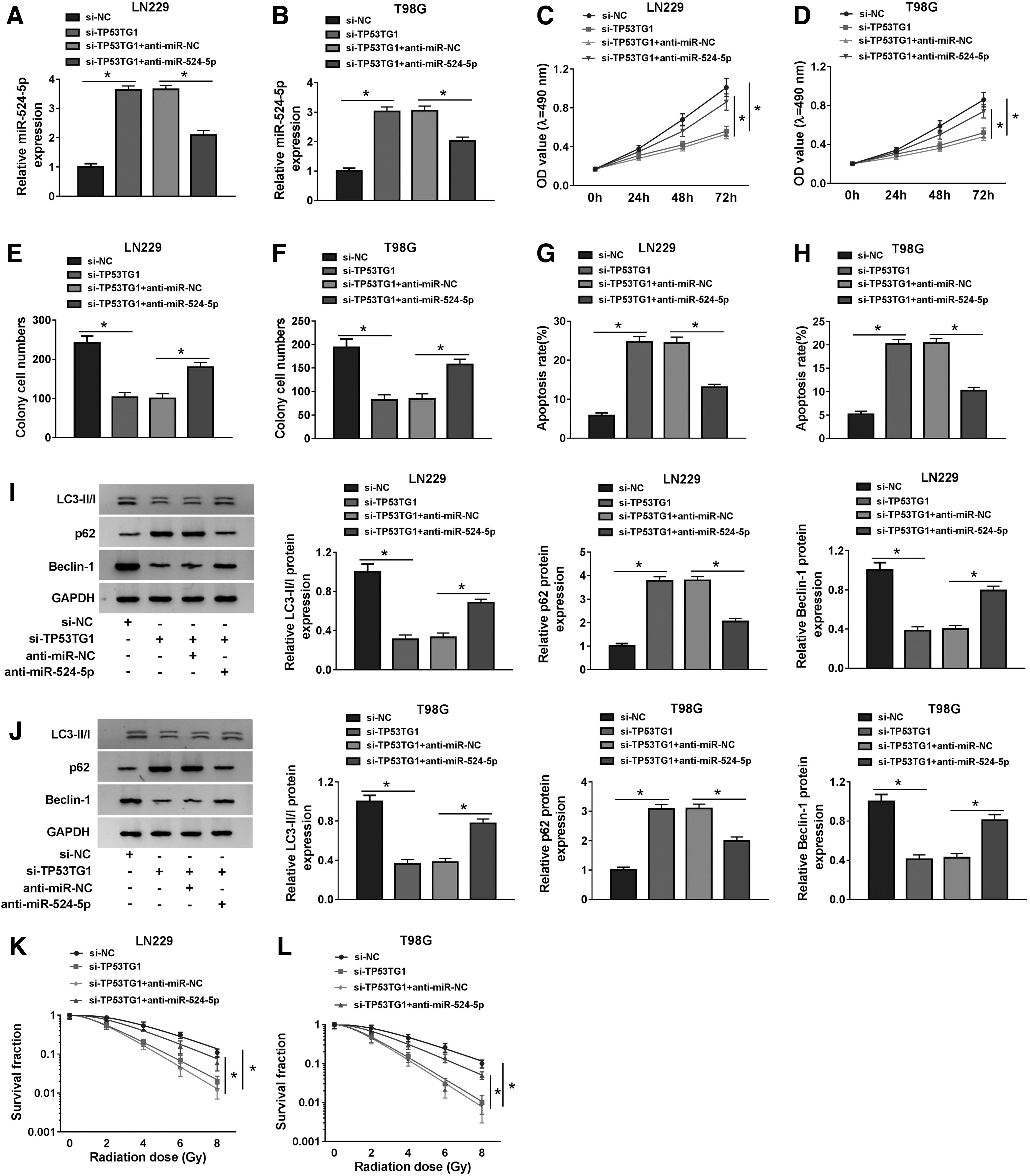
TP53TG1 contributes to the radioresistance of glioma cells through promoting the autophagy by sponging miR-524-5p. We transfected si-NC, si-TP53TG1, si-TP53TG1 + anti-miR-NC or si-TP53TG1 + anti-miR-524-5p into glioma cells.
RAB5A is a direct functional target of miR-524-5p in glioma cells
StarBase predicted that RAB5A possessed the binding sites with miR-524-5p (Fig. 5A). The relative luciferase activity was prominently reduced in miR-524-5p and RAB5A 3′-UTR-WT cotransfected group other than miR-NC and RAB5A 3′-UTR-WT group (Fig. 5B, C). There was a notable reduction in the mRNA and protein levels of RAB5A in glioma cells transfected with miR-524-5p compared with that in miR-NC group (Fig. 5D–G). The expression pattern of RAB5A was explored in glioma tissues and cells. As given in Figure 5H and I, RAB5A mRNA and protein were highly expressed in glioma tissues in comparison with that in matching nontumor tissues. As exhibited in Figure 5J and K, a significant increase in the mRNA and protein expression of RAB5A was observed in the two glioma cells in contrast to that in NHA cells. The high expression of RAB5A in glioma tissues and cells might suggest its pivotal role in the progression of glioma. Moreover, we also explored the influence of 6 Gy radiation on the expression of RAB5A in glioma cells. The mRNA and protein expression of RAB5A was both significantly enhanced in 6 Gy-exposed glioma cells (Fig. 5L–O). In summary, RAB5A was a target of miR-524-5p in glioma cells, and the aberrant expression of RAB5A might imply its important role in the progression and radioresistance of glioma.

RAB5A is a direct functional target of miR-524-5p in glioma cells.
miR-524-5p suppresses the malignancy of glioma cells through suppressing RAB5A
We cotransfected miR-524-5p and RAB5A into LN229 and T98G cells to elucidate the influence of miR-524-5p/RAB5A axis on the functions of glioma cells. The abundance of RAB5A mRNA and protein was prominently decreased with the transfection of miR-524-5p, and the addition of RAB5A ectopic expression plasmid regained the mRNA and protein levels of RAB5A in glioma cells (Fig. 6A–D). The inhibitory effects caused by miR-524-5p accumulation on the proliferation and colony formation were attenuated by the overexpression of RAB5A in glioma cells (Fig. 6E, H). As mentioned in Figure 6I and J, the overexpression of miR-524-5p elevated the apoptosis rate of glioma cells, and the addition of RAB5A counteracted the promoting effect aroused by miR-524-5p overexpression. We also explored the relationship among miR-524-5p/RAB5A axis, autophagy, and radioresistance in glioma cells. As given in Figure 6K–N, the autophagy and survival fraction of glioma cells were prevented by the transfection of miR-524-5p, and the introduction of RAB5A increased the autophagy and survival fraction of glioma cells. In summary, miR-524-5p increased the radiosensitivity of glioma cells through suppressing autophagy by directly inhibiting RAB5A.
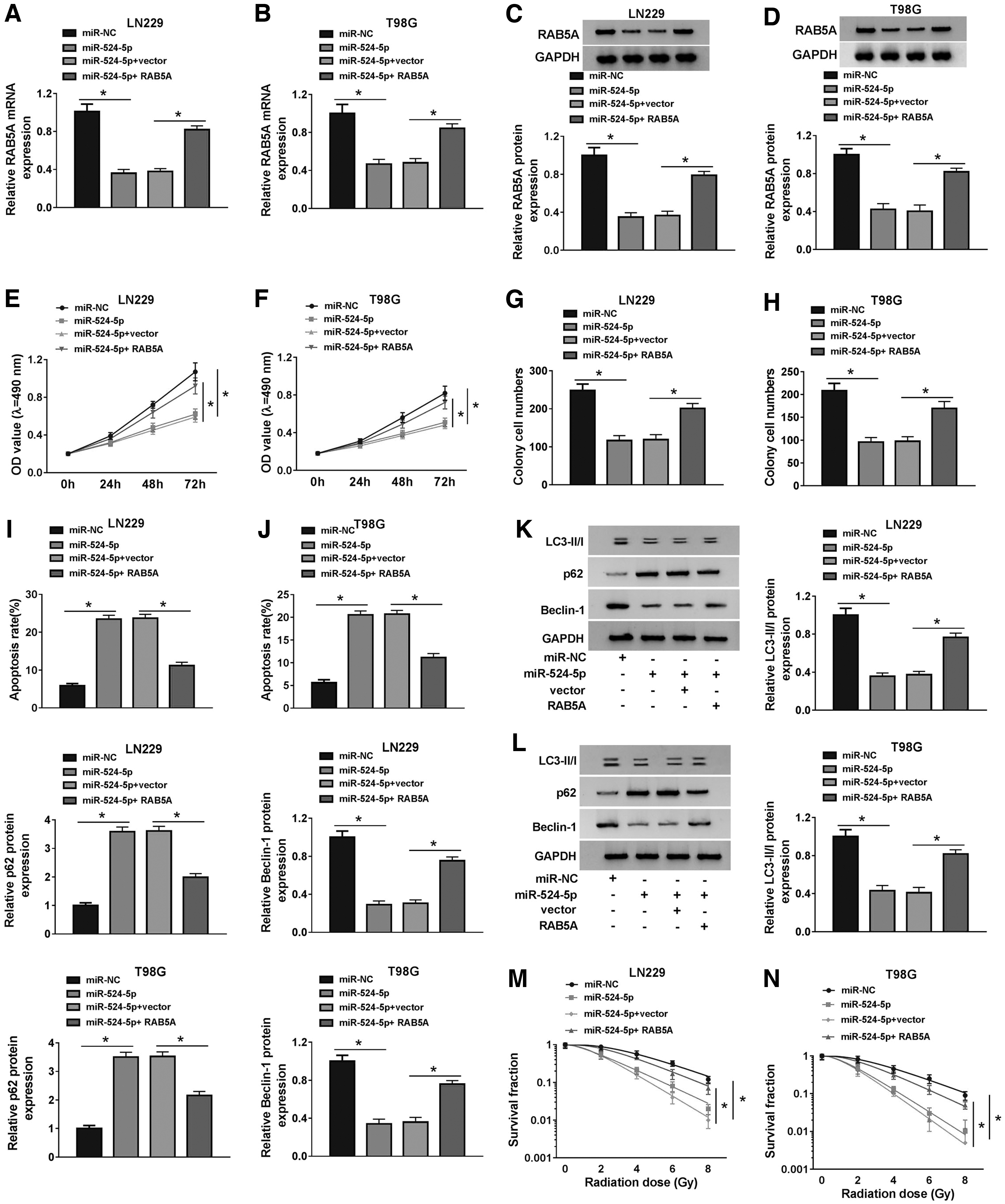
miR-524-5p suppresses the malignancy of glioma cells through suppressing RAB5A. LN229 and T98G cells were transfected with miR-NC, miR-524-5p, miR-524-5p + vector, or miR-524-5p + RAB5A.
TP53TG1 could enhance the expression of RAB5A through serving as a sponge for miR-524-5p in glioma cells
Based on the above findings, we proposed the hypothesis that TP53TG1 exerted its oncogenic role in glioma through regulating miR-524-5p/RAB5A signaling. To validate our hypothesis, LN229 and T98G cells were cotransfected with si-TP53TG1 and anti-miR-524-5p. As given in Figure 7A–D, TP53TG1 knockdown downregulated the mRNA and protein levels of RAB5A, and the abundance of RAB5A mRNA and protein was recovered with the addition of anti-miR-524-5p. Taken together, TP53TG1 upregulated RAB5A through sponging miR-524-5p in glioma cells.
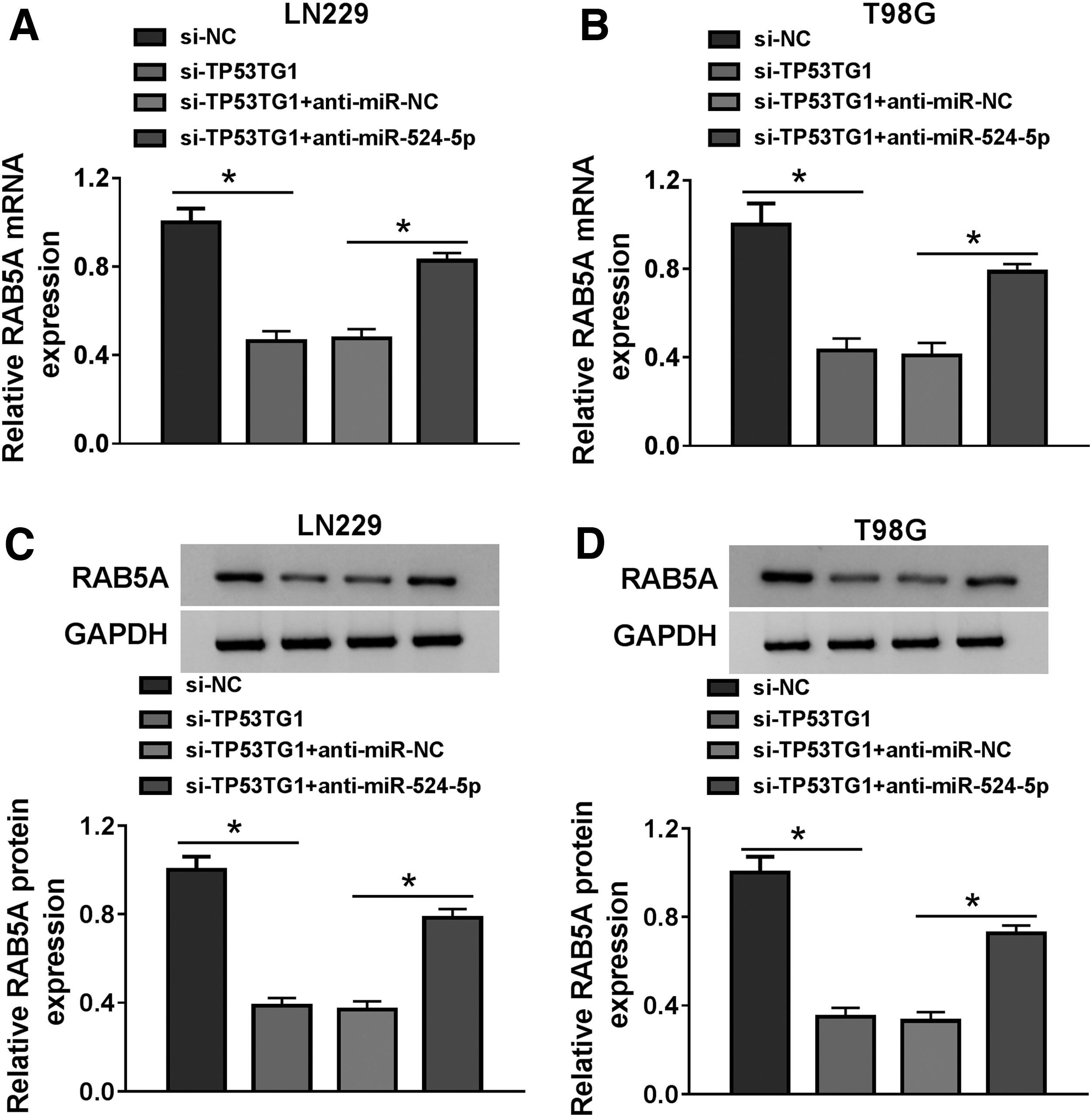
TP53TG1 could enhance the expression of RAB5A through serving as a sponge for miR-524-5p in glioma cells.
TP53TG1 depletion potentiates the radiosensitivity of glioma cells by miR-524-5p/RAB5A in vivo
The role of TP53TG1/miR-524-5p/RAB5A axis was also investigated in vivo. Nude mice were divided into three groups: sh-NC, sh-NC + radiation, and sh-TP53TG1 + radiation. LN229 cells expressing sh-NC or sh-TP53TG1 were injected into immunodeficient nude mice to form xenograft tumors. The size of tumors and the crucial molecules in tumor tissues were measured. As given in Figure 8A, the volume of glioma tumors was significantly reduced in sh-NC + radiation group compared with that in sh-NC group, and TP53TG1 depletion further inhibited the growth of glioma tumors compared with that in sh-NC + radiation group. Meanwhile, the weight of glioma tumors dissected from mice at 5 weeks after injection was declined in sh-NC + radiation group than that in sh-NC group, and the silencing of TP53TG1 enhanced radiation-mediated growth inhibition of glioma tumors (Fig. 8B). The expression of TP53TG1 and RAB5A mRNA and protein was decreased and the enrichment of miR-524-5p was increased in sh-TP53TG1 + radiation group relative to sh-NC + radiation group (Fig. 8C, D). In summary, TP53TG1 depletion contributed to radiation-mediated growth inhibition of glioma through miR-524-5p/RAB5A axis in vivo.
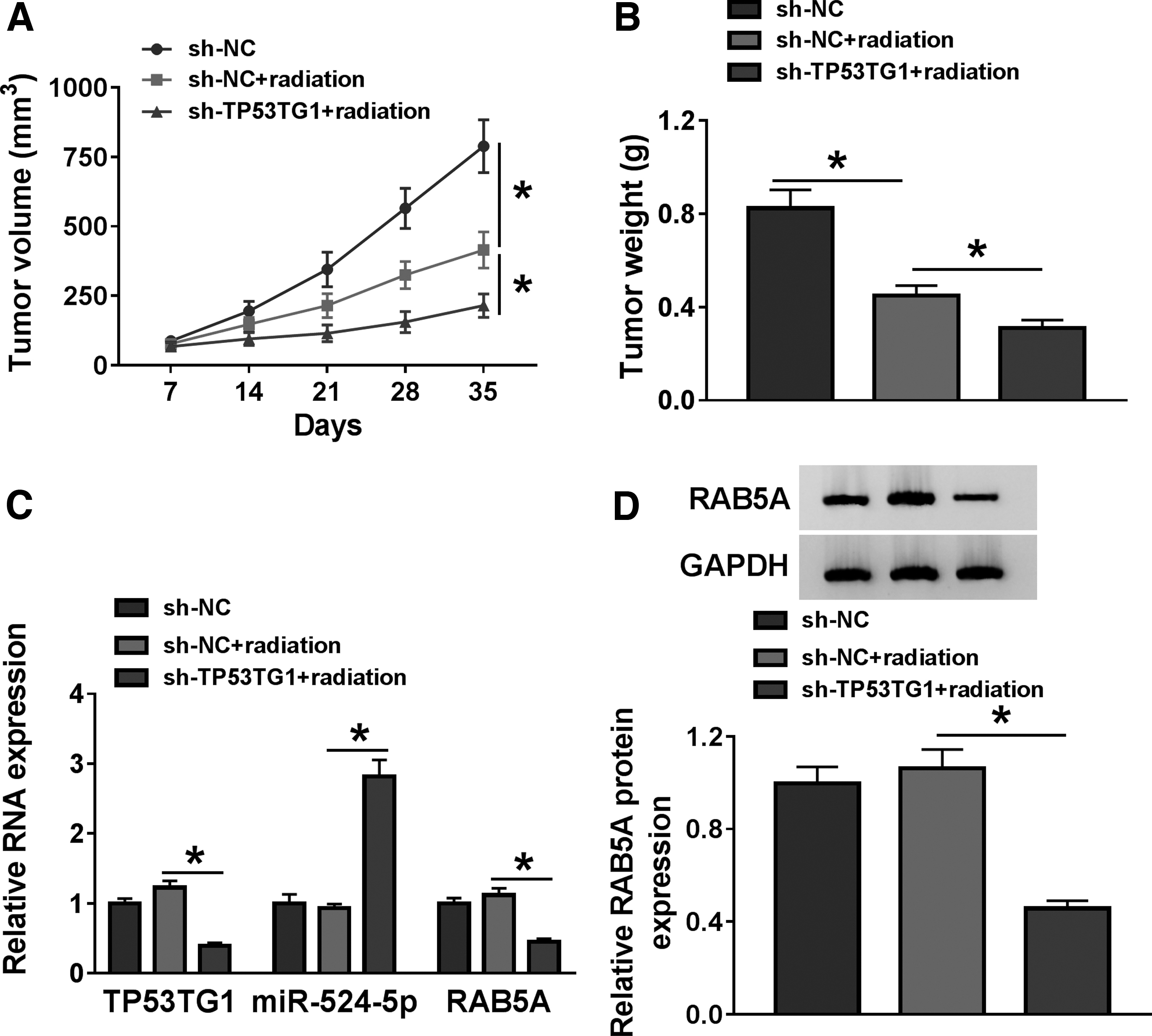
TP53TG1 depletion potentiates the radiosensitivity of glioma cells via miR-524-5p/RAB5A in vivo.
Discussion
lncRNAs play important roles in the initiation and progression of glioma. 26 Cui et al. proved that lncRNA CCAT1 elevated the malignancy of glioma cells through directly suppressing miR-181b. 27 Fu et al. reported that lncRNA PVT1 promoted the progression of glioma. 28 lncRNA TP53TG1 was significantly upregulated in glioma. Besides, a notable increase in the expression of TP53TG1 was found in X-ray-exposed glioma cells in a dose-dependent manner. The abnormal expression of TP53TG1 after radiation hinted its crucial role in radioresistance of glioma cells. Autophagy is a conserved cellular process that helps cells to maintain homeostasis through degrading excess cellular components. Autophagy was found to be overactivated in solid tumors because of the absence of oxygen and nutrients. 29 Autophagy was also activated in patients after chemotherapy or radiotherapy. Taylor et al. reported the correlation between autophagy and radioresistance or chemoresistance in GBM. 30 However, the role of autophagy in the radioresistance of cancer cells is controversial. 30 –34 For instance, Comincini et al. claimed that autophagy activation elevated the radiosensitivity of GBM cells. 34 Ye et al. demonstrated that the activation of autophagy potentiated the radioresistance of glioma cells. 35 Loss-of-function experiments revealed that TP53TG1 played an oncogenic role in glioma by accelerating the radioresistance through promoting autophagy, and TP53TG1 also facilitated the proliferation and colony formation and impeded the apoptosis of glioma cells. The protumor role of TP53TG1 was alignment with former articles related to glioma and pancreatic ductal adenocarcinoma. 14,36
To illustrate the potential mechanism behind TP53TG1 in the progression of glioma, we intended to seek the downstream genes of TP53TG1. miR-524-5p has been reported to be a tumor suppressor in various cancers. For example, Liu et al. claimed that miR-524-5p hampered the progression of gastric cancer. 37 Liu et al. demonstrated that miR-524-5p restrained the development of papillary thyroid carcinoma. 38 Xu et al. claimed that miR-524 suppressed the proliferation and metastasis of glioma cells by NCF2. 19 We found that miR-524-5p was a direct functional target of TP53TG1 in glioma cells. Consistent with the above articles, 19,37,38 we found that miR-524-5p served as the downstream target of TP53TG1 to repress the malignancy of glioma cells. Besides, we found that TP53TG1 facilitated the radioresistance through enhancing the autophagy of glioma cells by sponging miR-524-5p.
RAB5A was validated as a target of miR-524-5p in glioma cells. RAB5A was involved in the progression of multiple cancers. 39,40 Moreover, it has been reported that RAB5A is associated with autophagy. Ravikumar et al. found that RAB5A promoted the formation of autophagosome, suggesting that RAB5A was an autophagy-related protein. 41 Fu et al. proved that RAB5A, acting as one of the targets of MALAT1/miR-101 axis, promoted MALAT1-mediated autophagy and proliferation of glioma cells. 25 Tan et al. found that RAB5A modulated the phenotype and functions of vascular smooth muscle cells through promoting autophagy. 42 We found that miR-524-5p inhibited the radioresistance of glioma cells through suppressing autophagy by RAB5A. Further studies showed that RAB5A was regulated by TP53TG1/miR-524-5p axis in glioma cells. The role of TP53TG1/miR-524-5p/RAB5A axis was also confirmed in vivo. TP53TG1 accelerated the growth of glioma tumors by elevating the expression of RAB5A through sponging miR-524-5p in vivo.
Nevertheless, more efforts should be conducted to uncover the molecular mechanism by which RAB5A regulates radioresistance and autophagy of glioma cells.
Conclusions
Overall, lncRNA TP53TG1 contributed to the radioresistance through promoting autophagy by miR-524-5p/RAB5A axis, and TP53TG1 also mediated the promotion on the proliferation and colony formation and suppression on the apoptosis of glioma cells. TP53TG1/miR-524-5p/RAB5A axis might provide a new target for overcoming the radioresistance of glioma patients.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.