
Abstract
Background:
Emerging studies manifested that exosomal RNAs had pivotal roles in human cancer therapies. This article aimed to research the regulatory mechanism of exosomal circRNA-plasmacytoma variant translocation 1 (circ-PVT1) in cisplatin (DDP) resistance of gastric cancer (GC).
Methods:
Exosomes were isolated by ExoQuick® method and ultracentrifugation and then identified through transmission electron microscope and the examination of exosome markers. Related proteins were detected using Western blot. Quantitative real-time polymerase chain reaction (qRT-PCR) was applied for measuring circ-PVT1, microRNA-30a-5p (miR-30a-5p), and Yes-associated protein 1 (YAP1) expression. The half inhibitory concentration (IC50) of DDP was assessed by 3-(4, 5-dimethylthiazol-2-y1)-2, 5-diphenyl tetrazolium bromide (MTT). Cell apoptosis and invasion were, respectively, determined using flow cytometry and transwell assay. Target relationship was confirmed by dual-luciferase reporter assay. The impact of circ-PVT1 on DDP resistance was explored via xenograft tumor assay.
Results:
Exosomal circ-PVT1 was upregulated while miR-30a-5p was downregulated in DDP-resistant GC serums and cells. Circ-PVT1 knockdown repressed DDP resistance in DDP-resistant GC cells via promoting apoptosis and decreasing invasion or autophagy by negatively targeting miR-30a-5p. YAP1 was a direct target of miR-30a-5p. MiR-30a-5p overexpression inhibited DDP resistance via reducing YAP1. Circ-PVT1 modulated YAP1 expression by targeting miR-30a-5p. Circ-PVT1 depression expedited DDP sensitivity of GC via miR-30a-5p/YAP1 axis in vivo.
Conclusion:
Exosomal circ-PVT1 facilitated DDP resistance via modulating autophagy, invasion and apoptosis by miR-30a-5p/YAP1 axis in GC cells. Exosomal circ-PVT1 might be a prospective indicator in DDP therapy of GC.
Highlights
Exosomal circ-PVT1 is upregulated while miR-30a-5p is downregulated in cisplatin-resistant GC serums and cells
Knockdown of circ-PVT1 suppresses cisplatin resistance in cisplatin-resistant GC cells via inducing apoptosis and inhibiting invasion or autophagy by negatively targeting miR-30a-5p
MiR-30a-5p directly interacts with YAP1, and circ-PVT1 regulates YAP1 via targeting miR-30a-5p
Circ-PVT1 depression enhances cisplatin sensitivity of GC by miR-30a-5p/YAP1 in vivo
Introduction
Gastric cancer (GC) is the fifth leading cause of cancer related death according to recent statistics. 1,2 The survival of GC patients has been advanced with radiography, endoscopy, serology, and chemotherapy. 3,4 Cisplatin (DDP) is an effective chemotherapy of GC. 5 However, the efficacy of this therapy is seriously impacted due to acquired resistance. Thus, the authors focused on exploration of the molecular mechanism in the development of GC resistance to DDP.
Exosomes, a class of extracellular vesicles (EVs), are featured as roundness with a diameter of 30–100 nm and lipid bilayer membranes. 6 Exosomes can release cargo biomolecules, including RNAs, proteins, or lipids, into the extracellular environment, 7,8 exerting regulation in different biological processes of recipient cells, which is known as the third intercellular communication. 9,10 Several studies have unveiled that exosomal circular RNAs (circRNAs), belong to noncoding RNAs (ncRNAs), had an active application in human diseases, 11 such as lung adenocarcinoma 12 and hepatocellular carcinoma, 13 and GC. 14 CircRNA-plasmacytoma variant translocation 1 (circ-PVT1) was upregulated in GC tissues, 15 but the biological function of exosomal circ-PVT1 and its functional mechanism in the DDP resistance of GC remain poorly understood.
MiRNAs are another genre of ncRNAs with only 20–25 nucleotides. They play essential regulatory effects on cellular behaviors by decreasing the abundance of target mRNAs. 16,17 MiRNA-544 was reported to promote DDP sensitivity in esophageal squamous cell carcinoma (ESCC) via the inhibition of E2F5. 18 Lu et al. declared that miRNA-424 participated in the regulation of DDP resistance in GC by targeting SMURF1. 19 In this study, the authors intended to investigate the role of miR-30a-5p in DDP resistance of GC.
Yes-associated protein 1 (YAP1) has been considered as one of the most momentous effectors of the Hippo pathway and has crosstalk with tumorigenesis. 20 Mounting articles have clarified the regulatory role of YAP1 in DDP resistance, such as ESCC, 21 breast cancer, 22 laryngeal squamous cell carcinoma, 23 and so on. Li et al. proved that the high expression of YAP1 was associated with poor prognosis of GC. 24 However, the function of YAP1 in DDP resistance in GC is not fully addressed.
This article hammered at the regulatory relationship between circ-PVT1 and miR-30a-5p as well as the combination between miR-30a-5p and YAP1 in GC cells. The significance of exosomal circ-PVT1/miR-30a-5p/YAP1 in DDP resistance of GC was uncovered by in vitro and in vivo assays.
Materials and Methods
Serum collection
Patients with GC (n = 60) were recruited to Shiyan People's Hospital. They were subjected to the drug therapy of DDP and divided into sensitive and resistant groups with 30 per group. Blood samples were collected and centrifuged at 3000 × g for 10 min, and then the supernatant was transferred into RNase-free tubes and stored in ultralow temperature refrigerator of −80°C. All individuals who participated in this study signed informed consent. This article was authorized by the Ethics Committee of Shiyan People's Hospital.
Exosomes isolation and assessment
The serum exosomes were extracted using ExoQuick® ULTRA EV Isolation Kit (System Biosciences, Johnstown, PA) following the producer's instruction. In brief, ExoQuick ULTRA was added to 250 μL serum for 30 min at 4°C. After spinning at 3000 × g for 10 min, the pellets were suspended and added to the prewashed ExoQuick ULTRA columns to spin again at 1000 × g for 30 s. Exosomes were obtained after pellets in the bottom of the tube were resuspended with 50 μL phosphate-buffered saline (PBS; HyClone, Logan, UT). Cell-exosomes were precipitated through ultracentrifugation (UC) as previously reported. 25 Whereafter, the identification of exosomes was implemented via morphology investigation by transmission electron microscope (TEM; JEM-1400, JEOL, Japan) 26 and the assaying of representative exosomes markers (cluster of differentiation 63 (CD63) and CD9) by Western blot.
Western blot
Proteins were extracted by RIPA lysis solution (Beyotime, Shanghai, China). Then, 40 μg proteins were separated through sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto the polyvinylidene fluoride membranes (Millipore, Bedford, MA), followed by blocking using 5% skim milk (Beyotime) for 3 h and incubating with primary antibodies overnight at 4°C. The primary antibodies anti-CD63 (ab217345, 1:1000), anti-CD9 (ab92726, 1:1000), anti-light chain 3B (anti-LC3B; ab51520, 1:1000), anti-P62 (ab109012, 1:1000), anti-P-glycoprotein (anti-P-gp; ab103477, 1:1000), anti-YAP1 (ab52771, 1:1000), and anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Abcam, ab9485, 1:3000) were purchased from Abcam (Cambridge, United Kingdom). Then membranes were incubated with secondary antibody (Abcam, ab205718, 1:5000) for 1 h, and protein signals were examined using enhanced chemiluminescence reagent (Abcam) and analyzed by ImageJ software (NIH, Bethesda, MD).
Quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (qRT-PCR) was administrated using TB Green® Premix Ex Taq™ II Kit (Takara, Dalian, China) in line with the description of previous study. 27 The relative expression levels of circ-PVT1 or YAP1 and miR-30a-5p were standardized by GAPDH and U6 through the 2−ΔΔCt approach. 28 The sequences of primers in this article were as follows: circ-PVT1 (Forward: 5′-ATCGGTGCCTCAGCGTTCGG-3′, Reverse: 5′-CTGTCCTCGCCGTCACACCG-3′); miR-30a-5p (Forward: 5′-CAGTGCAGGGTCCGAGGT-3′, Reverse: 5′-AACACGCTGTAAACATCCTCGAC-3′); YAP1 (Forward: 5′-GCTACAGTGTCCCTCGAACC-3′, Reverse: 5′-TCCTTCCAGTGTTCCAAGGT-3′); GAPDH (Forward: 5′-GGGAGCCAAAAGGGTCATCA-3′, Reverse: 5′-TGATGGCATGGACTGTGGTC-3′); U6 (Forward: 5′-CTCGCTTCGGCAGCACA-3′, Reverse: 5′-AACGCTTCACGAATTTGCGT-3′).
Cell culture and DDP treatment
Human normal gastric epithelial cell line GES-1 and GC cell lines (HGC-27 and a human gastric adenocarcinoma cell line [AGS]) were bought from COBIOER Biosciences Co., Ltd. (Nanjing, China). AGS cells were maintained in F12K medium and other cells were cultivated in Roswell Park Memorial Institute-1640 (RPMI-1640; Invitrogen, Carlsbad, CA) in a 37°C humidified incubator with 5% CO2. All media were added with 10% fetal bovine serum (FBS; Gibco, Carlsbad, CA) and antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin; Gibco). HGC-27 and AGS cells were treated with DDP (DDP; Beyotime) to establish the DDP-resistant cell lines (HGC-27/DDP and AGS/DDP) as previously reported. 29
Cell transfection
Cell transfection was implemented via Lipofectamine3000 reagent (Invitrogen) according to the operating procedures provided by the manufacturer. Small interfering RNA (siRNA) against circ-PVT1 (si-circ-PVT1), short hairpin RNA against circ-PVT1 (sh-circ-PVT1), overexpression vector of YAP1 (YAP1), and respective negative controls (si-NC, sh-NC and pcDNA) were purchased from GenePharma (Shanghai, China). MiR-30a-5p mimic and inhibitor (miR-30a-5p and anti-miR-30a-5p) and matched negative controls (miR-NC and anti-miR-NC) were acquired from RiboBio (Guangzhou, China).
3-(4, 5-dimethylthiazol-2-y1)-2, 5-diphenyl tetrazolium bromide assay
3-(4, 5-dimethylthiazol-2-y1)-2, 5-diphenyl tetrazolium bromide (MTT) assay was applied for determining the half inhibitory concentration (IC50) of DDP. Cells were inoculated into 96-well plates overnight with 2 × 103 cells per well. After treatment with different concentrations of DDP, 20 μL MTT (Beyotime) was pipetted into each well and mixed for another 4 h, then the supernatant was discarded. After the incubation with 200 μL/well dimethyl sulfoxide (DMSO; Beyotime) for 10 min, cell viability was detected by the absorbance at 490 nm via a microplate reader. When cell viability was 50%, the concentration of DDP was considered as the IC50.
Flow cytometry
Cell apoptosis was assessed by Annexin V-fluorescein isothiocyanate (Annexin V-FITC)/propidium iodide (PI) regent kit (BD Biosciences, San Diego, CA). Collected HGC-27/DDP and AGS/DDP cells were washed by precooled PBS (HyClone) and centrifuged at 2000 rpm for 10 min, followed by the resuspension of cell pellet with 300 μL 1 × binding buffer. Five microliter Annexin V-FITC was pipetted into cell suspension to incubate for 15 min away from light. After adding 2.5 μL PI for 5 min, apoptotic cells were identified by a flow cytometer (BD Biosciences).
Transwell invasion assay
The invaded abilities of HGC-27/DDP and AGS/DDP cells were evaluated by transwell assay. At first, Matrigel (BD Biosciences) was used for coating the low side of the upper chamber in transwell chamber (8 mm pores; Costar, Cambridge, MA). Then cells in serum-free medium were seeded into the upper chamber, and the lower chamber was filled with specific medium containing 10% FBS. After 48 h, cells were fastened using methanol (Supelco, Bellefonte, PA) and stained by crystal violet (Supelco). After wiping off uninvaded cells with a wet cotton swab, invaded cells in the inverse side of the membrane were counted under a microscope.
Dual-luciferase reporter assay
StarBase v2.0 was adopted for predicting the targets of circ-PVT1 and miR-30a-5p. The combination between miR-30a-5p and circ-PVT1 or YAP1 was affirmed by dual-luciferase reporter assay. First, the authors constructed luciferase reporters through molecular cloning by the pmirGLO vector (Promega, Madison, WI), including wide-type reporters (WT-circ-PVT1 and YAP1 3′UTR-WT) with the binding sites for miR-30a-5p and mutant-type reporters (MUT-circ-PVT1 and YAP1 3′UTR-MUT) with mutant sites for miR-30a-5p. Then luciferase activities of HGC-27/DDP and AGS/DDP cells cotransfected with above reporters and miR-30a-5p or miR-NC were assayed via dual-luciferase reporter assay system (Promega) following the producer's direction. Firefly activity was normalized by Renilla activity.
Xenograft tumor assay
This xenograft experiment was approved by the Animal Ethics Committee of Shiyan People's Hospital. BALB/c nude mice (6-week-old) were obtained from Shanghai SIPPR-BK Laboratory Animal Co. Ltd. (Shanghai, China). Xenograft tumor model was established using HGC-27/DDP cells stably expressed sh-circ-PVT1 with sh-NC as reference. Transfected cells (2 × 106/0.2 mL PBS) were subcutaneously injected into mice. After 8 d, mice were intraperitoneally injected with 6 mg/kg DDP or PBS every 3 d. Tumor volume was recorded every 3 d according to the formula: length × width 2 × 0.5. The euthanasia was executed to all mice after 29 d, then tumor tissues were excised and weighed. The levels of circ-PVT1, miR-30a-5p, and YAP1 were examined using qRT-PCR or Western blot.
Statistical analysis
All the experiments were performed in three biological repetitions. Data are expressed as mean ± standard deviation and processed by SPSS 19.0 and GraphPad Prism 7. The linear relationship was analyzed using Spearman's correlation coefficient. The difference analysis was administrated through Student's t-test or one-way analysis of variance followed by Tukey's test. p < 0.05 was deemed as statistically significant.
Results
Exosomal circ-PVT1 was upregulated, while miR-30a-5p was downregulated in DDP-resistant GC serums and cells
To explore the package of circ-PVT1 in exosomes, the authors first isolated exosomes from serum specimens of GC patients and cells then performed the identification through the morphology and phenotypes. As shown in Figure 1A, the particles were observed as round-shaped vesicles with bistratal membranes and diameter size about 100 nm under the TEM. Also, the protein levels of CD63 and CD9 were examined via Western blot, in which both two positive markers were detected in the exosomes from GC serums and cells but not the centrifugal supernatant (Fig. 1B). Thus, collected particles in the study were identified as exosomes. In 30 GC tissues, 50% (15/30) of them showed the increased expression level of exosomal circ-PVT1, and the high circ-PVT1 was positively correlated with tumor-node-metas-tasis grade (p = 0.0029), lymph node metastasis (p = 0.0099), and tumor size (p = 0.0281) (Table 1). Subsequently, they measured the circ-PVT1 level in exosomes from DDP-sensitive or resistant GC serums and cells by qRT-PCR. Significantly, circ-PVT1 level in exosomes derived from DDP-resistant GC serums was much higher than that in exosomes from DDP-sensitive serums (Fig. 1C). Also, circ-PVT1 expression was overtly upregulated in exosomes from DDP-resistant GC cells (HGC-27/DDP and AGS/DDP) compared with sensitive cells (HGC-27 and AGS) and normal GES-1 cells (Fig. 1D). Then, they verified the increase of circ-PVT1 in DDP-resistant GC cells contrasted to parental cells and normal cells (Fig. 1E). On the contrary, miR-30a-5p had a lower expression in DDP-resistant GC serums and cells than that in sensitive serum and cells (Fig. 1F, G). Interestingly, there was a conspicuous negative relationship (r = −0.576, p < 0.001) between circ-PVT1 and miR-30a-5p levels in DDP-resistant GC serums (Fig. 1H).
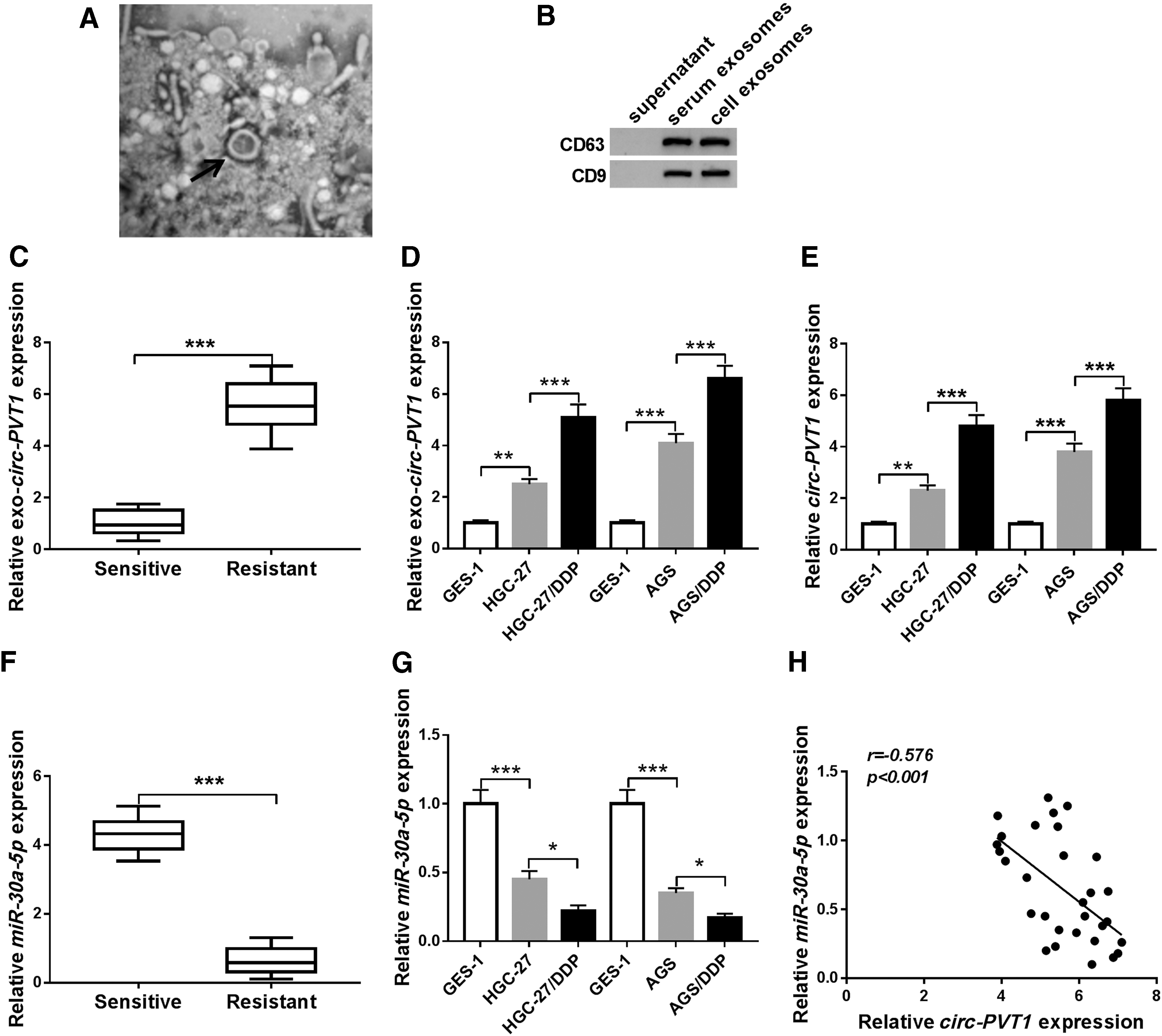
Exosomal circ-PVT1 was upregulated, while miR-30a-5p was downregulated in DDP-resistant GC serums and cells.
Relationship Between Serum Exosomal Circ-PVT1 Expression and Clinicopathologic Features of Gastric Cancer Patients
Chi-square test.
p < 0.05.
circ-PVT1, circRNA-plasmacytoma variant translocation 1; TNM, tumor-node-metas-tasis.
Circ-PVT1 knockdown repressed DDP resistance in DDP-resistant GC cells by inducing apoptosis and inhibiting invasion or autophagy
Next, the authors investigated the role of circ-PVT1 in regulating DDP resistance in GC. Through the analysis of MTT assay, the IC50 of DDP was markedly enhanced in HGC-27/DDP and AGS/DDP cells by comparison with HGC-27 and AGS cells, suggesting the generation of DDP resistance in HGC-27/DDP and AGS/DDP cells (Fig. 2A). The authors used si-circ-PVT1 transfection to interfere the expression of circ-PVT1, and the interference efficiency was distinct in HGC-27/DDP and AGS/DDP cells compared to si-NC group (Fig. 2B). After circ-PVT1 was knocked down, the IC50 of DDP was obviously decreased (Fig. 2C). Besides, transfection of si-circ-PVT1 triggered an enhancement of cell apoptosis (Fig. 2D), but a descent of cell invasion (Fig. 2E). Then, they examined the autophagy-related protein levels, and the results showed that DDP treatment enhanced LC3-II/I level (proautophagy indicator) and inhibited P62 protein expression (antiautophagy marker) as time goes on (Fig. 2F) in HGC-27 and AGS cells, implying that DDP treatment induced autophagy. Moreover, knockdown of circ-PVT1 inhibited LC3-II/I level, but promoted P62 level, and reduced the protein expression of P-gp (proresistance marker) in HGC-27/DDP and AGS/DDP cells (Fig. 2G, H). Taken together, circ-PVT1 knockdown repressed DDP resistance via inducing apoptosis and inhibiting invasion or autophagy in GC cells.
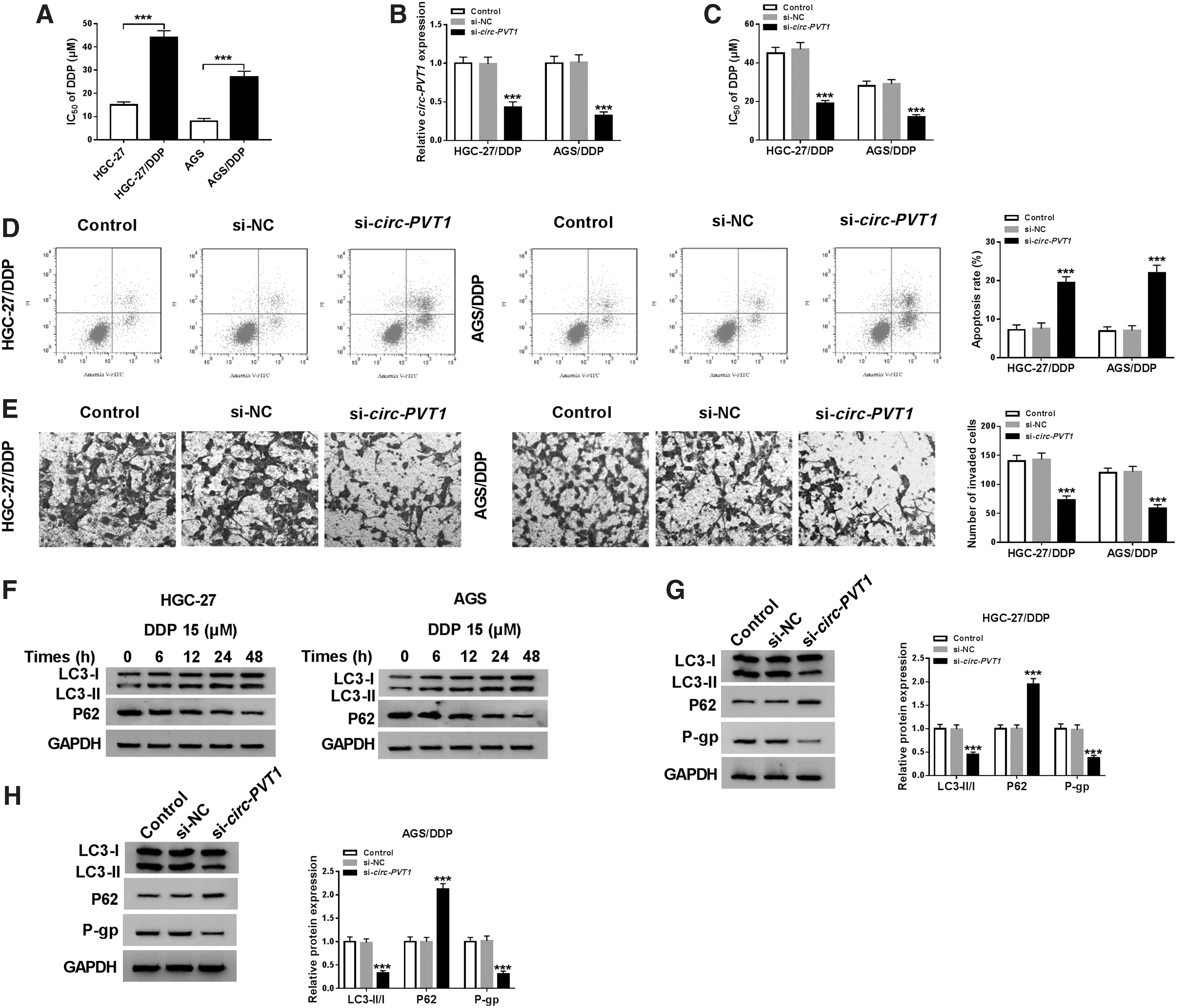
Circ-PVT1 knockdown repressed DDP resistance in DDP-resistant GC cells by inducing apoptosis and inhibiting invasion or autophagy.
The depletion of circ-PVT1 refrained DDP resistance in DDP-resistant GC cells by negatively targeting miR-30a-5p
Then, they found that circ-PVT1 had the complementary sites for miR-30a-5p through StarBase v2.0 (Fig. 3A). Dual-luciferase reporter assay revealed that the luciferase activity in WT-circ-PVT1 group was visibly decreased by miR-30a-5p transfection, but it remained unchanged in MUT-circ-PVT1 group in HGC-27/DDP and AGS/DDP cells (Fig. 3B, C). QRT-PCR analysis indicated the knockdown, and overexpression effects of si-circ-PVT1 and circ-PVT1 were successful (Fig. 3D), then si-circ-PVT1 transfection elevated miR-30a-5p expression, but circ-PVT1 introduction generated an opposite tendency (Fig. 3E). Furthermore, HGC-27/DDP and AGS/DDP cells were transfected with si-circ-PVT1, si-circ-PVT1+anti-miR-30a-5p, or relative controls to analyze the regulatory mechanism between circ-PVT1 and miR-30a-5p. As Figure 3F depicted, transfection of anti-miR-30a-5p clearly recovered the upregulation of miR-30a-5p caused by circ-PVT1 knockdown, and si-circ-PVT1-induced inhibition of DDP resistance was also relieved following downregulation of miR-30a-5p (Fig. 3G). After miR-30a-5p expression was disturbed, there was a mitigation of apoptosis promotion (Fig. 3H) and invasion repression (Fig. 3I) evoked by depression of circ-PVT1. In addition, miR-30a-5p inhibitor rescued the reduction of LC3-II/I and the increase of P62, as well as the inhibition of P-gp in HGC-27/DDP and AGS/DDP cells transfected with si-circ-PVT1 (Fig. 3J, K). Above results demonstrated that circ-PVT1 knockdown refrained DDP resistance by negatively promoting miR-30a-5p.
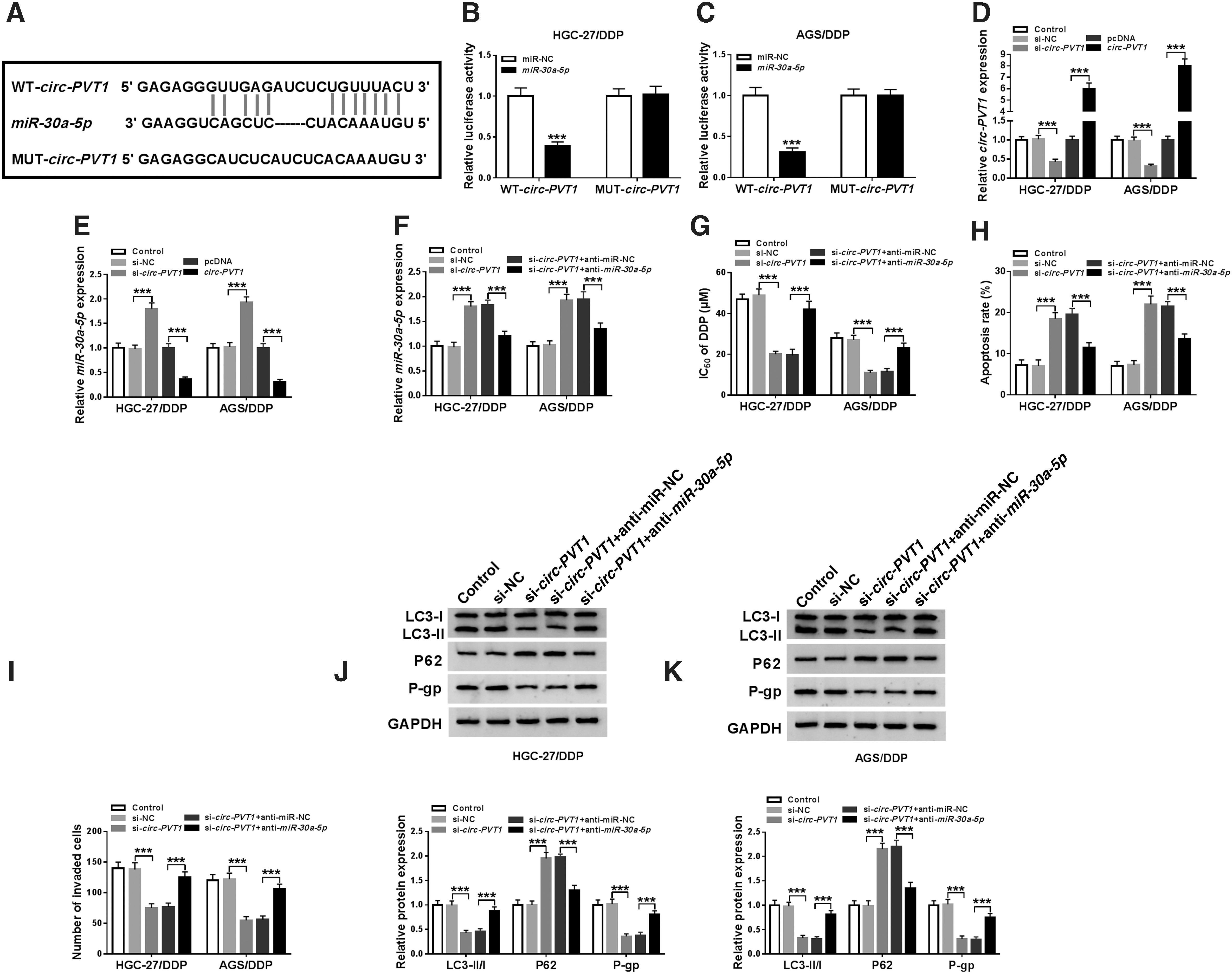
The depletion of circ-PVT1 refrained DDP resistance in DDP-resistant GC cells by negatively targeting miR-30a-5p.
miR-30a-5p directly interacted with YAP1
Whereafter, YAP1 was considered as a candidate target of miR-30a-5p because of the mutual binding sites in StarBase v2.0 (Fig. 4A). The authors conducted dual-luciferase reporter assay to verify whether miR-30a-5p targeted YAP1, and data displayed that miR-30a-5p overexpression reduced the luciferase activity in YAP1 3′UTR-WT group, but failed to descend that in YAP1 3′UTR-MUT group (Fig. 4B, C). The expression of YAP1 was detected using qRT-PCR and Western blot. Remarkably, the mRNA and protein levels were much higher in DDP-resistant GC serums than that in sensitive serums (Fig. 4D, E). Likewise, they found the upregulation of YAP1 mRNA and protein expression in DDP-resistant GC cells by contrast with sensitive GC cells and normal GES-1 cells (Fig. 4F, G) and between miR-30a-5p and YAP1 expression in DDP-resistant GC serums, Spearman's correlation coefficient analysis exhibited a prominent negative correlation (r = −0.6947, p < 0.001) (Fig. 4H). Hence, YAP1 was identified as a target of miR-30a-5p.
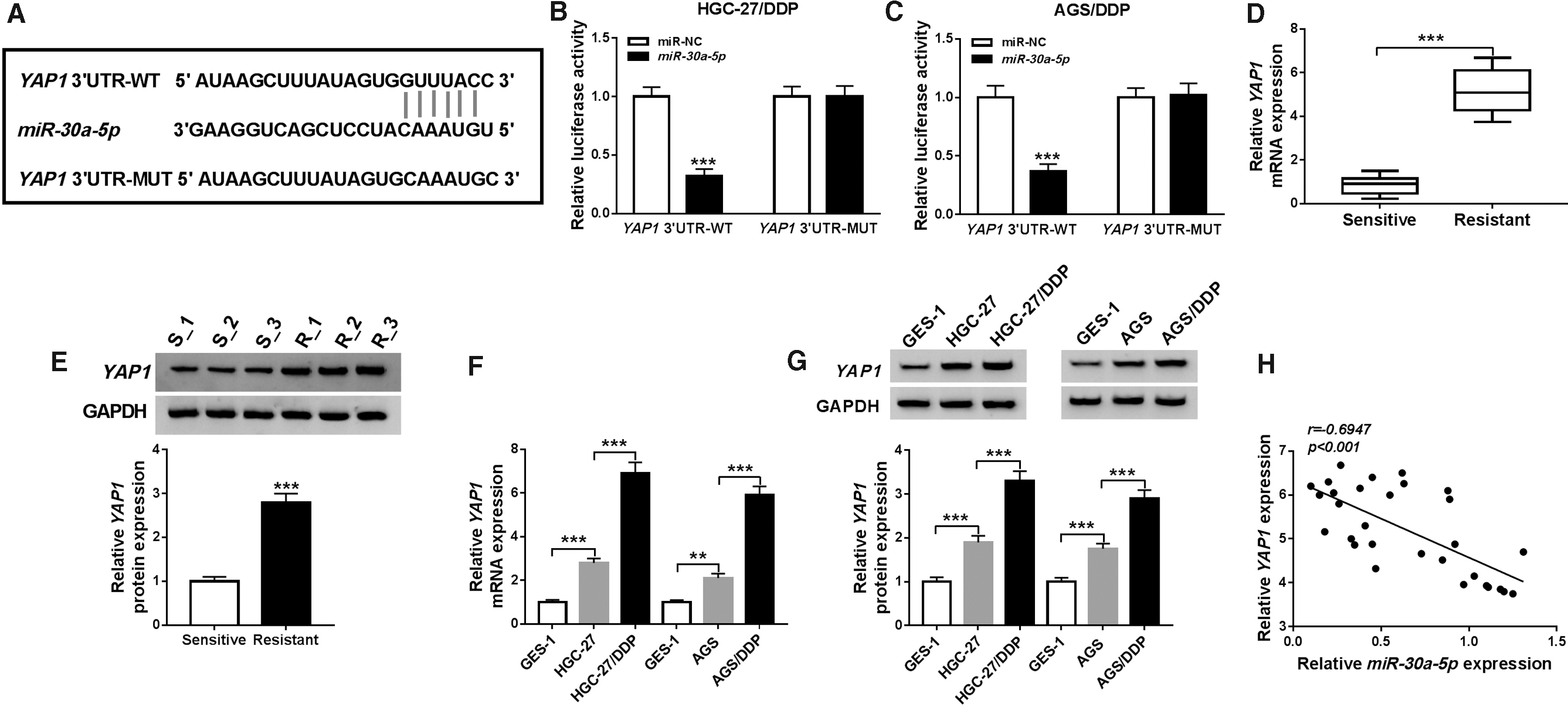
miR-30a-5p directly interacted with YAP1.
Overexpression of miR-30a-5p reduced DDP resistance in DDP-resistant GC cells via suppressing YAP1
Afterward, the modulatory regulation was ascertained after transfection with miR-30a-5p, miR-30a-5p+YAP1, or corresponding controls. Western blot manifested that miR-30a-5p transfection resulted in a lessening of YAP1 protein expression, which was abrogated by YAP1 upregulation in HGC-27/DDP and AGS/DDP cells (Fig. 5A, B). MTT assay demonstrated that the inhibition of IC50 of DDP induced by miR-30a-5p was reverted after YAP1 expression was enhanced (Fig. 5C). Also, overexpression of miR-30a-5p evoked the increase of apoptosis rate (Fig. 5D) and the descent of invaded cells (Fig. 5E), whereas ectopic expression of YAP1 abolished these effects. In addition, transfection of YAP1 reversed the repressive effects on LC3-II/I and P-gp protein levels and stimulative impact on P62 level motivated by miR-30a-5p (Fig. 5F, G). All in all, miR-30a-5p overexpression reduced DDP resistance in DDP-resistant GC cells by motivating apoptosis and repressing invasion or autophagy via targeting YAP1.

Overexpression of miR-30a-5p reduced DDP resistance in DDP-resistant GC cells via suppressing YAP1.
Knockdown of circ-PVT1 suppressed YAP1 expression by expediting miR-30a-5p level
To study whether YAP1 was affected by circ-PVT1, the authors designed the groups of si-circ-PVT1, si-circ-PVT1+anti-miR-30a-5p to transfect into HGC-27/DDP and AGS/DDP cells, including their respective controls. Western blot showed that si-circ-PVT1 transfection led to a downregulation of YAP1 protein expression, while the intervention of anti-miR-30a-5p ameliorated this inhibition in HGC-27/DDP (Fig. 6A) and AGS/DDP (Fig. 6B) cells. These results proved that circ-PVT1 knockdown could inhibit YAP1 expression via promoting miR-30a-5p.
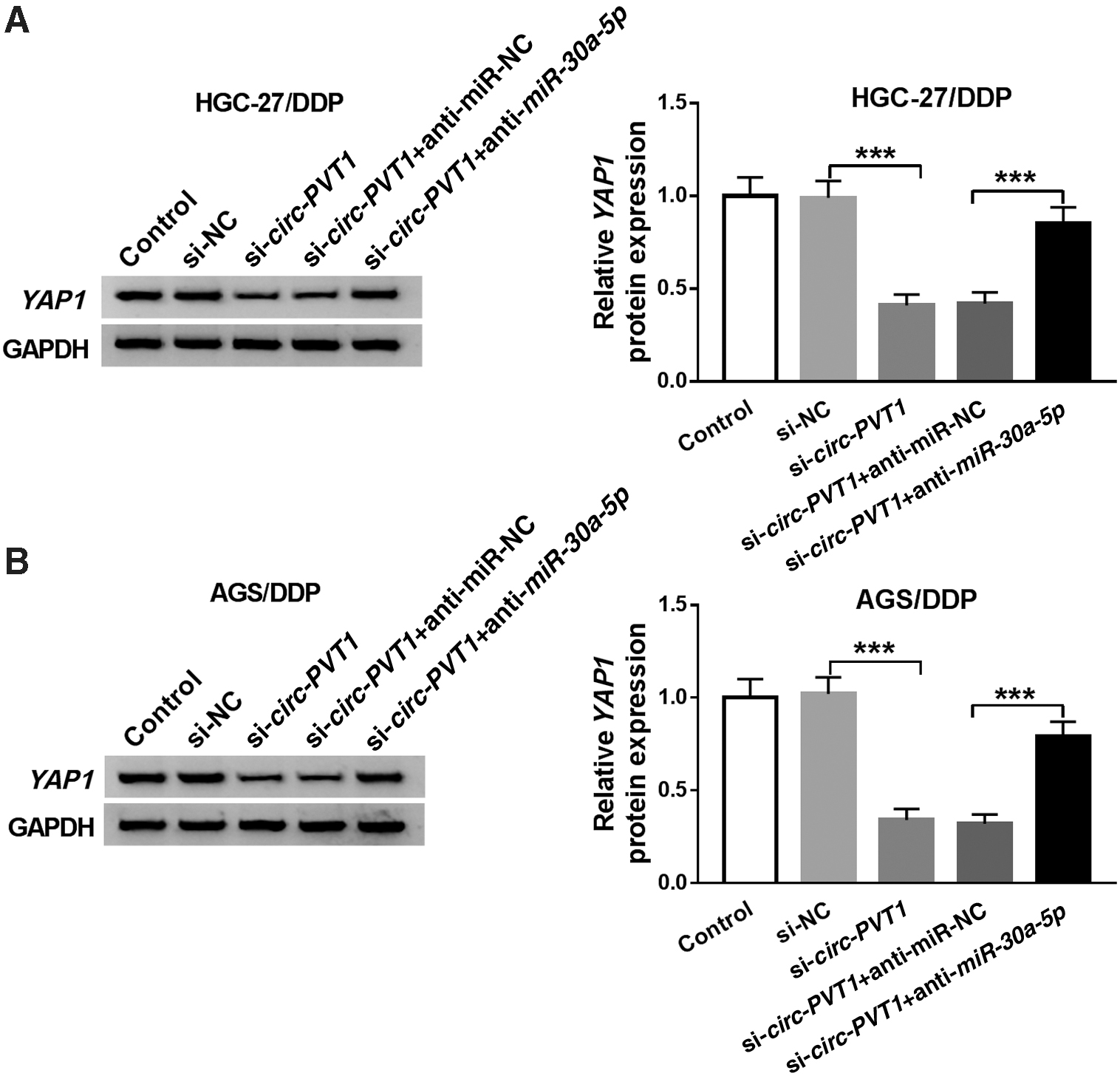
Knockdown of circ-PVT1 suppressed YAP1 expression by expediting miR-30a-5p level.
Circ-PVT1 depression improved DDP sensitivity of GC by miR-30a-5p/YAP1 axis in vivo
Finally, the xenograft tumor model was constructed to explore the influence of circ-PVT1 on DDP resistance in vivo. As described in Figure 7A, B, tumor volume and weight were lower in sh-circ-PVT1+DDP group than that in sh-NC+DDP group. Then, qRT-PCR revealed that knockdown of circ-PVT1 reduced the expression of circ-PVT1 (Fig. 7C), and there was an inverse phenomenon of miR-30a-5p level (Fig. 7D). Besides, the downregulation of YAP1 protein expression was shown in sh-circ-PVT1+DDP group compared with sh-NC+DDP group (Fig. 7E, F). Therefore, circ-PVT1 knockdown inhibited GC tumor growth by enhancing DDP sensitivity via miR-30a-5p/YAP1 axis in vivo.

Circ-PVT1 depression improved DDP sensitivity of GC by miR-30a-5p/YAP1 axis in vivo.
Discussion
It is imperative to explore crucial molecules and regulatory mechanism involved in the chemoresistance of GC. In this report, exosomal circ-PVT1 was found to be upregulated in GC with DDP resistance, and circ-PVT1 promoted DDP resistance by regulating autophagy, invasion, and apoptosis via miR-30a-5p/YAP1 axis in GC cells, which might be favorable for the DDP chemotherapy for GC patients.
Exosomes carried RNAs can involve the development of GC. 14,30 Zhang et al. found that exosomal circ-NRIP1 promoted GC evolution by sponging miR-149-5p. 31 Consistent with the finding, the authors found that circ-PVT1 was enriched in exosomes from GC serum samples and cells. Also, circ-PVT1 expressed highly in exosomes from DDP-resistant GC serums and cells, suggesting that exosomal circ-PVT1 might generate critical function in regulating DDP resistance of GC. Circ-PVT1 has been exhibited to be associated with drug resistance in several cancers. Zhu et al. attested that overexpression of circ-PVT1 facilitated doxorubicin and DDP resistance of osteosarcoma cells via the regulation of ABCB1. 32 In conformity with this discovery, circ-PVT1 contributed to the DDP resistance of GC cells in their article. Herein, the authors investigated the autophagy-related proteins. The upregulation of the LC3-II/I ratio represents the promotion of autophagy and the P62 increase symbolizes the inhibition of autophagy. 33 They testified that the role of circ-PVT1 on DDP resistance was achieved by inducing autophagy and invasion, but inhibiting cell apoptosis in GC cells.
CircRNAs function as microRNAs (miRNAs) “sponges” to play their parts generally. 34 MiR-30a-5p was identified as a target of circ-PVT1 in the study. MiR-30a-5p had a low expression in GC cells and inhibited cell proliferation and invasion. 35 The authors not only found the downregulation of miR-30a-5p in GC cells but also in DDP-resistant serums and cells, implying the involvement of miR-30a-5p in DDP resistance of GC. Yang et al. asserted that miR-30a-5p exerted the repression of DDP resistance in human small cell lung cancer. 36 In this article, miR-30a-5p inhibited DDP resistance of GC cells too, and circ-PVT1 regulated DDP resistance by targeting miR-30a-5p.
Subsequently, YAP1 was predicted as a target candidate of miR-30a-5p by StarBase v2.0, and dual-luciferase reporter assay validated the combination between miR-30a-5p and YAP1. YAP1, a member of YAP family, was reported to participate in the progression of many human cancers. 37 Tang et al. claimed that YAP1 had stimulative effects on tumorigenesis and chemoresistance in GC. 38 In this study, the authors discovered the high ectopic expression of YAP1 in DDP-resistant GC serum samples and cells, hinting the potential function of YAP1 in DDP resistance of GC. As expected, they verified that the accumulation of YAP1 counteracted the inhibition of DDP resistance by the miR-30a-5p intervention. In addition, circ-PVT1 regulated YAP1 expression by sponging miR-30a-5p in DDP-resistant GC cells. Furthermore, the depletion of circ-PVT1 could improve DDP sensitivity of GC via miR-30a-5p/YAP1 axis in vivo, and YAP1 can act as a key regulator to control the expression of downstream target genes (ANKRD1, Cyr61, and CTGF) in the Hippo pathway, further regulating cell proliferation and growth in cancers. 38 Whether circ-PVT1 can affect Hippo signaling pathway via mediating YAP1 in GC cells remains to be explored in their further research.
Conclusion
In summary, exosomal circ-PVT1 aggravated the generation of DDP resistance in GC cells through regulating autophagy, invasion, and apoptosis via negatively targeting miR-30a-5p and indirect regulation of YAP1. The exosomal circ-PVT1/miR-30a-5p/YAP1 regulatory axis might be useful for the DDP therapy in GC, affording a novel option in the molecular regulation for GC chemotherapy. The exosomal circ-PVT1 had the potential to be a promising diagnostic biomarker for GC treatment.
Footnotes
Authors' Contributions
Conception and design: W.Y.; Development of methodology: Q.M.; Acquisition of data: P.G.; Analysis and interpretation of data: Y.W.; Writing, review, and revision of article: W.Y.; All coauthors have reviewed and approved of the article before submission.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.