
Abstract
Objective:
To understand the regulation of long noncoding RNA DLX6-AS1-mediated miR-26a/EZH2 axis in the growth of colorectal cancer (CRC) cells.
Methods:
The expression of DLX6-AS1, miR-26a, and EZH2 was detected in CRC tissues by quantitative reverse transcription–polymerase chain reaction. The CRC HT-29 cell line was selected for transfection and subjected to observe the growth by MTT and colony formation assays, cell cycle by flow cytometry, and migration and invasion by wound healing and Transwell assays, respectively. Finally, the expression of cycle- and metastasis-related proteins was detected by Western blotting.
Results:
DLX6-AS1 and EZH2 were increased, with a decreased miR-26a in CRC tissues, showing significant negative correlations between DLX6-AS1 and miR-26a, and between miR-26a and EZH2. CRC patients at advanced stage or with lymphatic metastasis had higher DLX6-AS1 expression. Dual-luciferase reporter gene assay uncovered the targeting correlations between DLX6-AS1 and miR-26a, or miR-26a and EZH2. After transfection of DLX6-AS1 siRNA or EZH2 siRNA, the growth and metastasis of CRC cells were suppressed, arresting the cells in G0/G1 phase, with a magnificent reduction in the ratio of cells in S phase or G2/M phase; meanwhile, Cyclin D1, Vimentin, and MMP9 expressions decreased evidently, whereas E-cadherin expression was upregulated. Changes above were fully reversed after transfection of miR-26a inhibitor, whereas si-EZH2 transfection abolished the positive role of miR-26a inhibitor on growth of CRC cells.
Conclusion:
Silencing DLX6-AS1 may block the malignant features of CRC cells by inhibiting the expression of EZH2 through upregulation of miR-26a. Thus, it is critical to the development and progression of CRC.
Introduction
Colorectal cancer (CRC), according to the global statistical evidence, is the third most prevalent cancer and fourth in cancer-related deaths globally, with 16 million new cases annually and 771,000 deaths. 1 Despite great progress in the early diagnosis of CRC, many patients have progressed to an advanced stage and when diagnosed with little opportunity for surgical treatment, it made them choose chemotherapy or radiotherapy, with still unsatisfactory 5-year survival rate. 2 –4 CRC development and progression has been known to be a multistage and multistep process under various environmental and genetic factors, with little information regarding its pathogenesis. 5 Thus, exploring the pathogenesis of CRC and searching for the therapeutic target has been the major issues to be urgently solved in current studies of CRC.
Long noncoding RNA (lncRNA) and microRNA are critical noncoding RNAs in human genome. 6,7 As reported, lncRNA, as a kind of competitive endogenous (ceRNA), is involved in regulating the expression of target gene, thereby participating in various biological events. 8,9 lncRNA DLX6-AS1, located at chromosome 7q21.3, is a developmentally regulated lncRNA. 10 Latest evidence suggested the pivotal role of DLX6-AS1 as a ceRNA in tumor progression. For instance, Zhang et al. reported that DLX6-AS1 could promote the growth of HCC cells through modulating the miR-203a/MMP-2 pathway. 10 Zeng et al. reported that DLX6-AS1 was overexpressed in renal cell carcinoma, which could facilitate cell growth and tumorigenesis through miR-26a/PTEN axis. 11 As for miR-26a, it has been identified to be extensively expressed, and the dysregulation of miR-26a was implicated in the pathogenesis of solid tumors and malignancies in hemopoietic system. 12,13 Li et al. found that miR-26a, as the tumor suppressor gene, can inhibit the malignant biological features of CRC cells, 14 which could also repress the in vivo and in vitro metastasis of nasopharyngeal cancer cells by targeting EZH2, as indicated by Yu et al. 15 However, existing evidence has confirmed that EZH2, a member of polycomb protein complex family, was highly expressed in CRC, which may serve as a potential target in the treatment of tumors, including CRC. 16,17 Through the online bioinformatics tool, the potential binding sites between DLX6-AS1 and miR-26a, as well as between miR-26a and EZH2 was also pointed out. Considering the above, we inferred that DLX6-AS1 may affect the progression of CRC by regulating miR-26a/EZH2 axis.
In this study, we aimed to understand how DLX6-AS1 regulates the growth of CRC cells by mediating miR-26a/EZH2 axis, hoping to provide a new theoretical basis for the prophylaxis and treatment of CRC.
Methods
Ethical statement
Informed consent was obtained from every patient before the study, and this study was approved by the Ethics Committee on Clinical Trials of our hospital.
Subjects
Tissue specimens (including CRC tissues and the corresponding adjacent normal tissues) were obtained from 76 CRC patients who underwent radical resection of CRC and diagnosed as CRC as per the postoperative pathological test in this hospital between June 2015 and June 2018. Patients were between 36 and 82 years, with an average at 59.11 ± 6.60 years. Based on the standard issued by The American Joint Committee on Cancer, 18 there were 35 patients in Stage I or II and 41 patients in Stage III or IV. Lymphatic metastasis was identified in 33 patients, but not in other 43 patients. All patients reported no history of chemotherapy or radiotherapy before the surgery, and the specimens were preserved at −80°C.
Cell culture
Normal human colorectal epithelial cell line (NCM460) and different CRC cell lines (DLD-1, HCT-116, HT-29, SW480, and SW620) were purchased from the American Type Culture Collection. All cell lines were cultured in a thermostat incubator (Thermo) with 5% CO2 at 37°C in the RPMI-1640 (Hyclone) supplemented with 10% deactivated fetal bovine serum (FBS; Gibco), 100 U/mL penicillin (Hyclone), and 100 mg/mL streptomycin (Hyclone).
Cell grouping and transfection
HT-29 cells were selected for transfection and grouped as follows: control group (no transfection), si-NC group (transfected by NC siRNA), si-DLX6-AS1 group (transfected by DLX6-AS1 siRNA), miR-26a inhibitor group (transfected by miR-26a inhibitor), si-DLX6-AS1 + NC inhibitor group (cotransfected by DLX6-AS1 siRNA and NC inhibitor), si-DLX6-AS1 + miR-26a inhibitor group (cotransfected by DLX6-AS1 siRNA and miR-26a inhibitor), si-EZH2 group (transfected by EZH2 siRNA), and miR-26a inhibitor + si-EZH2 group (cotransfected by miR-26a and EZH2 siRNA). In this experiment, DLX6-AS1 siRNA, NC siRNA, EZH2 siRNA, miR-26a inhibitor, and miR-negative control were all provided by Guangzhou RiboBio Co., Ltd. Before transfection, cells were seeded on the 24-well plates, and transfected with Lipofectamine 2000 as per the instruction at confluence of ∼70%–80% (Invitrogen).
Quantitative reverse transcription–polymerase chain reaction
RNA extracts from the tissues or cells by use of the TRIzol reagent were used for preparing the cDNA with the TaqMan RNA Reverse Transcription kit (Applied Biosystems). Thereafter, PCR was performed by use of TaqMan Universal PCR kit (Qiagen, Germany). Primers used in this experiment were synthesized by Beijing Genomics Institute (Table 1). The relative expressions of the targeted genes were quantified with GAPDH and U6 as internal reference of DLX6-AS1 and EZH2, or miR-26a, respectively.
Primer Sequences of Quantitative Reverse Transcription–Polymerase Chain Reaction in This Study
Dual-luciferase reporter gene assay
DLX6-AS1/EZH2 3′-UTR fragment containing the binding site of miR-26a was inserted into the vector carrying the PGL3 luciferase reporter gene to construct the DLX6-AS1–3′UTR-WT plasmid, DLX6-AS1–3′UTR-MUT plasmid, EZH2–3′UTR-WT plasmid, and EZH2–3′UTR-MUT plasmid. Then, HT-29 cells were transfected by the WT/MUT plasmids carrying NC mimic or miR-26a mimic according to the instruction of LipofectamineTM 2000. After 24 h, the original medium was discarded, and cells were rinsed twice using phosphate-buffered saline (PBS). Thereafter, the activity of the luciferase in cells to be detected was expressed by the ratio of the activity of firefly luciferase to that of the renilla luciferase.
MTT assay
Cells were seeded on the 96-well plates at 1 × 104 cell/well in 200 μL medium, followed by culture at 37°C with 5% CO2. At 24, 48, and 72 h, respectively, 20 μL of MTT solution was added into each well, where cells were further incubated for another 4 h at 37°C. Subsequently, 150 μL of dimethylsulfoxide was added by shaking for 10 min without the supernatant. The optical density value of each well was determined at a wavelength of 490 nm using a microplate reader (MultiSkan FC; ThermoFisher, NY). This experiment was conducted in triplicate, and results were averaged as the final result.
Colony formation assay
The cultured cells in good growth were selected and digested in trypsin after three washes in PBS. After repeated blowing and beats, cells were distributed evenly in the suspension, 200 of which were then seeded on the six-well plates in each well. Plates were shaken slightly to disperse the cells evenly, followed by 2 weeks of culture at 37°C and 5% CO2. Once the cell colony became visible, culture was terminated by discarding the medium and two washes in PBS. Then, cells were dried in air, fixed in methanol, and stained in crystal violet for 15 min, followed by cell counting.
Wound healing assay
After being seeded on the six-well plates, cells were cultured to saturation. Without the supernatant, cells were rinsed in PBS twice, and a line was drawn slightly by use of a sterilized 10 μL Eppendorf tip. Thereafter, cells were rinsed gently twice, and cultured in 2 mL of RPMI-1640 medium supplemented with 10% FBS. At 0 and 48 h, cells in each group were observed and photographed under the inverted microscope to measure the wound length.
Transwell invasion assay
In the serum-free medium, cells were starved for 12 h, and then digested by trypsin for preparation of cell suspension in FBS-free Dulbecco's modified Eagle's medium, where the cell density was adjusted to 2 × 106/mL. After ultraviolet radiation for 2 h, Transwell chambers were placed in 24-well plate, and in each well, 100 μL of Matrigel was added and coagulated at 37°C and 5% CO2 for 4 h. Afterward, 100 μL of cell suspension was added into the upper chambers, and 800 μL of RPMI containing 10% FBS into the lower chambers. At 48 h, cells in the lower chambers were fixed in 90% ethanol, stained in 0.1% crystal violet, and counted under the microscope.
Western blotting
In RIPA solution (R0010; Beijing SolarBio Co., Ltd., Beijing, China), total proteins were extracted from the freshly collected tissue specimens, and the concentration was determined by use of PierceTM BCA Protein Assay Kit (Pierce Biotechnology, Rockford, IL). A total of 50 μg proteins were loaded for sodium dodecyl sulfate–polyacrylamide gel electrophoresis, followed by transferring the proteins into a nitrocellulose membrane. On the membrane, the unoccupied sites were blocked by incubating with the skimmed milk. After that, proteins on the membrane were probed by incubating with the primary mouse anti-human monoclonal antibodies against EZH2 (Abcam; ab32199), Cyclin D1 (Abcam; ab32199), Vimentin (Abcam; ab32199), MMP9 (Abcam; ab32199), E-cadherin (Abcam; ab32199), and β-actin (Abcam; ab8226) at 4°C overnight. After three washes in PBS (10 min/wash), the resultant immunoblots were further probed by incubating the horseradish peroxidase-conjugated secondary antibodies (Abcam; ab6785). The immunoblots were detected by incubation with enhanced chemiluminescence reagent, and the bands were developed in a gel imaging system.
Statistical analysis
All data from the three replicates of each experiment were processed using SPSS 21.0 software (SPSS, Inc., Chicago, IL). Measurement data were given as mean ± standard deviation, and compared using t-test between two groups or one-way analysis of variance followed by Tukey's post hoc for multiple group comparisons. p < 0.05 was considered statistically significant.
Results
Expression of DLX6-AS1 in CRC tissues and cell lines
Results of quantitative reverse transcription–polymerase chain reaction (qRT-PCR) indicated that a significant increase of DLX6-AS1 expression was identified in CRC tissues compared with tumor-adjacent tissues (all p < 0.05; Fig. 1A). Meanwhile, the CRC cell lines (including DLD-1, HCT-116, HT-29, SW480, and SW620) also manifested sharp increases in the expression of DLX6-AS1 cells relevant to the normal human colorectal epithelial cell line (NCM460), among which HT29 cells showed the most evident increases; hence, HT29 cells were eligible for the following in vitro experiments (all p < 0.05; Fig. 1B). In addition, DLX6-AS1 expression was correlated significantly with the lymphatic metastasis and TNM stages of CRC patients, and those patients with higher stage or lymphatic metastasis presented higher expression of DLX6-AS1 (all p < 0.05; Fig. 1C, D); however, it was not related to age, gender, tumor size, or differentiation (all p > 0.05).
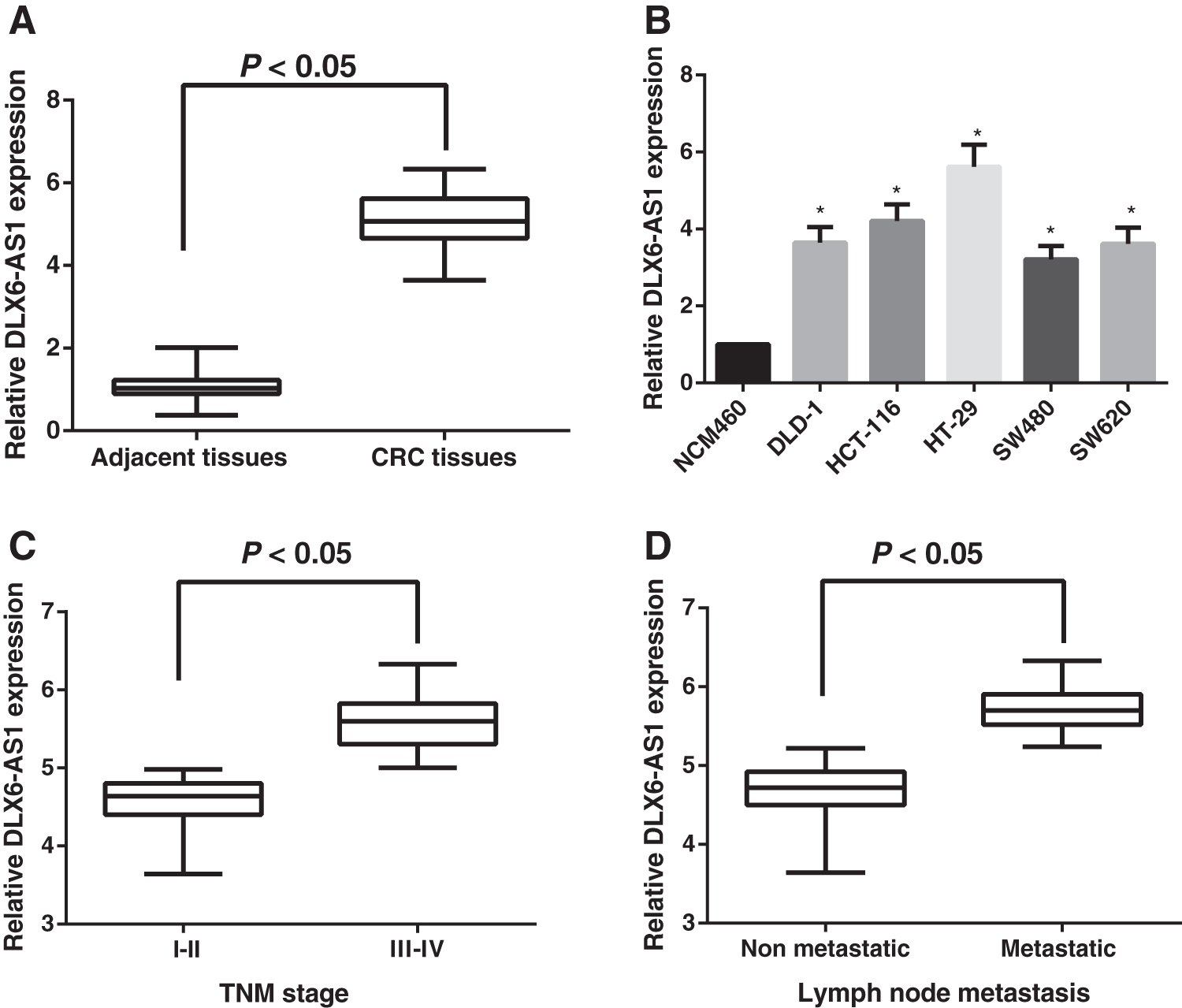
Expression of DLX6-AS1 in CRC tissues and cell lines.
Validation of the targeting relationship of miR-26a with DLX6-AS1 and EZH2
The expression of miR-26a and EZH2 in the CRC tissues was analyzed using qRT-PCR, and as indicated by the results, the miR-26a level decreased evidently but EZH2 expression showed the opposite changes in the CRC tissues compared with the tumor-adjacent tissues (all p < 0.05; Fig. 2A, B). The correlation analysis revealed negative correlations between DLX6-AS1 and miR-26a expressions (r = −0.813, p < 0.05; Fig. 2C), and between miR-26a and EZH2 expressions (r = −0.738, p < 0.05, Fig. 2D). According to the predictions of Starbase (
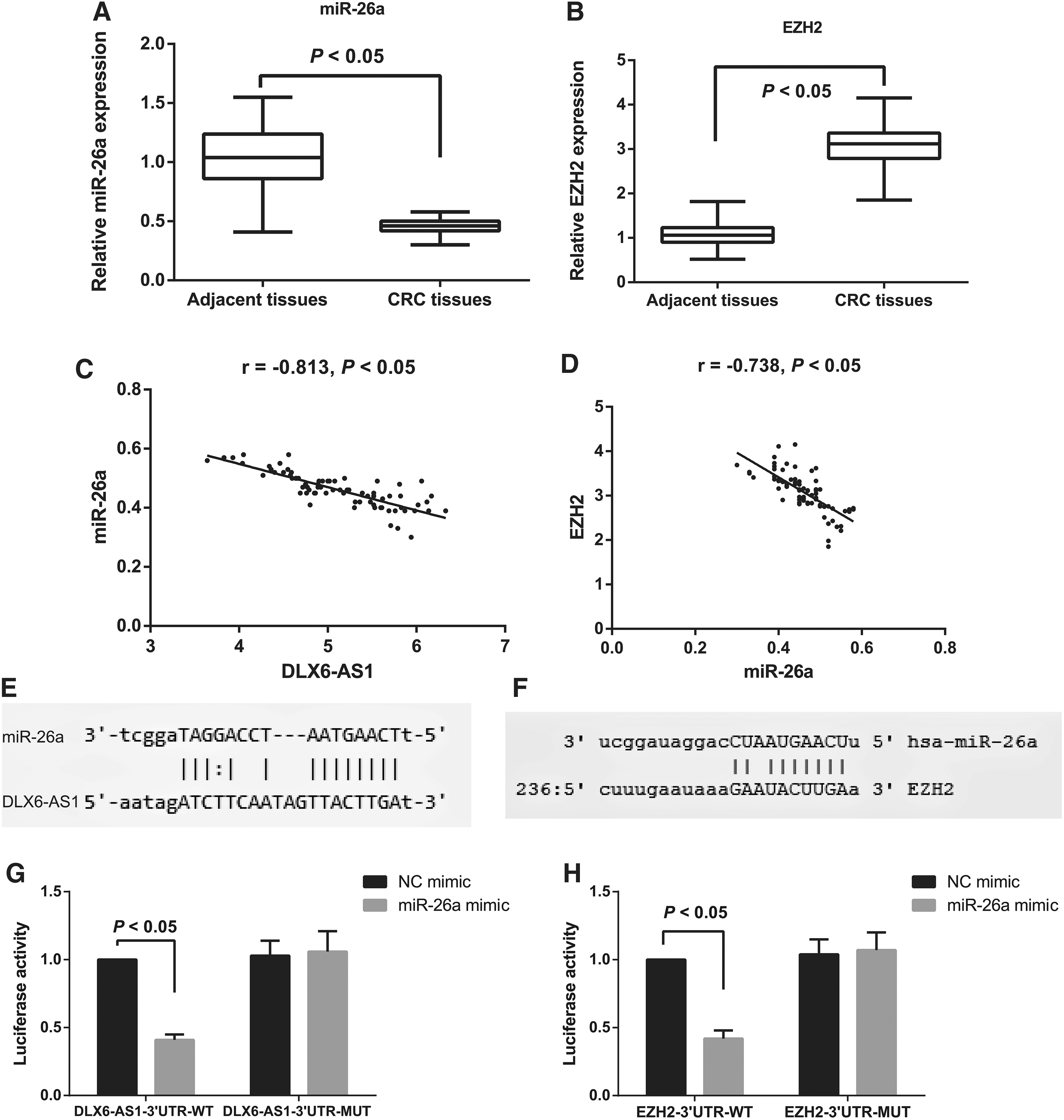
Validation of the targeting relationship of miR-26a with DLX6-AS1 and EZH2.
Expressions of DLX6-AS1, miR-26a, and EZH2 in transfected cells
The technique of qRT-PCR and Western blotting was used to measure the expressions of DLX6-AS1, miR-26a, and EZH2 in transfected cells (Fig. 3). Compared with the control group, no significant difference was found from cells in the si-NC group with regard to the expressions of DLX6-AS1, miR-26a, and EZH2 (all p > 0.05), but cells in the si-DLX6-AS1 group and si-DLX6-AS1 + NC inhibitor group showed downregulation of DLX6-AS1 and EZH2, and upregulation of miR-26a (all p < 0.05). Meanwhile, transfection of miR-26a inhibitor led to upregulated DLX6-AS1 and EZH2 and downregulated miR-26a (all p < 0.05). There were no significant changes in expressions of DLX6-AS1 and miR-26a after transfection of si-EZH2, although EZH2 was downregulated evidently (all p < 0.05). Moreover, higher expression of DLX6-AS1 and EZH2 and lower expression of miR-26a were found in si-DLX6-AS1 + miR-26a inhibitor group than si-DLX6-AS1 group (all p < 0.05). In addition, cotransfection of miR-26a inhibitor and si-EZH2 could further decrease the level of EZH2 compared with those transfected by miR-26a inhibitor alone (all p < 0.05).
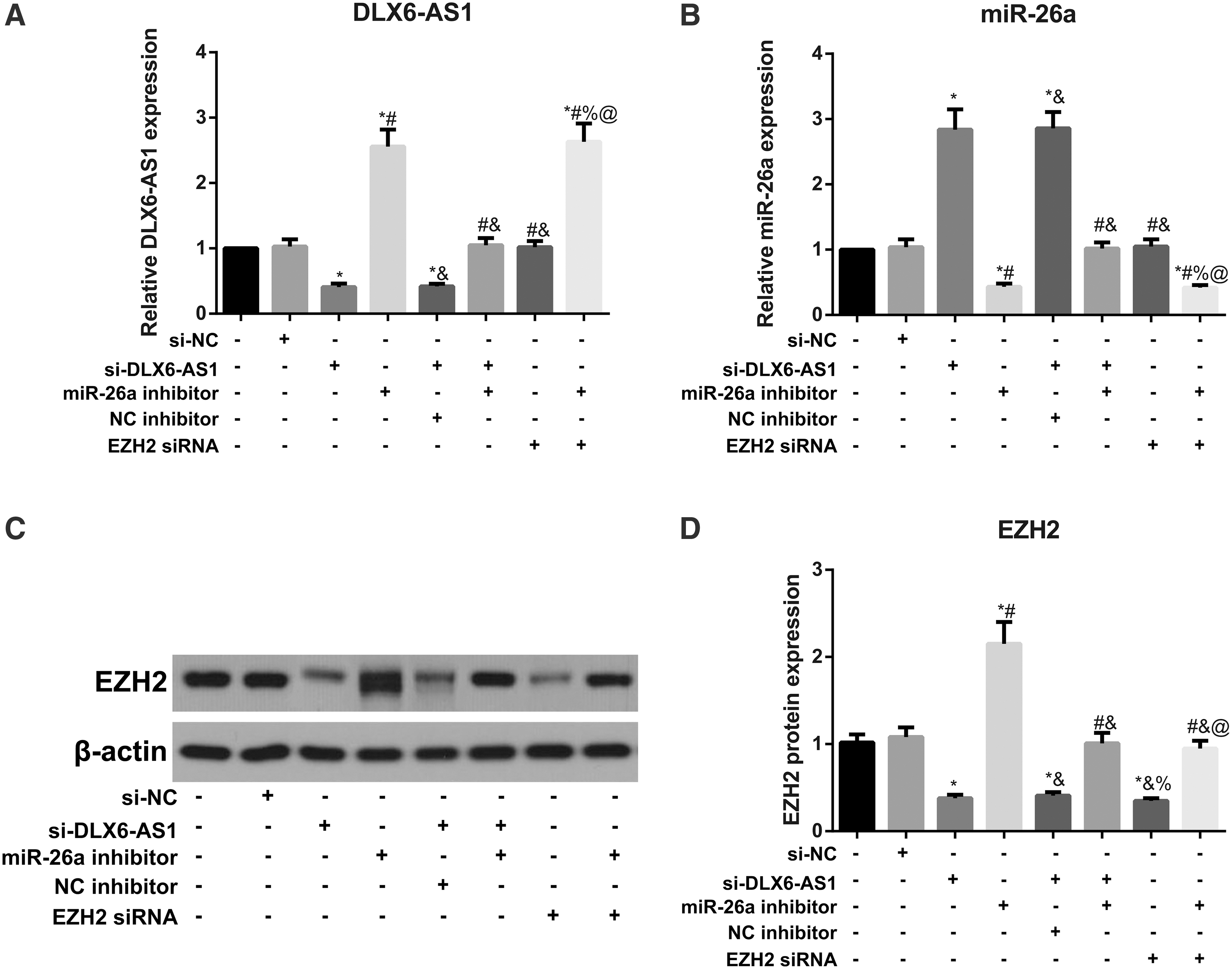
Expressions of DLX6-AS1, miR-26a, and EZH2 in transfected cells.
DLX6-AS1-mediated miR-26a/EZH2 axis to involve in the proliferation of CRC cells
The proliferation of CRC cells was evaluated by MTT assay and colony formation assay (Fig. 4). As a result, the proliferation ability and colony number in cells from the control group were not different from those transfected by si-NC (all p > 0.05), but were decreased significantly in cells transfected by si-DLX6-AS1, or si-EZH2, or cotransfected by si-DLX6-AS1 and NC inhibitor (all p < 0.05), which, however, were totally reversed by the transfection of miR-26a inhibitor. The proliferation ability of cells was higher in si-DLX6-AS1 + miR-26a inhibitor group than in the si-DLX6-AS1 group, but was lower in miR-26a inhibitor + si-EZH2 than in the miR-26a inhibitor group (all p < 0.05).
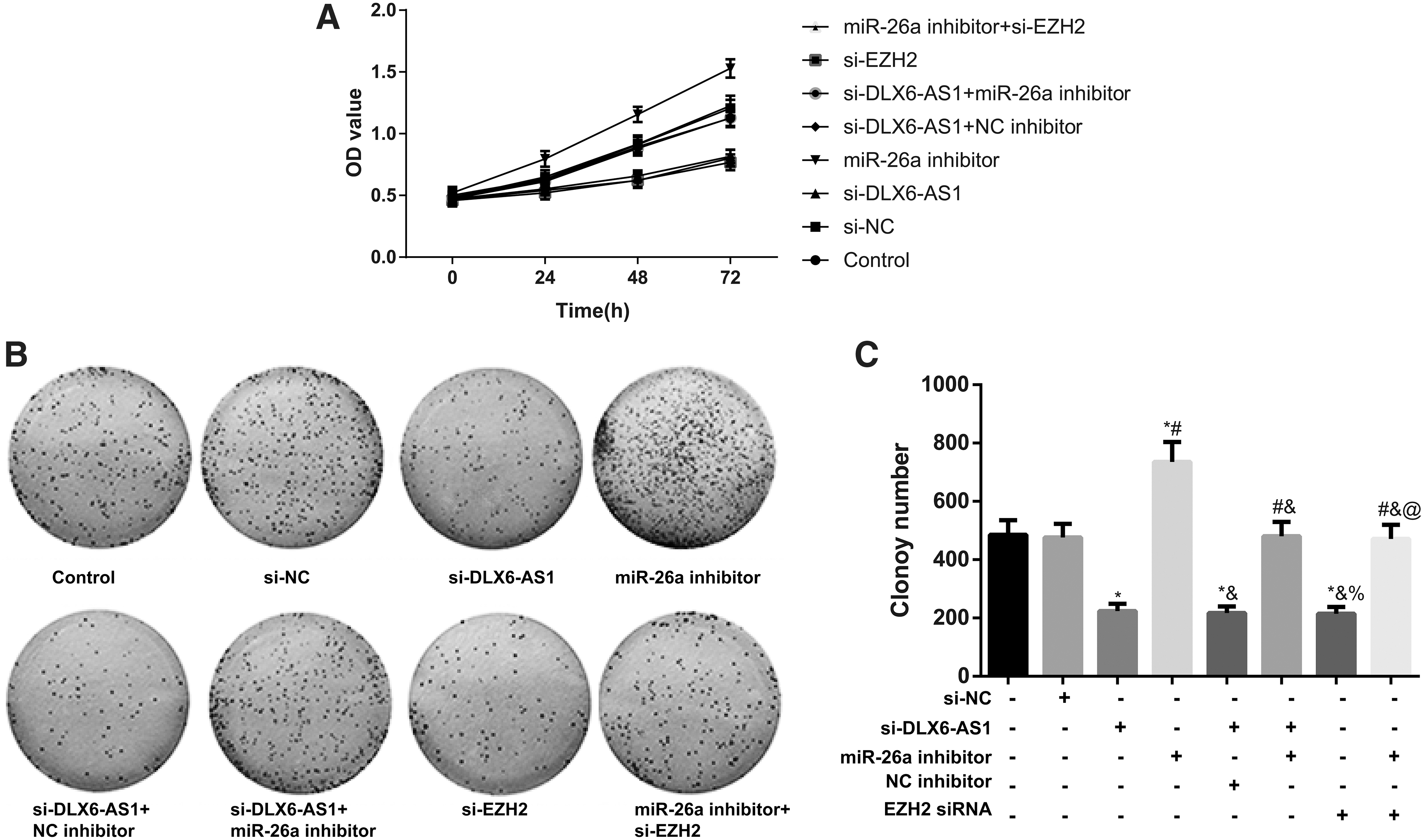
DLX6-AS1-mediated miR-26a/EZH2 axis to involve in the proliferation of CRC cells.
DLX6-AS1-mediated miR-26a/EZH2 axis to regulate the cycle of CRC cells
As given in Figure 5, cells in the si-DLX6-AS1 group and si-DLX6-AS1 + NC inhibitor group had a significant increase in the proportion of cells in G0/G1 phase, but a decrease in the proportion of cells in S or G2/M phase in comparison with the control group (all p < 0.05), whereas transfection of miR-26a inhibitor led to opposite changes. Compared with the si-DLX6-AS1 group, cotransfection of si-DLX6-AS1 and miR-26a inhibitor decreased the proportion of cells in G0/G1, but augmented that of cells in S or G2/M phase (all p < 0.05). However, compared with the miR-26a inhibitor group, co-transfection of miR-26a inhibitor and si-EZH2 further increased the proportion of cells in G0/G1 phase, but decreased that of cells in S or G2/M phase (all p < 0.05).

Distribution of cell cycles determined by flow cytometry. *p < 0.05 versus control group and si-NC group; # p < 0.05 versus si-DLX6-AS1 group and si-DLX6-AS1 + NC inhibitor group; & p < 0.05 versus miR-26a inhibitor group; % p < 0.05 versus si-DLX6-AS1 + miR-26a inhibitor group; @ p < 0.05 versus si-EZH2 group.
DLX6-AS1-mediated miR-26a/EZH2 axis to regulate the migration and invasion of CRC cells
As given in Figure 6, no significant difference was noted in the invasion and migration of cells between the control group and si-NC group (all p > 0.05), but significantly declined after transfection of si-DLX6-AS1 or si-EZH2, or co-transfection of si-DLX6-AS1 and NC inhibitor (all p < 0.05). However, these changes were reversed with the transfection of miR-26a inhibitor (all p < 0.05). Cells with the co-transfection of si-DLX6-AS1 and miR-26a inhibitor had the enhanced migration and invasion compared with the si-DLX6-AS1 group, whereas those cotransfected with the miR-26a inhibitor and si-EZH2 had reduced cell migration and invasion relevant to the miR-26a inhibitor group (all p < 0.05).
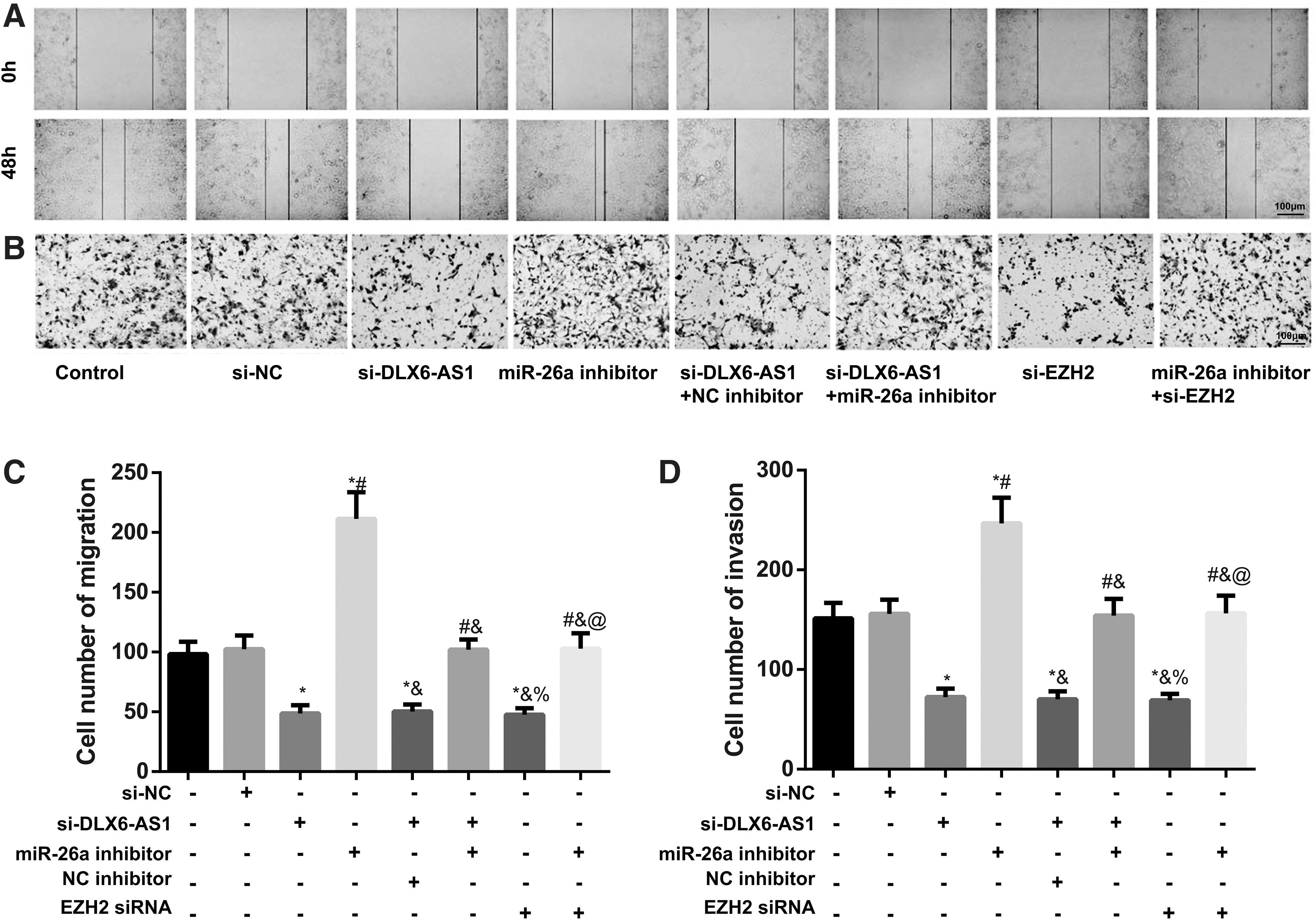
DLX6-AS1-mediated miR-26a/EZH2 axis to regulate the migration and invasion of CRC cells.
Comparison of the cycle- and metastasis-related proteins in CRC cells
The protein expressions of Cycline D1, Vimentin, and MMP9 were reduced evidently, whereas E-cadherin was upregulated evidently in the si-DLX6-AS1 group, si-DLX6-AS1 + NC inhibitor group, and si-EZH2 group compared with the control group, whereas transfection of miR-26a inhibitor resulted in opposite alterations (all p < 0.05; Fig. 7). Compared with the si-DLX6-AS1 group, significant increases were shown in Cyclin D1, Vimentin, and MMP9 in the si-DLX6-AS1 + miR-26a inhibitor group, with a significant decrease in E-cadherin (all p < 0.05). In comparison with the miR-26a inhibitor group, cotransfection of miR-26a and si-EZH2 decreased the expressions of Cyclin D1, Vimentin, and MMP9, but increased the expression of E-cadherin (all p < 0.05).
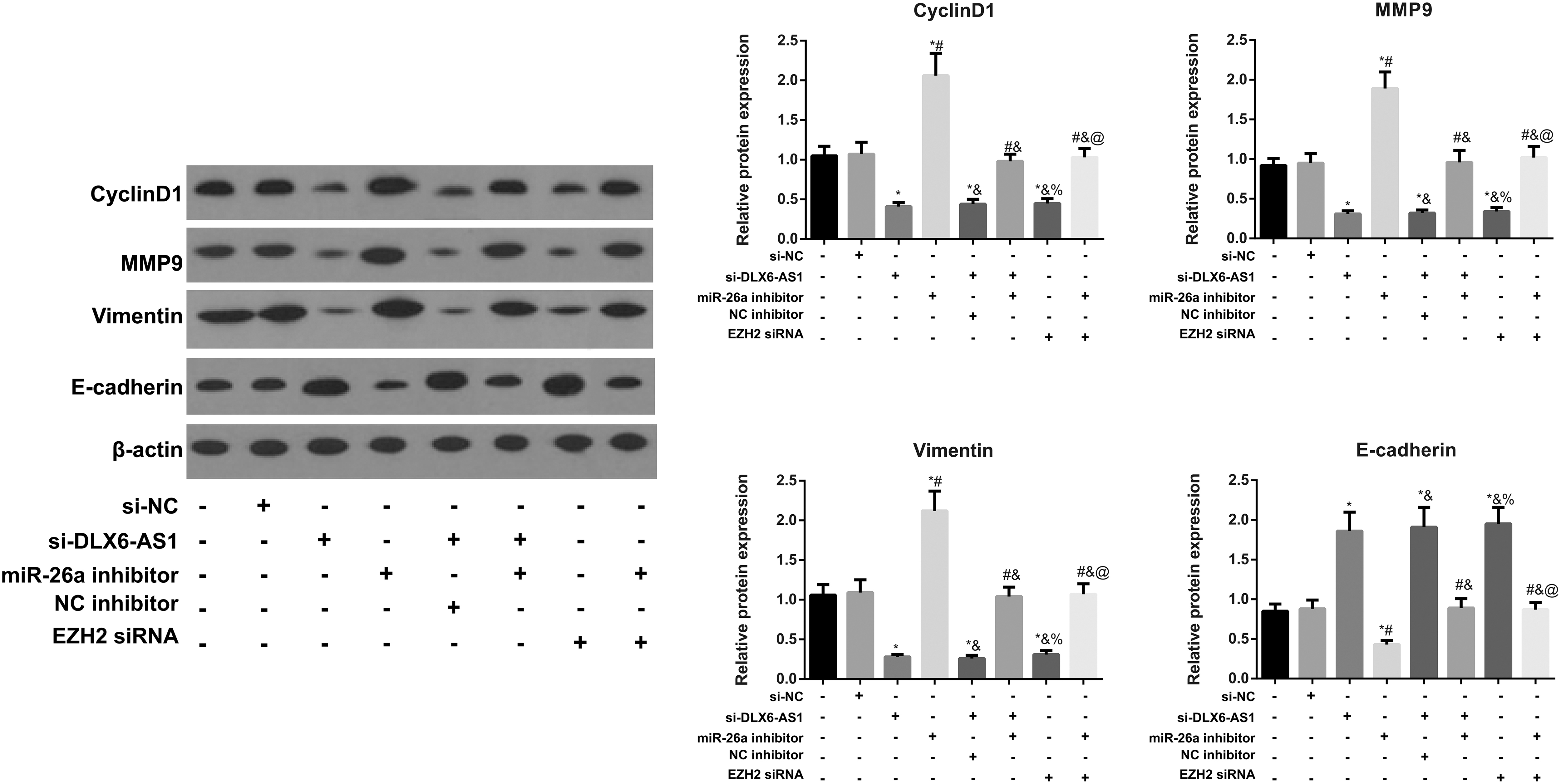
Expressions of cycle-related protein (Cyclin D1) and metastasis-related proteins (Vimentin, MMP9, and E-cadherin) in CRC cells detected by Western blotting. *p < 0.05 versus control group and si-NC group; # p < 0.05 versus si-DLX6-AS1 group and si-DLX6-AS1 + NC inhibitor group; & p < 0.05 versus miR-26a inhibitor group; % p < 0.05 versus si-DLX6-AS1 + miR-26a inhibitor group; @ p < 0.05 versus si-EZH2 group.
Discussion
We first found upregulation of DLX6-AS1 in CRC tissues and cells, which was further increased in those patients with higher TNM stage or lymphatic metastasis. Similarly, increased DLX6-AS1 was also identified in lung adenocarcinoma to be correlated with TNM stage and histological differentiation. 19 In gastric cancer, DLX6-AS1 upregulation may play an oncogenic role to be associated with clinical stage, lymphatic metastasis, and distant metastasis. 20 Besides, our study found downregulation of miR-26a in CRC tissues, with upregulation of EZH2, as detected by qRT-PCR, which was in line with the findings observed by Ghanbari et al. 21 and Fluge et al. in CRC, and EZH2 overexpression in CRC was related to high proliferation and poor prognosis of patients. 16 In particular, we further revealed negative correlations between DLX6-AS1 and miR-26a, and between miR-26a and EZH2, and the targeting relationship among them was analyzed in dual-luciferase reporter gene assay. Thus, it is reasonable to suppose that DLX6-AS1 may exert its effects on the progression of CRC through modulation of miR-26a/EZH2 axis.
To validate this hypothesis, we conducted the in vitro transfection on CRC HT29 cells, and then either silencing DLX6-AS1 or EZH2 inhibition could surppress CRC cell proliferation, and arrested the cells at G0/G1 phase, with increases in the proportions of cells in S phase or G2/M phase. However, inhibition of miR-26a promoted cell cycle and proliferation. Likewise, Li et al. found that DLX6-AS1 knockout resulted in the decreased proliferation and growth of HCC cells by targeting miR-424-5p, leading to the cycle arrest of cells at G0/G1 phase. 22 In general, the uncontrolled proliferation of tumor cells was suggested to be mainly attributed by the loss modulation of cell cycle. 23 EZH2 may promote cell proliferation by accelerating the transition of cell cycle from S phase to G2/M phase. 24 Cyclin D1, a key role in G1 phase, can facilitate the transition from G1 to S phase in cell cycle by forming a complex with cyclin-dependent protein kinase (Cdk) 4 and 6, finally causing the uncontrolled malignant changes in tumor cells. 25,26 Of interest, our results showed that suppressing DLX6-AS1 or EZH2 did decrease the expression of Cyclin D1, which was abolished by inhibition of miR-26a. Similarly, Yang et al. noted that DLX6-AS1 upregulated Cyclin D1 in pancreatic cancer by regulating the expression of miR-497-5p, thus promoting malignant changes. 27 Furthermore, after silenced DLX6-AS1, we found upregulation of miR-26a, but downregulation of EZH2, reversed by miR-26a inhibition. The inhibitory effect of si-DLX6-AS1 on cell proliferation in CRC was abolished by miR-26a inhibitor, the effect of which, however, was further reversed by si-EZH2. Xu et al. reported that the sponge absorption of SNHG6, by upregulating miR-26a/b, suppressed EZH2 and downregulated Cyclin D1 and CDK4/6, thereby curbing the proliferation, migration, and invasion of CRC cells and blocking cell cycles. 28 Therefore, inhibition of DLX6-AS1 could block the cell cycle and proliferation by suppressing the expression of EZH2 and Cyclin D1, which may be achieved by upregulation of miR-26a.
Accumulating evidence suggested that epithelium–mesenchymal transition is the critical pathway for the local infiltration and distant metastasis of epithelium-originated tumors. 29 EZH2 can catalyze the trimethylation of H3K27 in the promoter of gene that encodes E-cadherin, the marker of EMT, to block its expression. 30 The study of Min et al. discovered that in prostatic cancer, EZH2 binds to the promoter of DAB2IP to suppress the transcription, upregulate the expression of Vimentin and fibronectin (markers of mesenchyme), and downregulate the expression of E-cadherin to advance the tumor metastasis. 31 As a key link in local and distant metastasis of tumor cells, extracellular matrix degradation breaks down the balance between MMP and TIMP, thereby promoting the metastasis of tumor cells. 32,33 By suppressing TIMP and upregulating MMP in malignant tumors, EZH2 results in the degradation of extracellular matrix and the invasion of cancer cells. 34,35 After measurement in our study, silencing DLX-AS1 or EZH2 could decrease the expressions of Vimentin and MMP9, and increase the expression of E-cadherin, thus blocking cell migration and invasion; nevertheless, the transfection of miR-26a inhibitor triggered totally different changes. Liang et al. reported that in case of DLX6-AS1 knockout, the malignant features of gastric cancer cells were significantly suppressed, which could be achieved by the regulation of miR-204-5p/OCT1 axis. 36 According to the findings of Zeng et al., DLX6-AS1 was able to promote the tumorigenesis in renal cancer by targeting miR-26a/PTEN. 11 Consistently, Zhang et al. reported that lncRNA SNHG6 would bind to miR-26a to further regulate EZH2, with an increase in E-cadherin, and decreases in Snail, Vimentin, and N-cadherin, thus facilitating EMT, invasion, and migration of CRC cells. 37 Therefore, inhibition of DLX6-AS1, through upregulation of miR-26a, could downregulate EZH2 to modulate the downstream metastasis-related proteins, thereby inhibiting the malignant features of tumor cells.
To conclude, silencing DLX6-AS1 suppresses EZH2 expression by upregulating miR-26a to block the malignant biological features of CRC cells, which provide a new insight for CRC treatment. A limitation of this study was the relatively small number of patients and controls, and further large-scale experimental investigations are needed to verify the results of this study.
Ethics Approval and Consent to Participate
Informed content was obtained from every patient before the study, and this study was approved by the Ethics Committee on Clinical Trials of our hospital.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Footnotes
Acknowledgments
The authors give sincere appreciation to the reviewers for their helpful comments on this article.
Author Contributions
W.-Q.K. and J.-J.L. designed the article. J.D. and Z.-X.Y. performed and analyzed the data, and supervised the article. P.G. and Y.-L.L. drafted the article.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
No funding was received for this article.