
Abstract
Background:
Long non-coding RNA actin filament-associated protein1-antisense RNA 1 (AFAP1-AS1) was confirmed to be associated with tumorigenesis. However, the role of AFAP1-AS1 in breast cancer was little known.
Materials and Methods:
Quantitative real-time polymerase chain reaction (qRT-PCR) was used to detect the levels of AFAP1-AS1, microRNA-497-5p (miR-497-5p), and Septin 2 (SEPT2) in breast cancer tissues and cells. The cell proliferation, migration, and apoptosis were tested by Methylthiazolyldiphenyl-tetrazolium bromide (MTT), Transwell and Flow cytometry assays, respectively. The targeting relationship between genes was predicted by StarBase v.3.0 and confirmed by dual-luciferase reporter assay. Pearson's correlation coefficient was applied to examine the correlation between the two groups. SEPT2 protein expression was evaluated by Western blot. Xenograft models were established to investigate the role of AFAP1-AS1 knockdown in vivo.
Results:
AFAP1-AS1 was upregulated in breast cancer tissues and cells, and AFAP1-AS1 knockdown could hinder proliferation and migration of breast cancer cells, and contribute to cell apoptosis. MiR-497-5p, which was downregulated in breast cancer, was verified to be a target of AFAP1-AS1 and inversely correlated with AFAP1-AS1 expression. SEPT2, as a target gene of miR-497-5p, was negatively regulated by miR-497-5p and positively correlated with AFAP1-AS1 expression. Importantly, AFAP1-AS1 could upregulate SEPT2 expression by sponging miR-497-5p, and modulate cell progression by regulation of the miR-497-5p/SEPT2 axis in breast cancer.
Conclusion:
AFAP1-AS1 knockdown repressed the progression of breast cancer cells by sponging miR-497-5p and downregulating SEPT2.
Introduction
Breast cancer is a type of deadly cancer in women, with a high rate of nearly 29% among all women with cancer. 1,2 The breast cancer was characterized by the high recurrence rate and rapid metastasis. 3 The molecular pathogenesis of breast cancer is also elusive. 4 Recently, gene therapy for cancer has attracted wide attention and become an important means to study cancer. 5 It is an inevitable trend to elucidate the novel gene molecular mechanism related to the development of breast cancer.
Non-coding RNAs (ncRNAs) were expected to be the star factors in the next generation of molecular biology research, 6 including long non-coding RNAs (lncRNAs) and microRNAs (miRNAs). So far, many studies proved that lncRNAs were related to the progress of breast cancer. Han et al. indicated that lncRNA CCAT1 could promote the progression of triple-negative breast cancer. 7 LncRNA H19 knockdown suppressed cell growth and motility by modulating miR-138 in breast cancer. 8 LncRNA NORAD could predict poor prognosis and augment breast cancer development by modulating TGF-β pathway. 9 Actin filament-associated protein1-antisense RNA 1 (AFAP1-AS1) was found in esophageal adenocarcinoma. 10 Recently, many studies revealed that AFAP1-AS1 as an oncogene was exceptionally facilitated in many cancers. It has been found that AFAP1-AS1 accelerated cell progression of cervical cancer, 11 colon cancer, 12 and gastric cancer. 13 Zhang et al. reported that AFAP1-AS1 also promoted the procession of triple-negative breast cancer cells through modulating Wnt/β-Catenin pathway. 14 Moreover, AFAP1-AS1 was confirmed to enhance trastuzumab resistance in breast cancer. 15 However, the molecular mechanism of AFAP1-AS1 as an endogenous RNA of miRNA to regulate breast cancer progression remains unclear.
MiR-497-5p, as a majority of malignant tumor suppressor gene, has been concerned and studied by most scholars. For instance, overexpression of miR-497-5p inhibited the growth of osteosarcoma cells. 16 MiR-497-5p retarded cell progression via regulating SOX5 and FGF2 in non-small-cell lung cancer. 17,18 MiR-497-5p was reported to be downregulated in breast cancer. 19 However, Hu et al. found that miR-497 was significantly increased in oral squamous cell carcinoma and promoted cell metastasis by inhibiting the expression of SMAD7. 20 These results aroused our interest to investigate the effect of miR-497-5p on breast cancer.
Septin 2 (SEPT2) belongs to the septins protein family, which has 14 members. 21 Septins are a class of cytoskeletal proteins that regulate a variety of cellular functions, such as cell migration and apoptosis. 22,23 In the septin family, SEPT2 was thought to be an oncogene for biliary tract cancer 24 and hepatocellular carcinoma. 25 Furthermore, SEPT2 was related to lymph node metastasis, differentiation, and TNM staging in colorectal cancer. 26 SEPT2 was also involved in the development of leukemia as an MLL fusion partner. 27 Additionally, Zhang et al. indicated that SEPT2 could be implicated in the development of breast cancer. 28 However, whether SEPT2 could be modulated by lncRNA and miRNA has never been reported in breast cancer.
The study mainly researched the role of AFAP1-AS1 in the development of breast cancer by modulating miR-497-5p/SEPT2 axis, hoping to find novel markers for the diagnosis of breast cancer.
Materials and Methods
Clinical tissues and cell culture
Twenty breast cancer tissues and the corresponding adjacent normal tissues were harvested from 20 breast cancer patients who were subjected to surgery in Shengli Oilfield Central Hospital. The normal tissues were acquired at least 5 cm away from breast cancer tissues. All patients had not received any treatment before collecting the samples and they had written consent forms. The isolated pieces of tissues were transferred to liquid nitrogen and kept at −80°C. This study was approved by the Ethics Committee of Shengli Oilfield Central Hospital.
HCC70 was acquired from American Type Culture Collection (ATCC; Manassas, VA). Human breast cancer cell lines BT-549, MCF-7, and MDA-MB-231 and breast epithelial cell line MCF-10A were obtained from Procell Company (Wuhan, China). These cells were grown in Roswell Park Memorial Institute-1640 (RPMI-1640) (Sigma-Aldrich, St. Louis, MO) medium with 10% fetal bovine serum (FBS, Sigma-Aldrich) and 2% penicillin-streptomycin at 37°C with 5% CO2.
Quantitative real-time polymerase chain reaction
The RNA of AFAP1-AS1, miR-497-5p, or SEPT2 was extracted by TRIzol reagent (Invitrogen). Reverse transcription kit was purchased from TaKaRa (Wuhan, China). Then, quantitative real-time polymerase chain reaction (qRT-PCR) was performed on ABI 7500 Fast Real-Time PCR system (Applied Biosystems, Carlsbad, CA) using SYBR-Green PCR Master Mix (Thermo Fisher Scientific, Waltham, MA). The levels of AFAP1-AS1 and SEPT2 were standardized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). MiR-497-5p expression was normalized by U6. The relative expression was calculated according to the 2−ΔΔCt method. Sequence of primers: AFAP1-AS1, forward 5′-TCGCTCAATGGAGTGACGGCA-3′; reverse 5′-CGGCTGAGACCGCTGAGAACTT-3′. GAPDH, forward 5′-CACCCACTCCTCCACCTTTG-3′; reverse 5′-CCACCACCCTGTTGCTGTAG-3′. MiR-497-5p, forward 5′-CCTTCAGCAGCACACTGTGG-3′; reverse 5′-CAGTGCAGGGTCCGAGGTAT-3′. U6, forward 5′-CTCGCTTCGGCAGCACA-3′; reverse 5′-AACGCTTCACGAATTTGCGT-3′. SEPT2, forward 5′-GGAGCAGCAGCTTTGAACAC-3′; reverse 5′-TGCAGTTGATAGCGTCACCA-3′.
Cell transfection
Small interfering RNA (siRNA) against AFAP1-AS1 (si-AFAP1-AS1), si-NC, miR-497-5p-mimics, mimics-NC, miR-497-5p-inhibitor, and inhibitor-NC were synthesized by GenePharma (Shanghai, China). The coding sequence (CDS) of SEPT2 was inserted to the pcDNA3.1 vector (Invitrogen, Carlsbad, CA) to enhance SEPT2 expression. The lentivirus for AFAP1-AS1 knockdown (lv-sh-AFAP1-AS1) and negative control (lv-sh-control) was purchased from Wuyuan Company (Beijing, China). Lipofectamine 2000 was used for transfection (Invitrogen).
Cell proliferation assay
For the detection of proliferation, BT-549 and MCF-7 cells were seeded to 96-well plates for 12 h before transfection. At designated times after transfection, 10 μL of Methylthiazolyldiphenyl-tetrazolium bromide (MTT; Thermo Fisher Scientific) was added into the well and maintained for 3 h. After discarding the supernatant, 200 μL of dimethyl sulfoxide (DMSO) was added to solubilize the formazan. Finally, the cell proliferation was examined by measuring the absorbance at 490 nm on a UV microplate reader (Tecan Austria GmbH, Groedig, Austria).
Transwell migration assay
The migration of BT-549 and MCF-7 cells was assessed using Transwell assay without Matrigel. Cells were starved for a day before inoculation. Next, the cells were seeded onto the upper chamber with serum-free medium, and 600 μL RPMI 1640 containing 10% FBS was used to fill the lower chamber. One day after treatment, the cells on the lower surface were dyed with 0.1% crystal violet (Sigma-Aldrich) for 20 min and observed using a microscope.
Cell apoptosis assay
Cells were seeded into six-well plates and placed in an incubator for 12 h, followed by another 48 h after transfection. Then, cells were digested and washed, and double-stained with FITC-Annexin V and propionate iodide (PI) (BD Biosciences, Franklin Lake, NJ) (100 μg/mL) for 15 min in the dark. Finally, cell apoptosis was measured by Flow cytometry assay (BD Biosciences).
Dual-luciferase reporter assay
The putative binding sites between miR-497-5p and AFAP1-AS1 or SEPT2 were predicted by using StarBase v.3.0. Wild AFAP1-AS1 fragment (AFAP1-AS1-WT) containing miR-497-5p binding sites or mutant AFAP1-AS1 fragment (AFAP1-AS1-MUT) without miR-497-5p binding sites was cloned into the pmirGLO vector (EK-bioscience, Shanghai, China). Subsequently, luciferase reporter plasmids were transfected into breast cancer cells together with miR-497-5p-mimics or mimics-NC. The luciferase activity was measured by a dual-luciferase reporter kit (Solarbio, Beijing, China). Wild SEPT2-3′UTR fragment (SEPT2-WT) or mutant SEPT2-3′UTR fragment (SEPT2-MUT) containing miR-497-5p binding sites or not was cloned into the pmirGLO vector. The rest of the process was the same as described above.
Western blot assay
Proteins from BT-549 and MCF-7 cells and tumor tissues of nude mice were extracted through RIPA reagent (Solarbio). Proteins were then separated and transferred to polyvinylidene difluoride (PVDF) membranes (Thermo Fisher Scientific). After blockage with 5% milk powder at 37°C for 2 h, the membranes were incubated with the primary antibody of SEPT2 (dilution: 1: 1000; Thermo Fisher Scientific) or GAPDH (dilution: 1: 2000; Santa Cruz Biotechnology, CA) at 4°C for 12 h. Then, the membranes were washed and incubated with the secondary antibodies (dilution: 1: 3000; Thermo Fisher Scientific) for 1 h. The membranes were visually detected by the ECL assay (Thermo Fisher Scientific).
Animal experiment
Four-week-old female nude mice were applied for the study. The animal experiments were permitted by the Animal Ethics Committee of Shengli Oilfield Central Hospital. After transfection with lv-sh-AFAP1-AS1 or lv-sh-control, BT-549 cells were injected into the abdomen of mice. Tumor volume was surveyed by caliper once a week for 4 weeks. Finally, all the mice were euthanized and tumors were weighed. Relative expression levels of AFAP1-AS1, miR-497-5p, and SEPT2 in the resected tumors were measured.
Statistical analysis
The differences between the two experimental groups were analyzed by Student's t-test. Pearson's correlation coefficient was used to analyze the correlations. Data in this study were expressed as mean ± standard deviation (SD) and repeated at least three times separately. p-Value <0.05 was considered statistically significant.
Results
AFAP1-AS1 was upregulated in breast cancer tissues and cells
To explore whether AFAP1-AS1 contributed to the development of breast cancer, we examined the expression level of AFAP1-AS1 in breast cancer tissues by qRT-PCR. As shown in Figure 1A, AFAP1-AS1 was significantly increased in breast cancer tissues compared to those in normal tissues (n = 20). Meanwhile, we found that the expression of AFAP1-AS1 in positive tissues was enormously high (Fig. 1B). Then, AFAP1-AS1 expression was detected in human breast epithelial cell line (MCF-10A) and human breast cancer cell lines (BT-549, MCF-7, MDA-MB-231, and HCC70). The results showed that AFAP1-AS1 expression was also drastically enhanced in breast cancer cell lines (Fig. 1C). Results showed that patients with high expression of AFAP1-AS1 had short survival time (Fig. 1D). The data suggested that AFAP1-AS1 might be an oncogene in breast cancer.
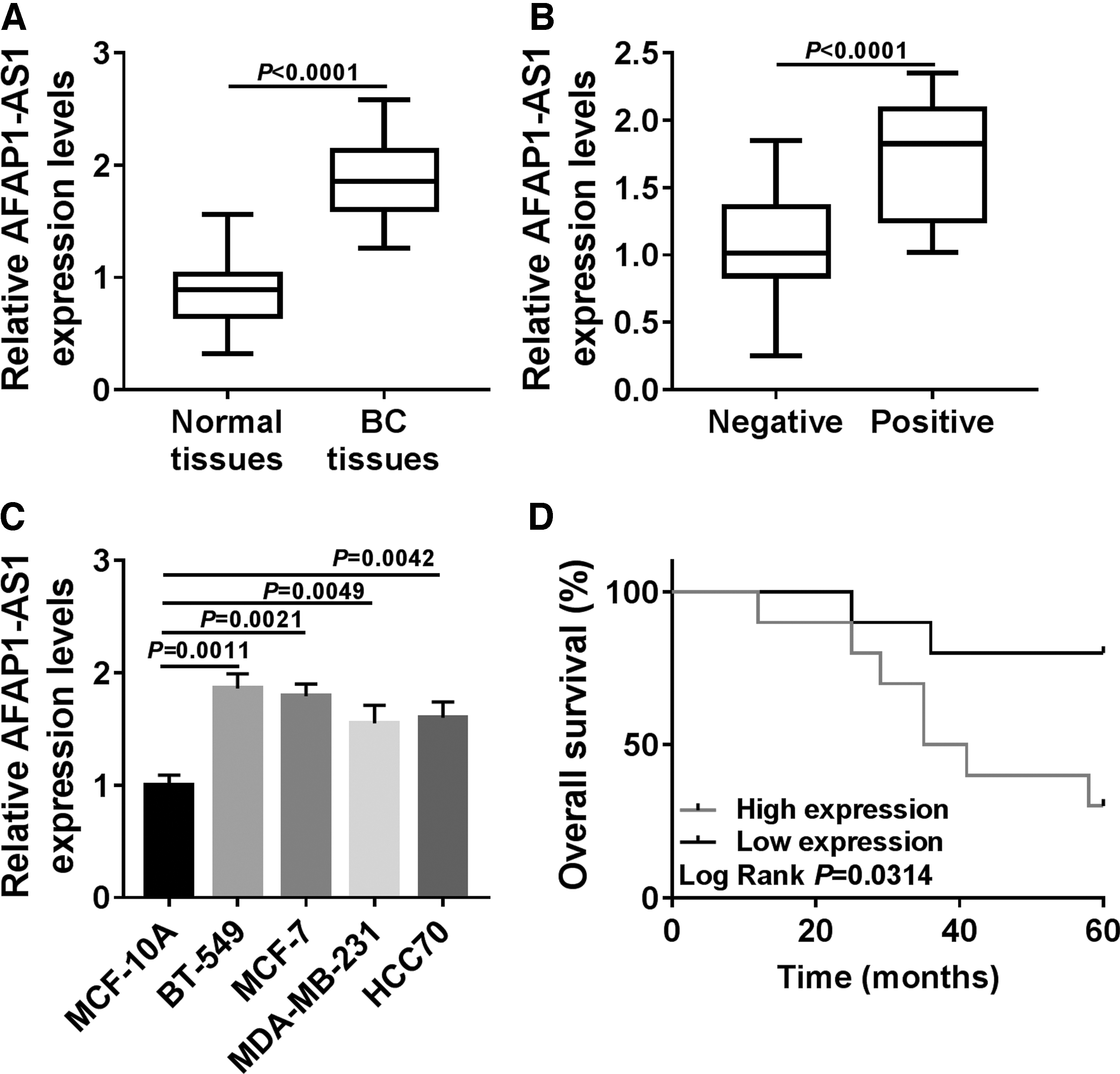
AFAP1-AS1 was upregulated in breast cancer tissues and cell lines.
Silencing AFAP1-AS1 inhibited proliferation and migration of breast cancer cells and led to apoptosis in vitro
Considering the high expression of AFAP1-AS1 in breast cancer, we examined the effect of AFAP1-AS1 knockdown on the biological behaviors of breast cancer cells. First, we interfered with AFAP1-AS1 in BT-549 and MCF-7 cells by transfecting si-AFAP1-AS1. As shown in Figure 2A and B, siRNAs targeting AFAP1-AS1 significantly reduced the expression level of AFAP1-AS1 in BT-549 and MCF-7 cells. Subsequently, MTT assay demonstrated that AFAP1-AS1 knockdown markedly impaired the proliferation of BT-549 and MCF-7 cells (Fig. 2C, D). Transwell assay indicated that AFAP1-AS1 knockdown attenuated the migration capacity of BT-549 and MCF-7 cells, and significantly decreased the number of cell migrations (Fig. 2E). Besides, we found that knockdown of AFAP1-AS1 could significantly increase the apoptosis rate of BT-549 and MCF-7 cells (Fig. 2F). We found a successful overexpression efficiency of AFAP1-AS1 in both BT-549 and MCF-7 cells (Supplementary Fig. S1A). Moreover, AFAP1-AS1 overexpression promoted cell proliferation, migration, and inhibited cell apoptosis (Supplementary Fig. 1B–E). In short, AFAP1-AS1 knockdown suppressed proliferation and migration of breast cancer cells, and promoted cell apoptosis in vitro.

Effect of AFAP1-AS1 on progression of breast cancer cells.
AFAP1-AS1 served as a ceRNA for miR-497-5p in breast cancer cells
LncRNAs have been shown to sponge miRNAs and modulate gene expression. 29 To test whether AFAP1-AS1 has a similar mechanism in breast cancer cells, we used StarBase v.3.0 to predict potential miRNAs that could bind to AFAP1-AS1. As shown in Figure 3A, AFAP1-AS1 and miR-497-5p had the potential binding sites and the dual-luciferase reporter assay further demonstrated the targeted relationship between them. The results showed that the luciferase activity in BT-549 and MCF-7 cells co-transfected with AFAP1-AS1-WT and miR-497-5p-mimics was significantly declined compared with that in cells co-transfected with AFAP1-AS1-WT and mimics-NC, and there was no distinct change in the luciferase activity of cells transfected with AFAP1-AS1-MUT (Fig. 3B, C). Simultaneously, we detected the expression of miR-497-5p in BT-549 and MCF-7 cells transfected with si-AFAP1-AS1. The data showed that AFAP1-AS1 knockdown steeply augmented the expression of miR-497-5p (Fig. 3D). These results further clarified that AFAP1-AS1 could target miR-497-5p. Thus, we explored the role of miR-497-5p in breast cancer. QRT-PCR assay showed that miR-497-5p was markedly constrained in breast cancer tissues and cell lines (Fig. 3E, F). Furthermore, miR-497-5p expression was negatively associated with the expression of AFAP1-AS1 in breast cancer tissues (r = −0.7744, p < 0.0001) (Fig. 3G). The results identified that miR-497-5p inhibitor could restore the inhibition of si-AFAP1-AS1 on the proliferation and migration of BT-549 and MCF-7 cells, and could also overturn the promotion of si-AFAP1-AS1 on apoptosis (Fig. 3H–K and Supplementary Fig. S2A-B). These findings strongly indicated that AFAP1-AS1 served as an miRNA decoy for miR-497-5p and implicated in the progression of breast cancer cells by regulating miR-497-5p.

Studies on the relationship between AFAP1-AS1 and miR-497-5p in breast cancer cells.
SEPT2 was directly targeted by miR-497-5p in breast cancer cells
To further explore the downstream mechanism of AFAP1-AS1/miR-497-5p, we looked for the target genes of miR-497-5p. StarBase v.3.0 predicted that there were binding sites between miR-497-5p and SEPT2-3′ UTR (Fig. 4A). Dual-luciferase reporter assay indicated that miR-497-5p-mimics significantly impaired the luciferase activity in BT-549 and MCF-7 cells transfected with SEPT2-WT compared with the same cells transfected with SEPT2-MUT (Fig. 4B, C). In addition, the mRNA and protein levels of SEPT2 in BT-549 and MCF-7 cells were significantly reduced by miR-497-5p-mimics, while miR-497-5p-inhibitor significantly induced the mRNA and protein levels of SEPT2 (Fig. 4D, E). Then, we analyzed the expression of SEPT2 in breast cancer tissues and found that SEPT2 expression was strikingly upregulated in breast cancer tissues compared with normal tissues (Fig. 4F), and the mRNA and protein levels of SEPT2 were also significantly increased in breast cancer cells (Fig. 4G, H). Moreover, there was a negative relationship between the expression of miR-497-5p and SEPT2 in breast cancer tissues (r = −0.706, p < 0.0001) (Fig. 4I). To understand whether SEPT2 could affect the regulation of biological behavior in breast cancer cells by miR-497-5p, SEPT2 was overexpressed in BT-549 and MCF-7 cells. The mRNA and protein levels of SEPT2 were distinctly upregulated in BT-549 and MCF-7 cells transfected with pcDNA3.1-SEPT2 (Fig. 4J, K). The results of the restoration experiment showed that miR-497-5p-mimics could inhibit the proliferation and migration of BT-549 and MCF-7 cells and promote apoptosis, while these effects could be recovered by overexpression of SEPT2 (Fig. 4L–O and Supplementary Fig. S3A-B). These results suggested that miR-497-5p directly targeted and regulated SEPT2, and SEPT2 could regain the inhibition of miR-497-5p on breast cancer cell development.

Identification of the targeting relationship between miR-497-5p and SEPT2 in breast cancer cells.
AFAP1-ASl upregulated the expression of SEPT2 by acting as a ceRNA of miR-497-5p in breast cancer cells
In this part, we aimed to explore whether AFAP1-AS1 could regulate SEPT2 expression through miR-497-5p in breast cancer cells. The data indicated that the declined mRNA and protein expression levels of SEPT2 in BT-549 and MCF-7 cells caused by si-AFAP1-AS1 could be recovered by miR-497-5p inhibitor (Fig. 5A–D). Moreover, overexpression of SEPT2 in BT-549 and MCF-7 cells could invert the inhibitory effect of si-AFAP1-AS1 on cell proliferation and migration (Fig. 5E–G), and alleviate the promoting effect of si-AFAP1-AS1 on cell apoptosis (Fig. 5H). Collectively, these results revealed that AFAP1-AS1 modulated the expression of SEPT2 by serving as a ceRNA for miR-497-5p in breast cancer cells.

AFAP1-AS1 upregulated the expression of SEPT2 by modulating miR-497-5p in breast cancer cells.
AFAP1-ASl knockdown inhibited tumor growth by regulating the miR-497-5p/SEPT2 axis in vivo
Next, we established the nude mouse model to study the effect of AFAP1-AS1 on tumor growth in vivo. The data showed that tumor growth in nude mice was significantly hindered after subcutaneous inoculation of BT-549 cells transfected with lv-sh-AFAP1-AS1, which was reflected by the reduction of tumor volume and weight (Fig. 6A, B). Furthermore, the expression levels of AFAP1-AS1, miR-497-5p, and SEPT2 in isolated tumor tissues were examined. In the lv-sh-AFAP1-AS1 group, AFAP1-AS1 was dramatically declined and miR-497-5p was remarkably elevated compared with that in lv-sh-control group (Fig. 6C, D). The mRNA and protein levels of SEPT2 were significantly decreased in the lv-sh-AFAP1-AS1 group (Fig. 6E, F). The above results implied that AFAP1-AS1 knockdown could impede tumor growth in vivo by modulating the miR-497-5p/SEPT2 axis.

Discussion
AFAP1-AS1 was one of the classical lncRNAs and played a pivotal role in the development of various cancers. 30 Li et al. indicated that AFAP1-AS1 augmented cell growth and invasion in osteosarcoma. 31 Besides, AFAP1-AS1 could also promote the progression of gastric cancer. 32 These data confirmed that AFAP1-AS1 was positively associated with the proliferation and migration of cancer cells. This positive correlation was due to the high expression of AFAP1-AS1 in cancers. AFAP1-AS1 has been reported as an oncogene with high expression in gastric cancer, 33 retinoblastoma, 34 and breast cancer. 35 However, the molecular mechanism of AFAP1-AS1 in breast cancer progression remains largely unclear.
Here, we found that AFAP1-AS1 was strikingly aggrandized in breast cancer. Additionally, interference with AFAP1-AS1 impaired proliferation and migration of breast cancer cells, and elevated cell apoptosis rate in vitro, and retarded the growth of tumor in vivo. These results were consistent with previous data. 36
Recently, the competitive endogenous RNAs (ceRNAs) theory revealed a novel regulatory mode of lncRNAs. 37 Wu et al. indicated that AFAP1-AS1 regulated the expression of ACVR1 by competitively binding with miR-384 as a ceRNA in pancreatic cancer. 38 Therefore, we intended to construct an AFAP1-AS1-miRNA-mRNA network to better understand the regulatory effect of AFAP1-AS1 on breast cancer. MiR-497-5p was confirmed to be directly targeted by AFAP1-AS1 and their expression in breast cancer tissues was inversely correlated. In contrast to the AFAP1-AS1, miR-497-5p was significantly dwindled in breast cancer, which was in accordance with previous results in breast cancer. 19 Besides, AFAP1-AS1 could negatively regulate miR-497-5p expression in breast cancer cells, and interfering with miR-497-5p could alleviate the suppressive effects of si-AFAP1-AS1 on the proliferation and migration of breast cancer cells, and restore the promoting effect of si-AFAP1-AS1 on apoptosis. These results implied that AFAP1-AS1 induced breast cancer development by functioning as a sponge of miR-497-5p. Lian et al. reported that AFAP1-AS1 could promote the metastasis of nasopharyngeal carcinoma as a ceRNA of miR-423-5p. 39
Subsequently, SEPT2, a gene targeted by miR-497-5p, attracted our attention. It has been widely reported to be upregulated in many cancers including breast cancer. 28,40 In our study, SEPT2 was enormously promoted in breast cancer. Similarly, SEPT2 expression in breast cancer cells was inversely regulated by miR-497-5p and a distinct negative correlation between them was found in breast cancer tissues. Moreover, upregulation of SEPT2 partially regained the suppressive impacts of miR-497-5p-mimics on proliferation and migration of breast cancer cells and the promotion effect on apoptosis, which was consistent with the previous results that overexpression of SEPT2 could promote the process of breast cancer cells. 28 To justify the rationality of AFAP1-AS1/miR-497-5p/SEPT2 mechanism in breast cancer, some recovery experiments were carried out. We demonstrated that AFAP1-AS1 not only upregulated the expression of SEPT2 by sponging miR-497-5p but also promoted the progression of breast cancer cells via modulating SEPT2 in vitro. More importantly, knockdown of AFAP1-AS1 could impede tumor growth via the miR-497-5p/SEPT2 axis in vivo. These results confirmed our idea that the miR-497-5p/SEPT2 regulatory network controlled by AFAP1-AS1 could play a role in the development of breast cancer.
Overall, we revealed that interference with AFAP1-AS1 inhibited the progression of breast cancer via modulating SEPT2 by competitively binding with miR-497-5p. These results implied that AFAP1-AS1 might be a new target for the treatment of breast cancer as an oncogene.
Footnotes
Acknowledgment
The authors sincerely appreciate all members who participated in this study.
Disclosure Statement
The authors have declared that they have no conflicts of interest.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.