
Abstract
Background:
Long noncoding RNA (lncRNA) small nucleolar RNA host gene 6 (SNHG6) has been reported to be an oncogene in a variety of cancers. However, the role of SNHG6 and its associated mechanisms in Wilms' tumor progression remain largely unknown.
Methods:
The expression of SNHG6, microRNA-429 (miR-429), and FGF receptor substrates 2 (FRS2) messenger RNA (mRNA) was detected by quantitative real-time polymerase chain reaction. Cell proliferation was analyzed through 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay and plate colony assay. The apoptosis was assessed by flow cytometry. Cell glycolytic metabolism was analyzed through detecting the lactate dehydrogenase activity, glucose uptake, lactate production, and ATP level. The target relationship between miR-429 and SNHG6 or FRS2 was predicted by miRcode or Starbase and then validated by dual-luciferase reporter assay and RNA pull-down assay. Murine xenograft model was established to validate the function of SNHG6 in vivo.
Results:
The level of SNHG6 was elevated in Wilms' tumor tissues and cells, and SNHG6 played an oncogenic role to promote the proliferation and glycolysis and restrain the apoptosis of Wilms' tumor cells. MiR-429 was identified as a target of SNHG6, and miR-429 interference partly reversed the inhibitory effects induced by SNHG6 silencing on the malignant behaviors of Wilms' tumor cells. FRS2 mRNA bound to miR-429 in Wilms' tumor cells. SNHG6 upregulated the expression of FRS2 through acting as a sponge of miR-429. MiR-429-induced influences in Wilms' tumor cells were largely counteracted by the overexpression of FRS2. SNHG6 silencing suppressed the Wilms' tumor growth through miR-429/FRS2 axis in vivo.
Conclusion:
SNHG6 accelerated Wilms' tumor progression through regulating miR-429/FRS2 signaling in vitro and in vivo.
Introduction
Wilms' tumor commonly occurs in the urinary system of children. The combined therapy of surgical resection, chemo-, and radiotherapy has improved the 5-year survival rate of patients with Wilms' tumor, 1 but there are still some children who died because of metastasis, recurrence, or other reasons. WT1 and P53 were implicated in the development of Wilms' tumor. 2,3 However, the regulatory mechanisms behind Wilms' tumor progression remain poorly understood. Therefore, uncovering the molecular mechanisms behind Wilms' tumor progression and finding novel therapeutic targets are crucial for the treatment of Wilms' tumor.
Long noncoding RNAs (lncRNAs) are a group of RNAs that include 200 nucleotides at least, and they are generally unable to code proteins. LncRNAs possess binding sites with microRNAs (miRNAs), thus they are also known as miRNA sponges. 4 –7 Small nucleolar RNA host gene 6 (SNHG6) has been reported to facilitate Wilms' tumor progression through sponging miR-15a. 8 In this study, the authors identified another signal pathway by which SNHG6 promoted the progression of Wilms' tumor.
MicroRNAs (miRNAs) bind to messenger RNAs (mRNAs) through their “seed” sites to downregulate or impair the translation of these target mRNAs. MiRNAs could regulate diverse cellular behaviors, including cell proliferation and metastasis. 9 Emerging evidences suggested that miR-429 was involved in the cell proliferation and apoptosis of Wilms' tumor. 10 However, the underlying signal regulatory network of miR-429 in Wilms' tumor progression is not fully addressed.
They first assessed functions of SNHG6 in Wilms' tumor progression through conducting loss-of-function experiments. Subsequently, SNHG6/miR-429/FRS2 axis was identified to disclose the molecular mechanism by which SNHG6 functioned in Wilms' tumor.
Materials and Methods
Tissue specimen collection
A total of 25 pairs of Wilms' tumor specimens and their matching adjacent nontumor specimens were obtained from patients with Wilms' tumor at The Second Hospital of Dalian Medical University. Tissue samples were immediately stored in −80°C after surgical resection until used. This research was performed with permission of the Ethics Committee of The Second Hospital of Dalian Medical University, and each subject had submitted informed consent before the resection operation. The correlation between SNHG6 expression and pathological characteristics in patients with Wilms' tumor is shown in Table 1.
Correlation Between SNHG6 Expression and Pathological Characteristics in Patients with Wilms' Tumor (n = 25)
p < 0.05.
Using median expression level of SNHG6 as cutoff.
Cell lines
Human embryonic kidney cell line HEK293T was purchased from American Tissue Culture Collection (Manassas, VA), and HEK293T was cultivated with Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS; Gibco). Human Wilms' tumor cell lines, WiT-49 and GHINK-1 (BeNa Culture Collection, Beijing, China), were cultivated with DMEM/nutrient mixture F-12 (Gibco) (1:1) containing 10% FBS and Maccyo’5 medium (Invitrogen, Carlsbad, CA) added with 15% FBS, respectively. Ten percent penicillin/streptomycin mixture was added into the culture medium of all the above cell lines.
Quantitative real-time polymerase chain reaction
SYBR green and primers (GeneCopoeia) were used in amplification reaction. The enrichment of SNHG6, miR-429, and FRS2 was examined by the 2−ΔΔCt method. 11 The primer sequences are shown in Table 2. U6 (for miR-429) or glyceraldehyde-3-phosphate dehydrogenase (GAPDH; for SNHG6 and FRS2) acted as the housekeeping gene in this study.
Primer Sequences for Quantitative Real-Time Polymerase Chain Reaction
Cell transfection
SNHG6 small interfering RNAs (si-SNHG6#1 and si-SNHG6#2) and siRNA negative control (si-NC), SNHG6 overexpression plasmid (pcDNA-SNHG6), FRS2 overexpression plasmid (pcDNA-FRS2), pcDNA-NC, short hairpin RNA against SNHG6 (sh-SNHG6), and sh-NC were obtained by GenePharma (Shanghai, China). MiR-429 mimics (miR-429), miR-NC, miR-429 inhibitor (anti-miR-429), and anti-miR-NC were obtained from Ribobio (Guangzhou, China). Transfection was performed using Lipofectamine™ 3000 (Thermo Fisher Scientific, Shanghai, China).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
After transfection, MTT (10 μL, 5 mg/mL; Invitrogen) was added to the wells of cell culture plate. Following 4-h incubation, the cell supernatant was discarded. One hundred microliters of dimethyl sulfoxide (DMSO; Sigma, St. Louis, MO) was added to the above wells to dissolve the formazan. The optical density value was measured by a microplate reader at 570 nm.
Plate colony assay
Transfected WiT49 and GHINK-1 cells were replated into six-well cell culture plates (200 cells/well), and the cell culture plates were continued to culture for 2 weeks. During this period, cell culture medium supplemented with serum was refreshed every 3 d. Cell colonies were stained with sulforhodamine B, and the number of colonies was counted.
Cell apoptosis analysis
Cells were collected and simultaneously dyed with 5 μL Annexin V-combined fluorescein isothiocyanate (FITC; Solarbio, Beijing, China) and propidium iodide (PI; Solarbio) in a dark atmosphere for 10 min after transfection for 72 h. The apoptotic cells (FITC+ and PI+/−) were recognized and counted by the flow cytometer.
Western blot assay
The total proteins were separated through 12% separating gel and then transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA). After 1-h blocking, the PVDF membranes were incubated with primary antibody against B cell lymphoma 2 (Bcl-2; ab185002, Abcam, Cambridge, MA), Bcl-2 Associated X Protein (Bax; ab32503; Abcam), FRS2 (ab193363; Abcam), or GAPDH (ab37168; Abcam) overnight at 4°C, and then the membranes were incubated with horse radish peroxidase-labeled secondary antibody. Protein bands were detected through the Enhanced Chemiluminescent System Kit (Beyotime, Shanghai, China).
Glycolysis analysis
The glycolysis of Wilms' tumor cells was analyzed through measuring the lactate dehydrogenase (LDH) activity, the uptake of glucose, and the production of lactate and ATP using the LDH Activity Assay Kit (Sigma), Glucose Assay Kit (BioVision, Milpitas, CA), Lactate Assay Kit (Sigma), and Firefly Luciferase-Based ATP Assay Kit (Beyotime), respectively.
Bioinformatic prediction
The interacted miRNAs of SNHG6 were predicted by miRcode database, while Starbase database was used to predict the interacted mRNAs of miR-429.
Dual-luciferase reporter assay
To verify the combination between miR-429 and SNHG6, the sequence of SNHG6, including wild-type (WT)-binding sites (5′-AGUAUU-3′) or mutant-binding sites (5′-GACAGC-3′) with miR-429, was amplified and inserted to the luciferase gene downstream of pmirGLO vector (Promega, Madison, WI), named as WT-SNHG6 or MUT-SNHG6, respectively. Cells were cotransfected with WT-SNHG6 or MUT-SNHG6 and miR-429 or miR-NC. Luciferase activity was analyzed after 48-h transfection using the Dual-Luciferase Reporter Assay Kit (Promega). The binding between miR-429 and FRS2 mRNA was similarly validated.
RNA pull-down assay
RNA pull-down assay was used to confirm the target relationship between miR-429 and SNHG6 or FRS2 in Wilms' tumor cells. MiR-429 was biotinylated to generate Bio-miR-429. Meanwhile, the binding sites with SNHG6 or FRS2 in miR-429 were mutated, and Bio-miR-429-MUT was obtained through biotinylation. Wilms' tumor cells were transfected with Bio-miR-NC, Bio-miR-429, or Bio-miR-429-MUT for 48 h. Subsequently, Wilms' tumor cells were lysed and incubated with Dynabeads M-280 Streptavidin (Invitrogen). The abundance of SNHG6 and FRS2 mRNA was detected by quantitative real-time polymerase chain reaction (qRT-PCR).
Murine xenograft assay
Nude mice xenograft experiments were permitted by the Animal Research Committee of The Second Hospital of Dalian Medical University. BALB/c nude mice (Orient Bio, Inc., Seongnam, South Korea) were randomly grouped into sh-NC group (n = 6) and sh-SNHG6 group (n = 6). The right flank of the nude mice was subcutaneously injected with WiT49 cells (2 × 106 cells) stably transfected with sh-NC or sh-SNHG6, and the tumor volume was calculated every week with the formula of volume = (width 2 × length)/2. After 5 weeks following the inoculation, the weight of resected tumors was recorded. Tumor tissues were used to detect the levels of SNHG6, miR-429, and FRS2 protein.
Statistical analysis
All data involved in this study were displayed in the form of mean ± SD from three independent experiments. The comparisons were assessed by Student's t-test and one-way analysis of variance followed by Tukey's test. Spearman's coefficient was utilized to evaluate the liner correlation relationship. p-Value <0.05 was considered as statistical significance.
Results
SNHG6 level is aberrantly elevated in Wilms' tumor tissues and cells
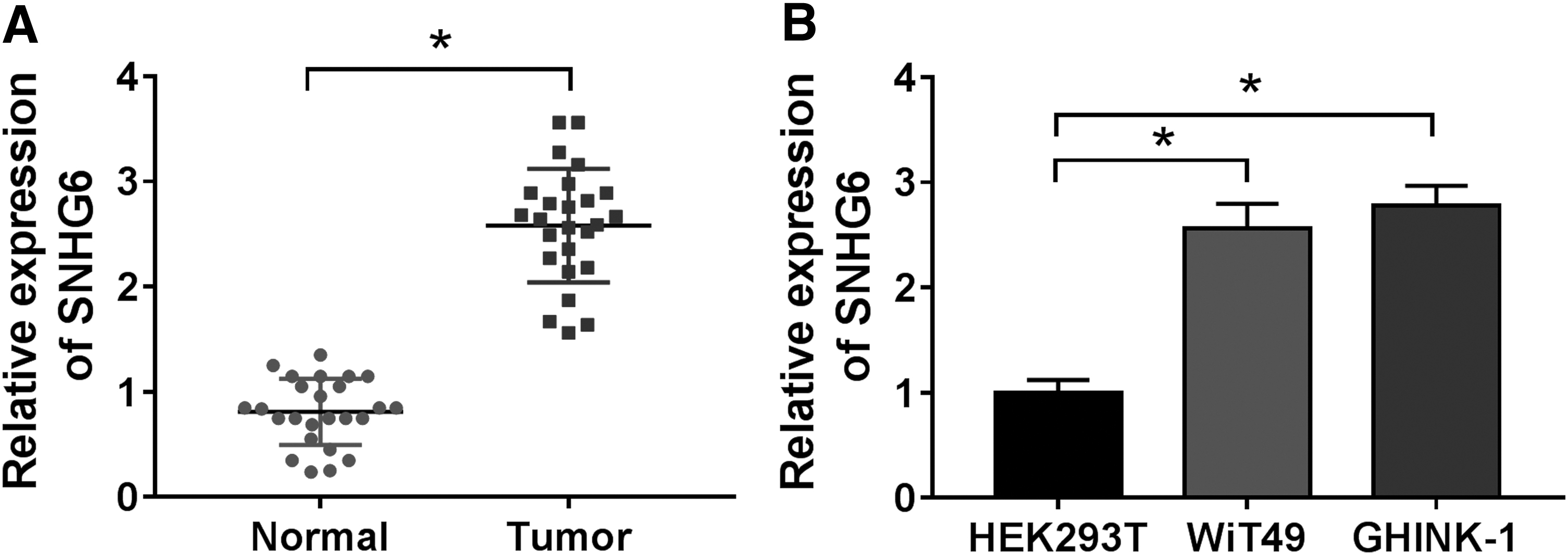
To explore the biological role of lncRNA SNHG6 in Wilms' tumor, qRT-PCR was conducted to evaluate the expression of SNHG6 in Wilms' tumor specimens (n = 25) and two Wilms' tumor cell lines along with paired nontumor specimens (n = 25) and human embryonic kidney cell line HEK293T. The results demonstrated that the abundance of SNHG6 was significantly enhanced in Wilms' tumor tissues and cell lines (Fig. 1A, B).
SNHG6 level is aberrantly elevated in Wilms' tumor tissues and cells.
Furthermore, they assessed the correlation between SNHG6 expression and pathological characteristics in patients with Wilms' tumor. Patients with Wilms' tumor were divided into high SNHG6 group (n = 12) and low SNHG6 group (n = 13) using the median expression level of SNHG6 as cutoff. High expression of SNHG6 was correlated with unfavorable histopathological type, positive lymphatic metastasis, and advanced tumor stage in patients with Wilms' tumor (Table 1). The abnormal expression of SNHG6 might imply its crucial role in Wilms' tumor.
SNHG6 promotes the proliferation and glycolysis while inhibits the apoptosis of Wilms' tumor cells
To investigate the biological functions of SNHG6 in Wilms' tumor cells, si-SNHG6#1 and si-SNHG6#2 were designed to knockdown SNHG6 in WiT49 and GHINK-1 cells. The knockdown efficiencies of these two siRNAs were assessed by qRT-PCR. SNHG6 abundance was significantly decreased with the transfection of si-SNHG6#1 or si-SNHG6#2 compared with si-NC group (Fig. 2A). Si-SNHG6#2 was selected for loss-of-function experiments since its higher knockdown efficiency than si-SNHG6#1.

SNHG6 promotes the proliferation and glycolysis while inhibits the apoptosis of Wilms' tumor cells.
The influence of SNHG6 interference on cell proliferation was assessed through MTT assay and plate colony assay. Cell proliferation curve was generated through analyzing the numbers of viable cells in the two groups after transfection for 0, 24, 48, or 72 h. The results suggested that SNHG6 silencing suppressed the proliferation of Wilms' tumor cells (Fig. 2B, C). Furthermore, the number of colonies was notably decreased with the interference of SNHG6 (Fig. 2D), which further demonstrated that SNHG6 knockdown inhibited the proliferation ability of Wilms' tumor cells.
Cell apoptosis was analyzed through counting the percentages of early and late apoptotic cells through flow cytometry and detecting the expression of apoptosis-related proteins (Bcl-2 and Bax) through Western blot assay. SNHG6 knockdown elevated the apoptosis rate of Wilms' tumor cells (Fig. 2E). Furthermore, the results of Western blot demonstrated that the level of antiapoptotic protein Bcl-2 was decreased, while the enrichment of proapoptotic protein Bax was elevated in si-SNHG6#2-transfected WiT49 and GHINK-1 cells (Fig. 2F, G). These findings together suggested that SNHG6 silencing triggered the apoptosis of Wilms' tumor cells.
Warburg effect is an important hallmark for cancer cells. Cancer cells exhibit altered metabolic phenotype that featured by increased glycolytic metabolism and decreased oxidative phosphorylation even with enough oxygen. 12 Warburg effect provides growth advantages for cancer cells. Apart from obtaining energy from glycolysis, cancer cells also benefit from the reduced production of reactive oxygen species in mitochondria.
In this study, the authors explored if SNHG6 silencing regulated the glycolytic metabolism of Wilms' tumor cells through detecting the LDH activity, the utilization of glucose, and the production of lactate and ATP. SNHG6 silencing decreased LDH activity and suppressed the glucose consumption and lactate production (Fig. 2H–J). Moreover, ATP level was notably decreased with the silencing of SNHG6 in Wilms' tumor cells (Fig. 2K). These results demonstrated that SNHG6 interference hampered the glycolysis of Wilms' tumor cells (Fig. 2H–K). Taken together, SNHG6 facilitated the proliferation and glycolysis and impeded the apoptosis of Wilms' tumor cells.
LncRNA SNHG6 is a molecular sponge for miR-429
To explore the molecular mechanisms by which SNHG6 modulated the biological functions of Wilms' tumor cells, they aimed to find the downstream components of SNHG6. miRcode online software was used to predict potential miRNA targets of SNHG6. As shown in Figure 3A, among 10 candidate miRNA targets of SNHG6, the expression of miR-139-5p, miR-144, miR-429, miR-22-3p, and miR-204 was upregulated with the silencing of SNHG6 in WiT49 cells. Besides, after silencing SNHG6, the levels of miR-144, miR-429, miR-22-3p, and miR-204 were elevated in GHINK-1 cells (Fig. 3B). MiR-429 attached their attention due to its most obvious negative regulatory relationship with SNHG6 in the two Wilms' tumor cell lines. The putative binding sites between SNHG6 and miR-429 predicted by miRcode database are shown in Figure 3C.

LncRNA SNHG6 is a molecular sponge for miR-429.
To test if these putative sites were essential for the target interaction between SNHG6 and miR-429, they mutated the above sites by “GACAGC.” Dual-luciferase reporter assay was performed to confirm the target relationship between SNHG6 and miR-429. SNHG6 sequence containing wild-type or mutant-type binding sites with miR-429 was cloned into pmirGLO vector to generate WT-SNHG6 or MUT-SNHG6 reporter vector, respectively. Luciferase activity was measured in WiT49 and GHINK-1 cells cotransfected with WT-SNHG6 or MUT-SNHG6 and miR-NC or miR-429.
Overexpression of miR-429 notably reduced the luciferase activity in WT-SNHG6 group compared with the MUT-SNHG6 group, indicating that miR-429 was a direct target of SNHG6 in Wilms' tumor cells (Fig. 3D, E). Furthermore, luciferase activity remained unchanged in MUT-SNHG6 group when cotransfected with miR-NC or miR-429 (Fig. 3D, E), suggesting that the predicted binding sites were indeed the binding sites between SNHG6 and miR-429.
Besides, RNA pull-down assay was also conducted to verify the interaction between miR-429 and SNHG6 in Wilms' tumor cells. WiT49 and GHINK-1 cells were transfected with Bio-miR-NC, Bio-miR-429, or Bio-miR-429-MUT. As shown in Figure 3F, SNHG6 was pulled down in Bio-miR-429-transfected group compared with Bio-miR-NC or Bio-miR-429-MUT group, suggesting the interaction between miR-429 and SNHG6 in Wilms' tumor cells.
To investigate the modulatory relationship between miR-429 and SNHG6, qRT-PCR was conducted to detect the level of miR-429 in WiT49 and GHINK-1 cells transfected with pcDNA-NC, SNHG6 overexpression plasmid (pcDNA-SNHG6), si-NC, or si-SNHG6. The abundance of miR-429 was decreased with the overexpression of SNHG6, while SNHG6 knockdown enhanced the level of miR-429 (Fig. 3G), suggesting that there was negative regulatory relationship between miR-429 and SNHG6 in Wilms' tumor cells.
Subsequently, the authors measured the expression pattern of miR-429 in Wilms' tumor. As indicated in Figure 3H and I, the level of miR-429 was reduced in Wilms' tumor tissues and cell lines compared with that in adjacent normal tissues and human embryonic kidney cell line HEK293T, which showed an inverse trend with the expression of SNHG6. The opposite expression trend between SNHG6 and miR-429 (shown in Fig. 3G) in Wilms' tumor also confirmed the negative regulatory relationship between miR-429 and SNHG6. The results of linear correlation analysis showed that the expression of miR-429 was negatively correlated with the level of SNHG6 (Fig. 3J). Taken together, these data revealed that SNHG6 downregulated miR-429 level through directly binding to it in Wilms' tumor cells.
SNHG6 silencing-mediated suppressive effects on the progression of Wilms' tumor are partly reversed by the addition of anti-miR-429
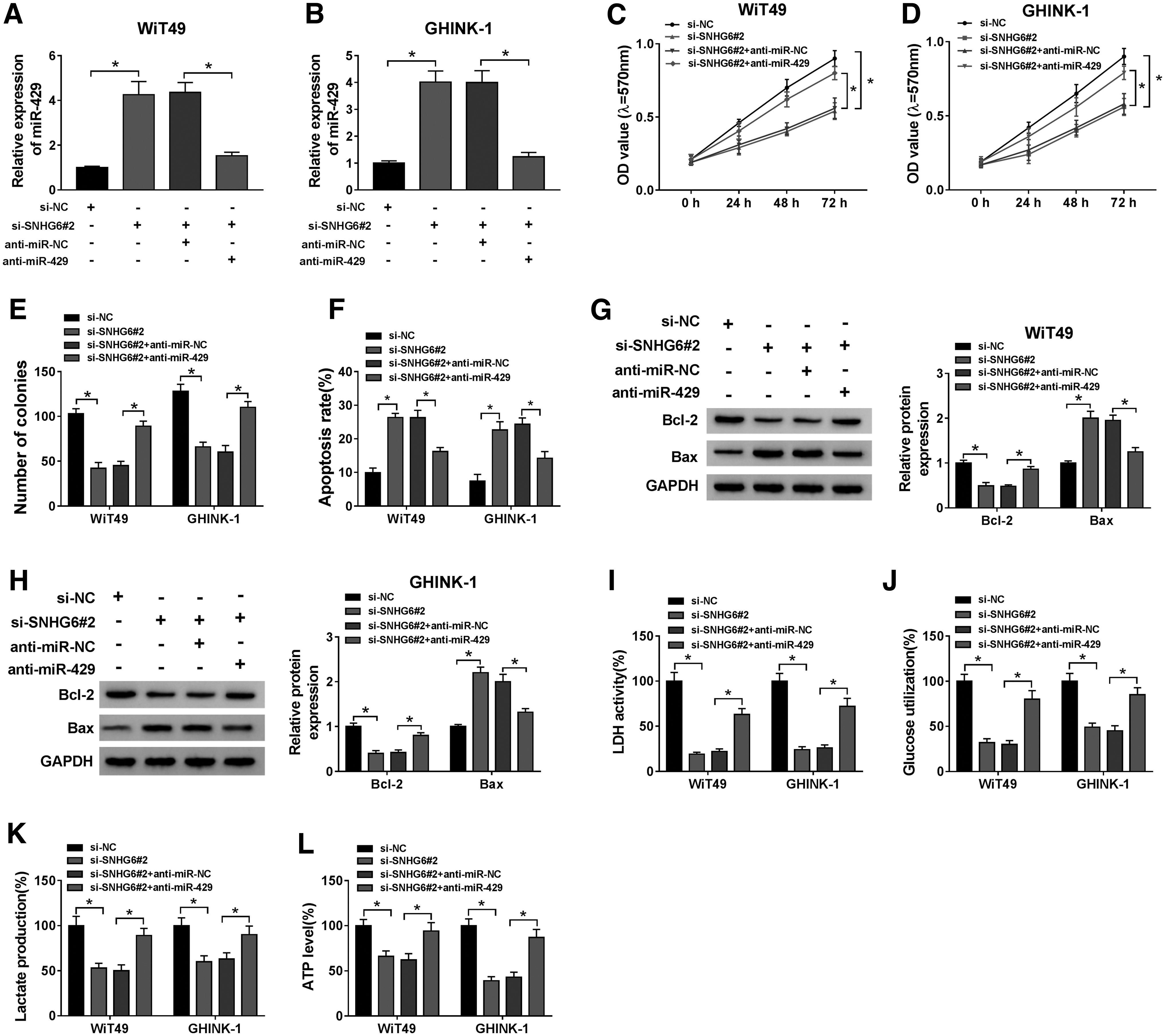
To test if SNHG6 functioned through targeting miR-429, the authors conducted rescue experiments through transfecting si-SNHG6 alone or together with anti-miR-429 into Wilms' tumor cells. The expression of miR-429 was detected by qRT-PCR. As indicated in Figure 4A and B, the relative expression of miR-429 was enhanced with the silencing of SNHG6 in Wilms' tumor cells, and miR-429 level was decreased in si-SNHG6 and anti-miR-429-cotransfected group.
SNHG6 silencing-mediated suppressive effects on the progression of Wilms' tumor are partly reversed by the addition of anti-miR-429.
The results of MTT assay and plate colony assay suggested that SNHG6 silencing suppressed the proliferation of Wilms' tumor cells, and this suppressive influence was largely counteracted by the interference of miR-429 (Fig. 4C–E). Si-SNHG6 transfection-induced apoptosis was partly attenuated by the introduction of anti-miR-429 in Wilms' tumor cells (Fig. 4F). According to the results of Western blot assay, SNHG6 interference decreased the expression of Bcl-2, whereas the expression of Bax was elevated in the SNHG6-silencing group (Fig. 4G, H). The addition of anti-miR-429 partly reversed si-SNHG6-mediated influence on the expression of Bcl-2 and Bax in Wilms' tumor cells (Fig. 4G, H). The results of flow cytometry and Western blot assay together demonstrated that SNHG6 silencing promoted cell apoptosis partly through elevating miR-429 level in Wilms' tumor cells.
Also, the authors explored if SNHG6 accelerated the glycolysis of Wilms' tumor cells through targeting miR-429. SNHG6 silencing suppressed LDH activity in Wilms' tumor cells, and LDH activity was largely recovered in si-SNHG6 and anti-miR-429-cotransfected group (Fig. 4I). Also, anti-miR-429 partly reversed the inhibitory influences of SNHG6 knockdown on the glucose utilization and lactate production in Wilms' tumor cells (Fig. 4J, K). SNHG6 silencing downregulated the level of ATP, and this inhibitory effect was partly alleviated by the addition of anti-miR-429 (Fig. 4L), suggesting that SNHG6 silencing restrained the production of ATP through upregulating miR-429 in Wilms' tumor cells. Accordingly, SNHG6 accelerated the progression of Wilms' tumor through targeting and downregulating miR-429.
FRS2 is a direct target of miR-429 in Wilms' tumor cells
As mentioned above, SNHG6 exerted its function at least partly through targeting miR-429 in Wilms' tumor cells. But how did miR-429 affect cell proliferation, apoptosis, and glycolysis? Bioinformatic analysis using Starbase database predicted that there existed complementary base pairs between FRS2 mRNA and miR-429 (Fig. 5A). The target relationship between miR-429 and FRS2 mRNA was then verified by dual-luciferase reporter assay. The luciferase activity was dramatically decreased in 3′-UTR-WT FRS2 and miR-429-cotransfected group compared with 3′-UTR-WT FRS2 and miR-NC group (Fig. 5B, C). However, miR-429 overexpression had almost no effect on the luciferase activity in 3′-UTR-MUT FRS2 group compared with 3′-UTR-MUT FRS2 and miR-NC group (Fig. 5B, C), indicated that FRS2 was a direct target of miR-429.

FRS2 is a direct target of miR-429 in Wilms' tumor cells.
The results of RNA pull-down assay revealed that FRS2 was enriched in Bio-miR-429-transfected group compared with that in Bio-miR-NC and Bio-miR-429-MUT group (Fig. 5D), suggesting the binding relationship between miR-429 and FRS2 in Wilms' tumor cells.
The mRNA and protein expression pattern of FRS2 in Wilms' tumor cells was tested by qRT-PCR and Western blot assay. As shown in Figure 5E and F, there was a notable upregulation on the mRNA and protein expression of FRS2 in Wilms' tumor tissues in comparison with that in matching nontumor tissues. Based on the expression of miR-429 and FRS2 mRNA, they generated the linear correlation curve as shown in Figure 5G. MiR-429 was negatively correlated with the mRNA expression of FRS2 (Fig. 5G). Also, FRS2 mRNA and protein were significantly elevated in Wilms' tumor cell lines compared with HEK293T cell line (Fig. 5H, I).
Subsequently, the authors tested the regulatory relationship among SNHG6, miR-429, and FRS2 in Wilms' tumor cells. Wilms' tumor cells were divided into four different transfected groups, including anti-miR-NC group, anti-miR-429 group, anti-miR-429 + si-NC group, and anti-miR-429 + si-SNHG6 group. FRS2 mRNA and protein expression was upregulated with the silencing of miR-429, and the abundance of FRS2 mRNA and protein was decreased again with the addition of si-SNHG6 (Fig. 5J–M). Overall, they concluded that FRS2 was a direct target of miR-429, and FRS2 was regulated by SNHG6/miR-429 axis in Wilms' tumor cells.
MiR-429 suppresses the progression of Wilms' tumor through targeting FRS2
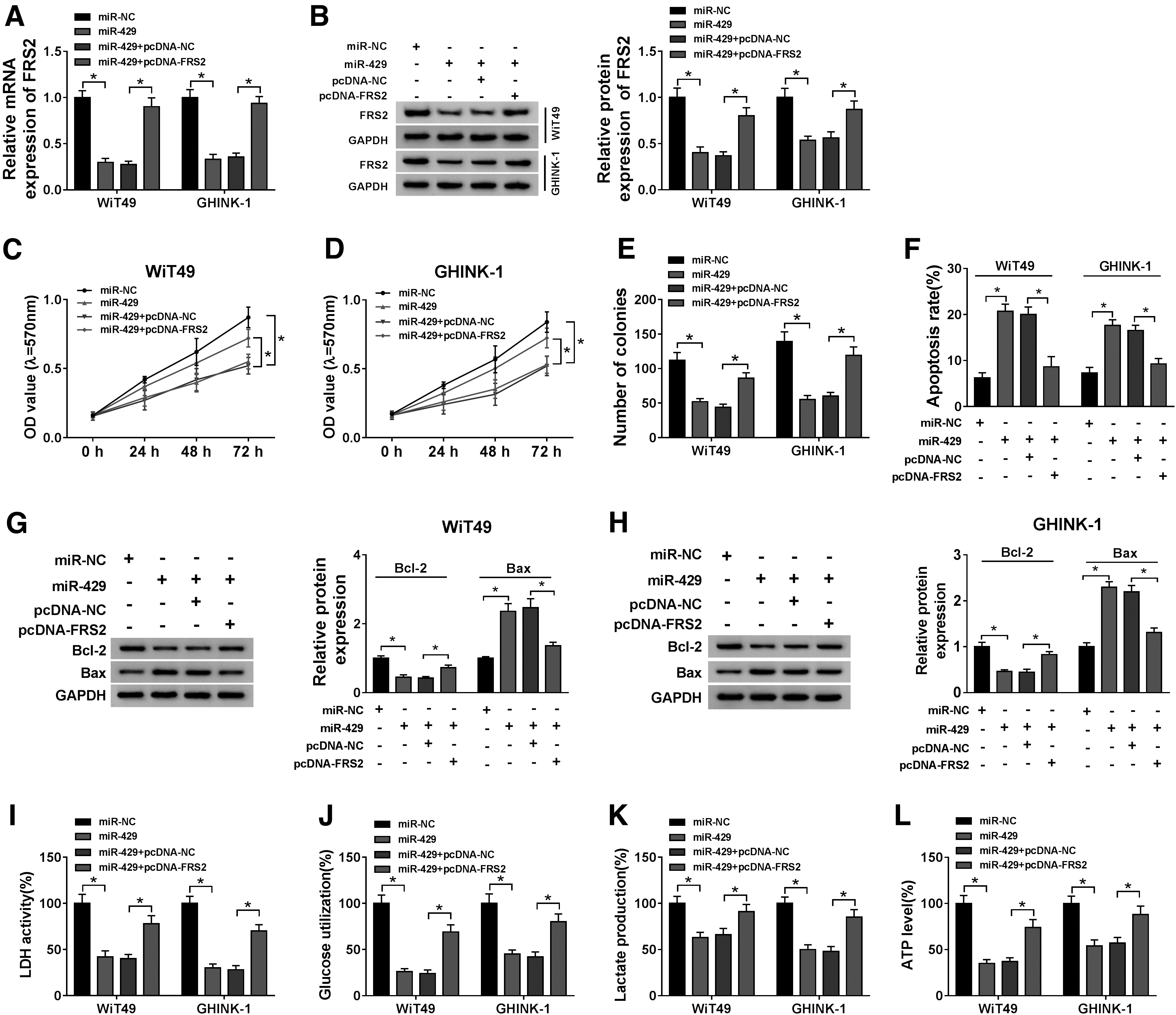
To investigate whether FRS2 was involved in miR-429-mediated influences on Wilms' tumor cells, they transfected miR-NC, miR-429, miR-429 + pcDNA-NC, or miR-429 + pcDNA-FRS2 into WiT49 and GHINK-1 cells to conduct rescue experiments. Before functional experiments, they measured the mRNA and protein expression of FRS2 in transfected Wilms' tumor cells. MiR-429 accumulation decreased the mRNA and protein abundance of FRS2, whereas FRS2 mRNA and protein levels were regained in miR-429 and pcDNA-FRS2-cotransfected group (Fig. 6A, B). On the grounds of the results of MTT assay and plate colony assay, they found that miR-429 restrained the proliferation, and the addition of FRS2 overexpression plasmid recovered the proliferation ability of Wilms' tumor cells (Fig. 6C–E).
MiR-429 suppresses the progression of Wilms' tumor through targeting FRS2.
The apoptosis was triggered with the overexpression of miR-429, and cell viability was recovered in miR-429 and pcDNA-FRS2 cotransfected group (Fig. 6F). The expression trend of proapoptotic protein Bax was consistent with the apoptosis rate of Wilms' tumor cells, whereas the expression of antiapoptotic protein Bcl-2 exhibited an inverse phenomenon to Bax (Fig. 6G, H), suggested that FRS2 overexpression counteracted the promoting effect of miR-429 on the apoptosis of Wilms' tumor cells.
The authors also detected the glycolysis ability of Wilms' tumor cells through measuring LDH activity, glucose utilization, and lactate and ATP production. MiR-429 accumulation suppressed the glycolysis of Wilms' tumor cells, whereas the glycolysis capacity was recovered with the introduction of FRS2 overexpression plasmid (Fig. 6I–L). The above data suggested that miR-429 exerted an antitumor role through targeting FRS2 in Wilms' tumor cells.
SNHG6 silencing restrains Wilms' tumor growth through miR-429/FRS2 axis in vivo
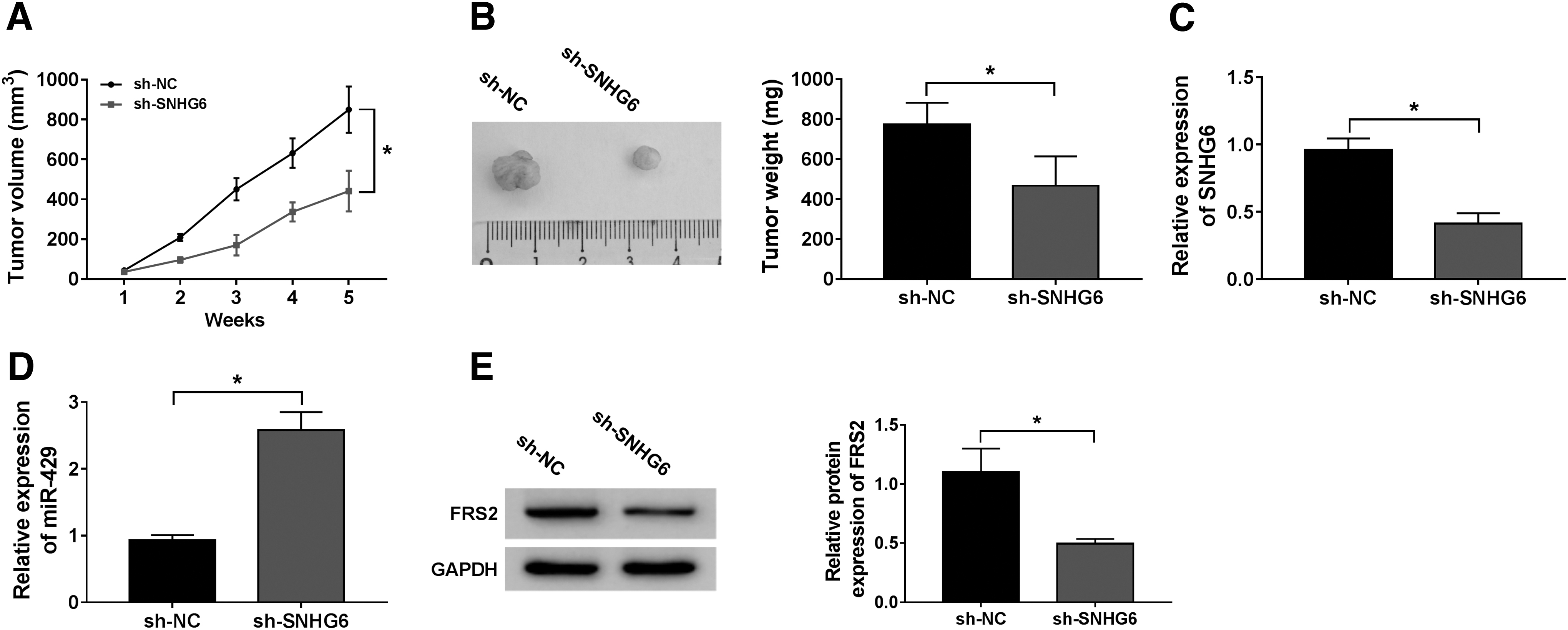
On the basis of the protumor role of SNHG6 in Wilms' tumor in vitro, they intended to explore if SNHG6 also exerted an oncogenic role in vivo. The authors established a murine xenograft model using the WiT49 cell line stably transfected with sh-SNHG6 or sh-NC to confirm the effect of SNHG6 in vivo. The tumor growth curve in sh-NC group and sh-SNHG6 group was generated through recording tumor volume every week using the formula of volume = (width 2 × length)/2. Tumors were resected from the mice after 5-week inoculation, and the weight was also recorded.
As indicated in Figure 7A and B, tumors from the sh-SNHG6 group were smaller compared with the sh-NC group. The representative tumor images in two groups are shown in Figure 7B. The expression of SNHG6, miR-429, and FRS2 was detected in excisional tumor tissues by qRT-PCR or Western blot, respectively. SNHG6 level was notably decreased in sh-SNHG6 group, suggesting that the interference was effective (Fig. 7C). The abundance of miR-429 was upregulated in tumor tissues from sh-SNHG6 group compared with that in sh-NC group (Fig. 7D). The enrichment of FRS2 protein was reduced in resected tumor tissues from sh-SNHG6 group compared with the sh-NC group (Fig. 7E). Therefore, they concluded that SNHG6 promoted Wilms' tumor growth at least partly through miR-429/FRS2 axis in vivo.
SNHG6 silencing restrains Wilms' tumor growth through miR-429/FRS2 axis in vivo.
Discussion
Wilms' tumor is a common malignancy that occurs in the urinary system of the children. LncRNA SNHG6 has been reported to play an oncogenic role in diverse cancers, including Wilms' tumor. 8,13,14 Nevertheless, the biological role of SNHG6 and the underlying mechanism in Wilms' tumor remain largely unknown. In this study, the authors identified SNHG6 as an oncogene and explored the crucial role of SNHG6/miR-429/FRS2 axis in Wilms' tumor progression.
Mounting evidences have pointed out the pivotal roles of lncRNAs in cellular physiological and pathological processes, including tumorigenesis. 15 –17 LncRNA SNHG6 level was abnormally increased and it acted as an oncogene in many kinds of cancers, including gastric cancer (GC), hepatocellular carcinoma (HCC), glioma, colorectal cancer (CRC), and Wilms' tumor. 8,18 –21 Yan et al. found that SNHG6 was overexpressed in GC, and SNHG6 promoted proliferation and motility of GC cells through regulating the levels of miR-101-3p and p27. 18 Cai et al. and Li et al. demonstrated that SNHG6 promoted glioma progression and CRC progression through regulating p21, respectively. 20,21
As for Wilms' tumor, Su et al. found that SNHG6 promoted cell proliferation and motility of Wilms' tumor cells through activating TAK1/JNK and Wnt/β-catenin signaling pathways through sponging miR-15a. 8 They found that the abundance of lncRNA SNHG6 was upregulated in Wilms' tumor. Functional experiments revealed that SNHG6 acted as an oncogene to promote the proliferation and glycolysis and impede the apoptosis of Wilms' tumor cells, and the protumor role of SNHG6 in Wilms' tumor was consistent with former work. 8
Accumulating articles have reported that lncRNAs functioned through acting as miRNA sponges to release downstream mRNAs, thus upregulating the levels of downstream mRNAs. To illustrate the associated working mechanisms by which SNHG6 promoted the progression of Wilms' tumor, SNHG6/miRNA interactions were predicted by miRcode online software. MiR-429 was a putative target of SNHG6, the relationship between SNHG6 and miR-429 was then validated by dual-luciferase reporter assay and RNA pull-down assay.
MiR-429 was reported to act as a tumor suppressor in multiple types of cancers. Deng et al. found that miR-429 was dramatically downregulated in osteosarcoma, and it played a suppressive role in tumor progression and metastasis through suppressing ZEB1 in osteosarcoma cells. 22 Xue and Tian reported that miR-429 restrained the metastasis of HCC cells through targeting and reducing the expression of RAB23. 23 Zhang et al. demonstrated that the abundance of miR-429 was declined, and it suppressed GC cell proliferation through downregulating FSCN1. 24
As for Wilms' tumor, Wang et al. found that miR-429 suppressed the proliferation of Wilms' tumor cells through targeting c-myc. 10 The enrichment of miR-429 was downregulated in Wilms' tumor. Subsequently, they wondered whether SNHG6 functioned through sponging miR-429, and rescue experiments were conducted. The results showed that SNHG6 promoted the progression of Wilms' tumor through targeting and downregulating miR-429.
MiRNAs are involved in the regulation of gene expression through restraining the translational process or declining the levels of target mRNAs in general. 25 –27 In terms of this article, FRS2 was predicted as a target of miR-429 by Starbase software. The relationship between miR-429 and FRS2 was confirmed by dual-luciferase reporter assay and RNA pull-down assay.
FRS2 is a component of FGFR signaling, and it has been reported to be an oncogene in many cancers. 28 –30 The enrichment of FRS2 was higher in Wilms' tumor compared with that in matching counterparts. Correlation analysis revealed that the abundance of FRS2 was inversely connected with the level of miR-429. To further investigate the role of FRS2 in miR-429-mediated effects of Wilms' tumor cells, rescue experiments were performed through transfecting miR-429 alone or together with pcDNA-FRS2 into Wilms' tumor cells. The results revealed that miR-429 suppressed the progression of Wilms' tumor through targeting FRS2.
Xenograft tumor model was built to explore the in vivo role of SNHG6 in the Wilms' tumor growth. Tumors generated from Wilms' tumor cells stably transfected with sh-SNHG6 grew more slowly compared with that in sh-NC group, suggested that SNHG6 accelerated the progression of Wilms' tumor in vivo.
Taken together, SNHG6 was identified as an oncogene in Wilms' tumor in this study. SNHG6 promoted Wilms' tumor progression through accelerating the proliferation and glycolysis and restraining the apoptosis of Wilms' tumor cells through miR-429/FRS2 axis. SNHG6/miR-429/FRS2 axis might be an underlying therapeutic target for Wilms' tumor.
Footnotes
Disclosure Statement
The authors declare that they have no financial conflicts of interest.
Funding Information
No funding was received for this article.