
Abstract
Background:
Circular RNAs (circRNAs) are regarded as important regulators in the tumorigenesis of multiple cancers. However, the characterization of circRNA exocyst complex component 6B (circEXOC6B) in ovarian cancer is barely known.
Materials and Methods:
Quantitative real-time polymerase chain reaction (qRT-PCR) was utilized to detect the enrichment of circEXOC6B, microRNA-376c-3p (miR-376c-3p), and forkhead box O3 (FOXO3). Cell proliferation was examined by Cell Counting Kit-8 (CCK8) assay and colony formation assay. Cell metastasis was measured by transwell assays. Western blot assay was conducted to examine the expression of proliferation and metastasis-related proteins and FOXO3. The chemoresistance of ovarian cancer cells was analyzed by CCK8 assay. Flow cytometry was used to detect cell apoptosis. The activities of caspase3 and caspase9 were analyzed through using colorimetric assay kits. The direct interaction between miR-376c-3p and circEXOC6B or FOXO3 was predicted by StarBase software and confirmed by dual-luciferase reporter assay and RNA immunoprecipitation (RIP) assay. Murine xenograft assay was conducted to verify the role of circEXOC6B on the paclitaxel (PTX) resistance of ovarian cancer cells in vivo.
Results:
The level of circEXOC6B was notably decreased in ovarian cancer tissues. Low level of circEXOC6B was associated with malignant pathological characteristics in ovarian cancer patients. CircEXOC6B suppressed the proliferation and motility and decreased the chemoresistance of ovarian cancer cells to PTX. CircEXOC6B functioned through directly targeting and downregulating miR-376c-3p. FOXO3 was a direct target of miR-376c-3p, and the abundance of FOXO3 was regulated by circEXOC6B/miR-376c-3p axis. CircEXOC6B accelerated the PTX sensitivity of ovarian cancer cells through acting as a decoy of miR-376c-3p to upregulate FOXO3 in vivo.
Conclusion:
CircEXOC6B suppressed the progression and PTX resistance of ovarian cancer cells through sequestering miR-376c-3p, thus enhancing FOXO3 level.
Introduction
Most ovarian cancer patients are diagnosed at late stage and exhibit dismal outcome with conventional therapeutic interventions. 1,2 Paclitaxel (PTX)-based chemotherapy was a conventional therapy method for patients with ovarian cancer. 3 However, tumor recurrence occurs with the development of chemoresistance. Therefore, understanding the potential mechanism behind the PTX resistance of ovarian cancer is needed to improve the outcome of chemotherapy.
Circular RNAs (circRNAs) are circular noncoding RNAs (ncRNAs) without 5′ and 3′ ends. CircRNAs are ideal targets for human diseases due to their stable structure. 4 Accruing studies have pointed out that the dysregulation of circRNAs was related to the progression of diverse cancers. 5 Besides, the aberrant expression of circRNAs was also related to the chemoresistance of multiple cancers. 6,7 Here, the authors intended to explore the role of circRNA exocyst complex component 6B (circEXOC6B) in the drug resistance of ovarian cancer cells.
MicroRNAs (miRNAs) have been reported to function as the downstream targets of circRNAs in cancers. For instance, Wang et al. proved that circ_0003159 blocked the development of gastric cancer via miR-223-3p/NDRG1 signaling. 8 In addition, miRNAs participated in the development of drug resistance in diverse cancers. For instance, miR-492 elevated the chemoresistance and promoted the metastasis of gastric cancer cells. 9 MiR-135a could facilitate the proliferation, motility, and chemoresistance of endometrial cancer cells. 10 MiR-376c has been reported to elevate the cisplatin resistance of ovarian cancer through ALK7. 11 In this study, they verified the combination between circEXOC6B and miR-376c-3p for the first time, and the effect of miR-376c-3p in the PTX resistance of ovarian cancer cells was explored.
Forkhead box O3 (FOXO3; FOXO3A) belongs to the forkhead box class O transcription factor family. 12,13 FOXO3 could have tumor promoting and tumor suppressor activities in human cancers. 14
Recent study clearly showed that high expression of FOXO3 predicted poor prognosis in ovarian cancer patients. 15 Nevertheless, there were also some articles which found that FOXO3 played a tumor suppressor role in ovarian cancer. FOXO3 depletion was an early event in the occurrence of high-grade serous ovarian cancer, 16 and the expression of FOXO3 was positively associated with the prognosis of ovarian cancer patients. 17,18 Salem et al. found that miR-590-3p facilitated the growth and aggressiveness of ovarian cancer cells via Cyclin G2 and FOXO3. 19 Here, the authors investigated the roles of FOXO3 in the malignant behaviors and chemoresistance of ovarian cancer cells.
According to the clinical data, they found that low expression of circEXOC6B might be a risk factor for ovarian cancer patients. Through conducting the bioinformatic prediction using online tool and verification via dual-luciferase reporter assay, they identified the target genes of circEXOC6B to illustrate its working network in the drug resistance of ovarian cancer cells.
Materials and Methods
Tissue collection
Ovarian cancer specimens (n = 60) and corresponding normal specimens (n = 60) were collected from Qingdao central hospital. The diagnoses of normal tissues and ovarian cancer specimens were performed by two pathologists. Every participant had submitted informed consent before the surgery. The procedures were authorized by the Medical Ethics Committee of Qingdao central hospital.
Quantitative real-time polymerase chain reaction
RNA samples were extracted using RNAiso kit (Takara, Dalian, China). Complementary DNA (cDNA) was obtained using HiScript II (Vazyme, Nanjing, China) and miRNA RT Kit (Haigene, Harbin, China). PCR was conducted with SYBR Premix Ex Taq™ GC master mix (Takara). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and U6 acted as the internal references with the quantification method of the 2−ΔΔCt. 20 The specific primers are presented in Table 1.
Primers for Quantitative Real-Time Polymerase Chain Reaction
Cell culture
A2780 and SKOV3 and normal human ovarian epithelial cell line (IOSE-80) were purchased from BeNa Culture Collection (Beijing, China). A2780 cells and SKOV3 cells were grown in Roswell Park Memorial Institute-1640 (RPMI-1640) medium (HyClone, South Logan, UT) added with 10% fetal bovine serum (FBS; Invitrogen, Gaithersburg, MD), 10% 100 U/mL penicillin, and 10% 100 μg/mL streptomycin. IOSE-80 cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco, Logan, Utah) supplemented with 10% FBS (Gibco), 10% 100 U/mL penicillin, and 10% 100 μg/mL streptomycin.
Cell transfection
The RNA oligoribonucleotides, including miR-376c-3p mimics (miR-376c-3p) and miRNA negative control (miR-NC), were purchased from GenePharma (Shanghai, China). CircEXOC6B overexpression (oe) plasmid (oe-circEXOC6B) and vector were also purchased from GenePharma. Lipofectamine 3000 reagent (Invitrogen) was used to transfect these RNA oligoribonucleotides and plasmids into ovarian cancer cells.
Cell Counting Kit-8 assay
For the detection of the proliferation ability, ovarian cancer cells were transfected with relevant plasmids or miRNAs followed by incubation with Cell Counting Kit-8 (CCK-8) (10 μL; Sigma, St. Louis, MO) for 4 h. For the detection of PTX resistance, ovarian cancer cells in 96-well plates were exposed to different doses of PTX. CCK-8 solution was added and the viability of ovarian cancer cells was determined through detecting the absorbance at 450 nm. IC50 value represented the half-maximum inhibitory concentration of PTX.
Colony formation assay
The capacity of ovarian cancer cells to form colonies was determined by colony formation assay. Two hundred single cells were plated into six-well plates and continued to culture for 2 weeks. The colonies were stained and counted.
Transwell assays
After precoating the inserts with Matrigel (transwell invasion assay), ovarian cancer cells suspended in 100 μL serum-free medium were seeded in the top chambers. The bottom chambers were filled with 600 μL medium added with 10% FBS. After culturing for 24 h, the invaded cells were dyed and imaged.
In transwell migration assay, ovarian cancer cells were directly seeded into the noncoated inserts, and the other steps were similar as above mentioned.
Western blot assay
Cell lysate was prepared using radioimmunoprecipitation assay (RIPA) kit (Beyotime, Jiangsu, China). Proteins were loaded onto sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and blotted on polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA). After blocking nonspecific sites, the membranes were probed with antiproliferating cell nuclear antigen (anti-PCNA; ab92552; Abcam, Cambridge, MA), anti-Cyclin D1 (ab16663; Abcam), anti-E-cadherin (ab40772; Abcam), anti-matrix metalloproteinase (anti-MMP9; ab219372; Abcam), anti-FOXO3 (ab109629; Abcam) or anti-GAPDH (ab181602; Abcam). The secondary antibody (ab205718; Abcam) was then incubated with the membranes. Enhanced chemiluminescence Western Blotting Substrate (Invitrogen) and X-ray films were used to visualize the protein bands.
Cell apoptosis analysis
Annexin V-fluorescein isothiocyanate (FITC)/propidine iodide (PI) Apoptosis Kit (Bender Med System, Vienna, Austria) was utilized in this study. Ovarian cancer cells were collected and rinsed with phosphate-buffered solution. After resuspended in binding buffer, these cells were incubated with Annexin V-FITC (62.5 ng/mL) and PI (5 μg/mL) for 10 min in a dark room and then analyzed with a flow cytometer.
Caspase activity detection
Colorimetric Assay Kits (R&D System, Minneapolis, MN) were utilized to measure the activities of caspase3 and caspase9. Cells were incubated with reaction buffer, including caspase3 or caspase9 specific colorimetric substrates for 1 h, and the optical density value was detected at 405 nm.
RNA extraction for nucleus and cytoplasm
PARIS™ Kit (Invitrogen) was used in this study. Ovarian cancer cells were disrupted using cell fractionation buffer. The nuclear section was separated from the cytoplasmic section by centrifugation.
Dual-luciferase reporter assay
The wild-type and mutant-type binding sites with miR-376c-3p in circEXOC6B and the 3′untranslated region (3′UTR) of FOXO3 were synthesized and inserted into psiCHECK-2 vector, generating circEXOC6B-WT, circEXOC6B-MUT, FOXO3 3′UTR-WT, and FOXO3 3′UTR-MUT. Ovarian cancer cells were transfected with these plasmids and miR-NC or miR-376c-3p. A Dual-Luciferase Reporter Assay Kit (Promega, Madison, WI) was used to analyze the luciferase activities in different groups.
RNA immunoprecipitation assay
Magna RNA immunoprecipitation (RIP) RNA-Binding Protein Immunoprecipitation Kit (Bersinbio, Guangzhou, China) was used in this study. Cell lysate was incubated with anti-Argonaute2 (anti-Ago2) (Millipore) or anti-Immunoglobulin G (anti-IgG) (Millipore) at 4°C overnight. Subsequently, magnetic beads were added to incubation for 1 h. Proteinase K was used to incubate with the samples for 1 h. Quantitative real-time polymerase chain reaction (qRT-PCR) was utilized to detect the abundance of circEXOC6B and miR-376c-3p.
Murine xenograft assay
BALB/c mice purchased from Orient Bio, Inc., (Seongnam, South Korea) were used in this study. The treatment method of nude mice had gotten permission of the Animal Research Ethics Committee of Qingdao central hospital. These nude mice were divided into four groups (n = 5). Two × 106 SKOV3 cells stably transfected with vector or oe-circEXOC6B were subcutaneously injected to the right side of mice back. After injection for 5 d, PTX (5 mg/kg) was intraperitoneally injected to the mice in PTX + vector group and PTX + oe-circEXOC6B group every 3 d. The tumor volume was measured every 5 d until 30 d after inoculation. The weight of tumors was measured using the formula of volume = π/6 × length × width × height.
Statistical analysis
Data are represented as mean ± standard deviation (SD). Student's t-test and one-way analysis of variance (ANOVA) followed by Tukey's test were used to analyze the differences as appropriate. Linear correlation was analyzed using Spearman's correlation coefficient. Survival curve of ovarian cancer patients was analyzed using log-rank test. p < 0.05 was considered to be significant.
Results
Low expression of circEXOC6B is a risk factor for ovarian cancer patients
CircEXOC6B was derived from the exon 3, exon 4, exon 5, and exon 6 of EXOC6B gene, and it was featured by continuous closed loop structure (Fig. 1A). CircEXOC6B was aberrantly downregulated in ovarian cancer tissues than that in matching nontumor tissues (Fig. 1B).
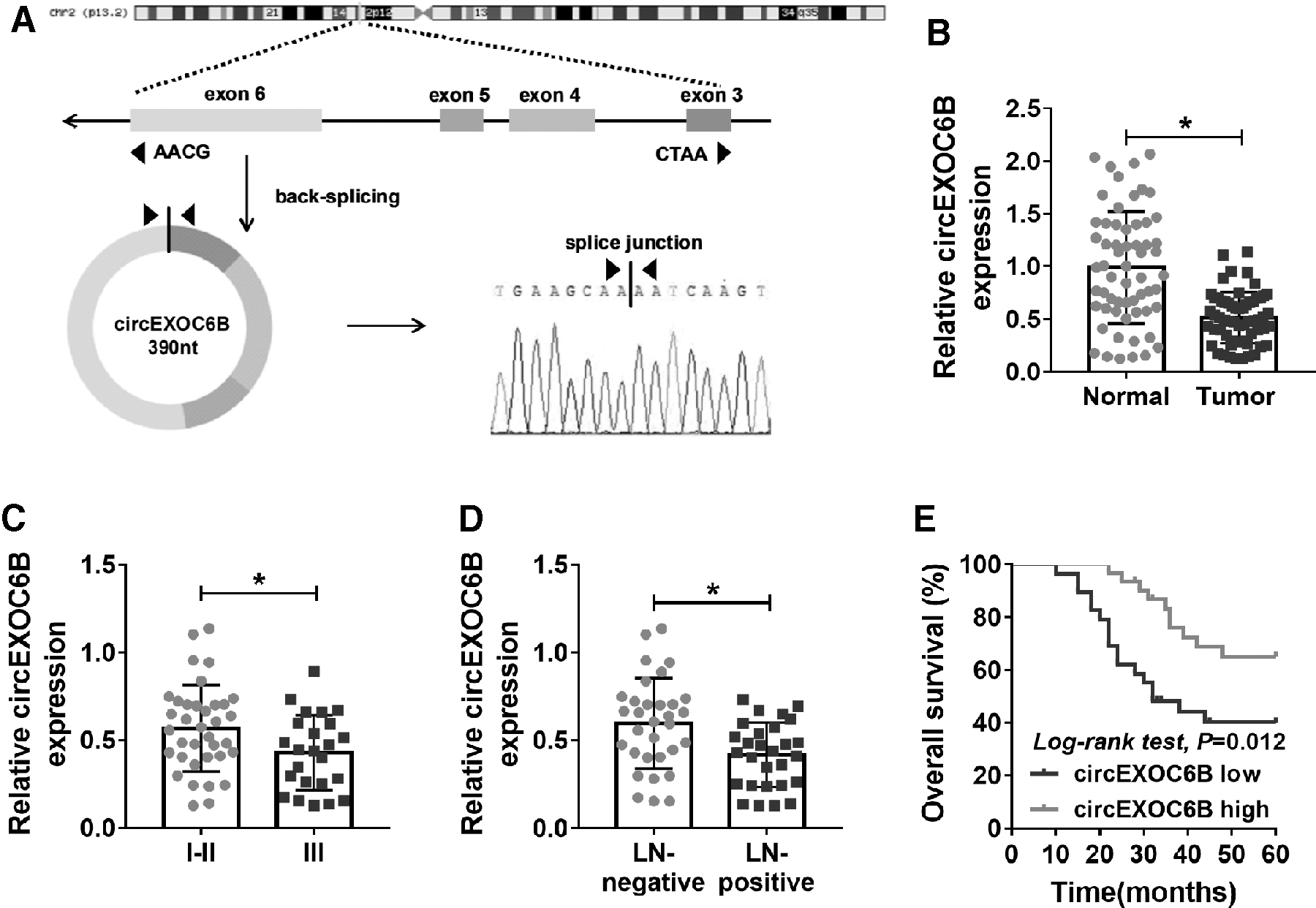
Low expression of circEXOC6B is a risk factor for ovarian cancer patients.
To assess the clinical significance of the abnormal downregulation of circEXOC6B in ovarian cancer patients, the authors analyzed the correlation between the expression of circEXOC6B and TNM staging or lymph node metastasis of ovarian tumors. As exhibited in Figure 1C and D, the expression of circEXOC6B was negatively related to the progression of ovarian cancer. Ovarian cancer patients were divided into two groups according to the median value of circEXOC6B level, and the high expression of circEXOC6B was associated with the long survival time of ovarian cancer patients (Fig. 1E). Overall, low level of circEXOC6B was a risk factor for ovarian cancer patients, and it might be a promising prognostic marker for ovarian cancer patients.
CircEXOC6B suppresses the proliferation and metastasis of ovarian cancer cells
The results of qRT-PCR revealed that circEXOC6B was downregulated in ovarian cancer cells compared with that in IOSE-80 cells (Fig. 2A). The overexpression efficiency of oe-circEXOC6B was high in ovarian cancer cells (Fig. 2B). The number of ovarian cancer cells in vector group outnumbered that in oe-circEXOC6B group after transfection for 48 and 72 h (Fig. 2C), suggested that circEXOC6B inhibited the proliferation of ovarian cancer cells. Meanwhile, the colony formation ability of ovarian cancer cells was significantly suppressed with the overexpression of circEXOC6B (Fig. 2D).
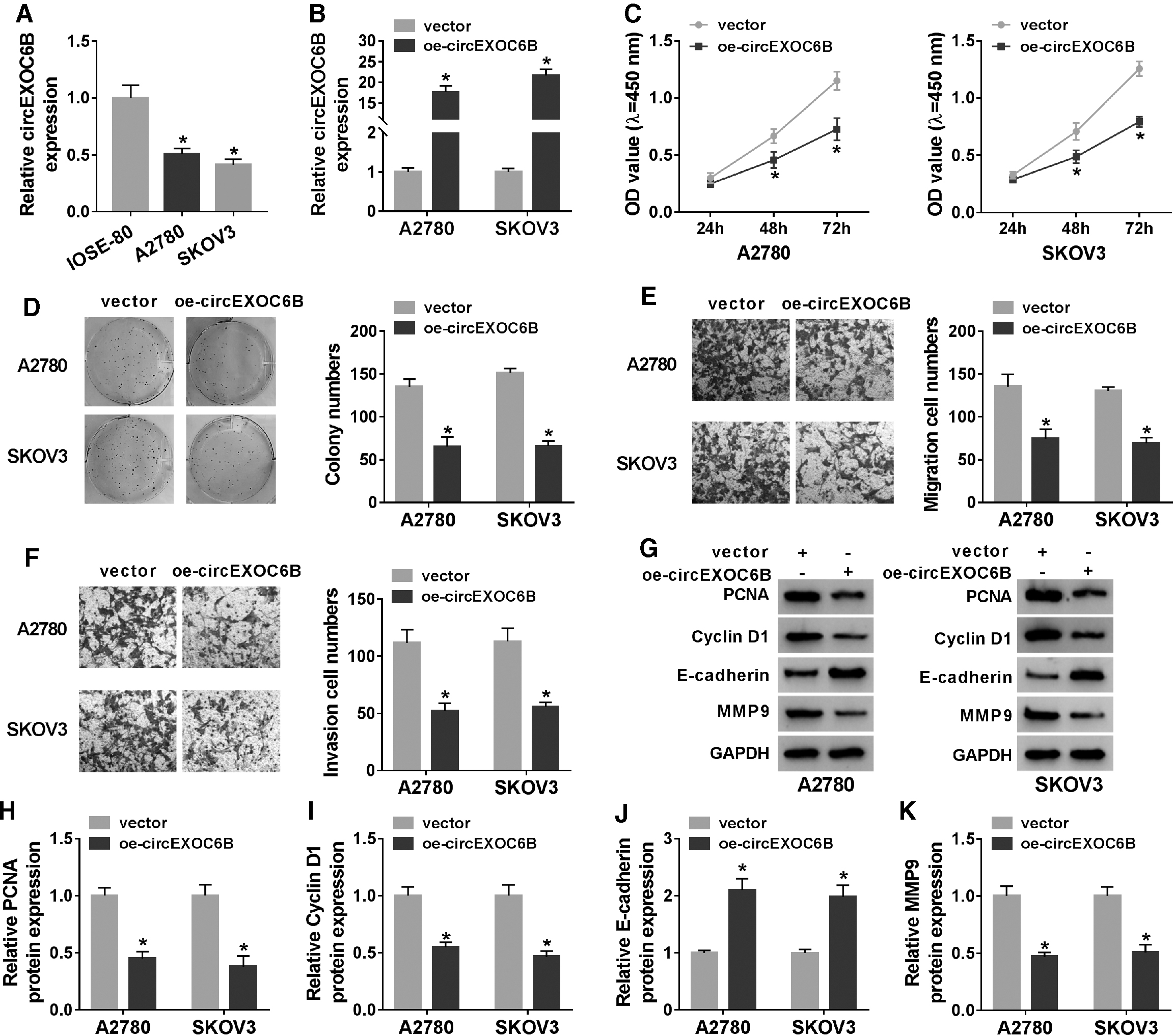
CircEXOC6B suppresses the proliferation and metastasis of ovarian cancer cells.
As mentioned in Figure 2E and F, the migration and invasion of ovarian cancer cells were notably inhibited in oe-circEXOC6B group than that in vector group. To further confirm the results, they conducted Western blot assay to detect the influence of circEXOC6B on the expression of proliferation and metastasis-associated proteins in ovarian cancer cells. As shown in Figure 2G–K, circEXOC6B overexpression downregulated the expression of PCNA, Cyclin D1, and MMP9, while upregulated the level of E-cadherin, demonstrated that circEXOC6B suppressed the proliferation and metastasis of ovarian cancer cells. In summary, circEXOC6B exerted an antitumor role to inhibit the proliferation and motility of ovarian cancer cells.
CircEXOC6B overexpression elevates the PTX sensitivity of ovarian cancer cells
To explore the effect of circEXOC6B on PTX resistance of ovarian cancer cells, they analyzed the IC50 value and apoptosis of ovarian cancer cells treated with oe-circEXOC6B and PTX using CCK8 assay and flow cytometry. As shown in Figure 3A and B, IC50 value of PTX was decreased with the accumulation of circEXOC6B, suggested that circEXOC6B overexpression elevated PTX sensitivity of ovarian cancer cells.
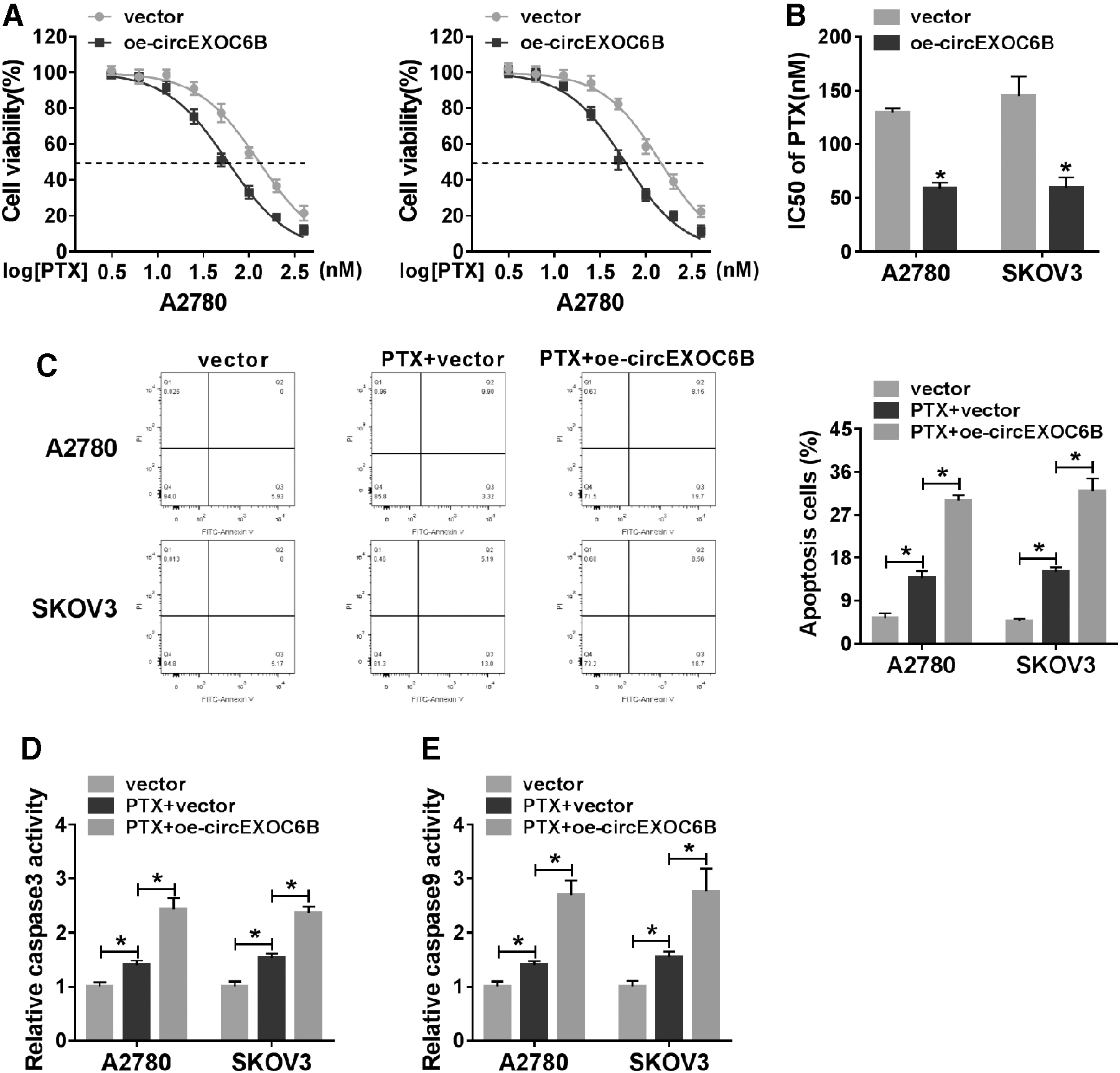
CircEXOC6B overexpression elevates the PTX sensitivity of ovarian cancer cells.
Moreover, the results of flow cytometry showed that PTX treatment promoted the apoptosis of ovarian cancer cells, and the addition of oe-circEXOC6B further accelerated the apoptosis of ovarian cancer cells than that in PTX + vector group (Fig. 3C). The authors also used colorimetric assay kits to confirm the influence of circEXOC6B overexpression on the apoptosis of ovarian cancer cells. As presented in Figure 3D and E, the activities of caspase 3 and caspase 9 revealed same trend to the apoptosis rate of ovarian cancer cells. Collectively, circEXOC6B ectopic expression sensitized ovarian cancer cells to PTX.
MiR-376c-3p is a target of circEXOC6B in ovarian cancer cells
CircEXOC6B mainly distributed in the cytoplasm of ovarian cancer cells (Fig. 4A). The putative binding sites between miR-376c-3p and circEXOC6B predicted by StarBase software are shown in Figure 4B. MiR-376c-3p and circEXOC6B-WT cotransfection reduced the luciferase activity in A2780 and SKOV3 cells, while the luciferase activity remained unchanged in circEXOC6B-MUT group with the addition of miR-NC or miR-376c-3p (Fig. 4C), suggested that miR-376c-3p bound to circEXOC6B via the predicted binding sites. CircEXOC6B and miR-376c-3p were both significantly upregulated in Ago2 group compared with that in IgG group (Fig. 4D), suggested that circEXOC6B could bind to miR-376c-3p-contained RNA-induced silencing complex (RISC).
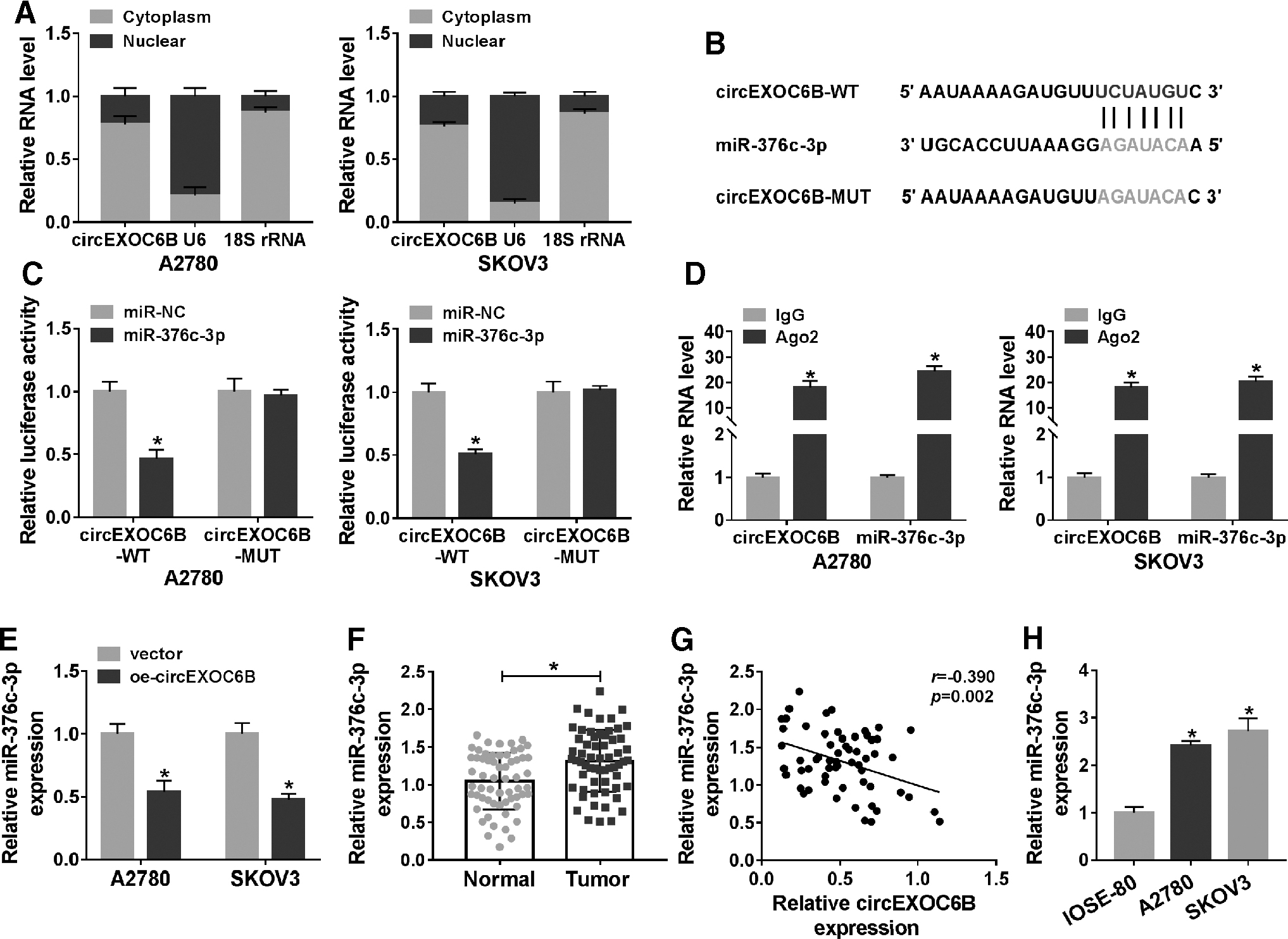
MiR-376c-3p is a target of circEXOC6B in ovarian cancer cells.
CircEXOC6B overexpression decreased the level of miR-376c-3p in ovarian cancer cells (Fig. 4E), suggesting the negative regulatory relationship between circEXOC6B and miR-376c-3p in ovarian cancer cells. MiR-376c-3p was found to be conspicuously upregulated in ovarian cancer tissues than that in corresponding normal tissues (Fig. 4F). There was an obvious negative correlation between the levels of miR-376c-3p and circEXOC6B (Fig. 4G). Moreover, the level of miR-376c-3p was prominently increased in ovarian cancer cells compared with that in IOSE-80 cells (Fig. 4H). Taken together, circEXOC6B could negatively regulate the expression of miR-376c-3p through direct interaction in ovarian cancer cells.
CircEXOC6B-mediated influence in ovarian cancer cells is overturned by the addition of miR-376c-3p
The functions of miR-376c-3p in cell proliferation, migration, and invasion and PTX resistance of ovarian cancer cells were assessed through overexpressing miR-376c-3p using miR-376c-3p mimic. As displayed in Supplementary Figure S1A, miR-376c-3p mimic transfection significantly upregulated miR-376c-3p level in ovarian cancer cells.
MiR-376c-3p overexpression promoted the proliferation of ovarian cancer cells via CCK8 assay and colony formation assay (Supplementary Fig. S1B, C). The migration and invasion abilities of ovarian cancer cells were both enhanced with the accumulation of miR-376c-3p (Supplementary Fig. S1D, E). The expression of PCNA, Cyclin D1, and MMP9 was elevated while E-cadherin expression was reduced in miR-376c-3p overexpression group compared with miR-NC group, suggesting that miR-376c-3p overexpression promoted the proliferation and metastasis of ovarian cancer cells (Supplementary Fig. S1F–J).
The IC50 value of PTX was enhanced in miR-376c-3p overexpression group than that in miR-NC group (Supplementary Fig. S1K). PTX-induced apoptosis in ovarian cancer cells was attenuated by the overexpression of miR-376c-3p (Supplementary Fig. S1L). Furthermore, the activities of caspase3 and caspase9 were elevated in PTX addition group, and these effects were largely alleviated by the overexpression of miR-376c-3p (Supplementary Fig. S1M, N). These findings together demonstrated that miR-376c-3p contributed to the progression and PTX resistance of ovarian cancer cells in vitro.
The authors cotransfected oe-circEXOC6B and miR-376c-3p into ovarian cancer cells to conduct rescue experiments. Oe-circEXOC6B inhibited the progression and PTX resistance of ovarian cancer cells as previously mentioned (Fig. 5A–M). The addition of miR-376c-3p diminished the inhibitory effect of circEXOC6B overexpression on the proliferation of ovarian cancer cells (Fig. 5A, B). CircEXOC6B-mediated inhibition on the migration and invasion of ovarian cancer cells was mitigated by the introduction of miR-376c-3p (Fig. 5C, D). Besides, the transfection of miR-376c-3p attenuated the influence of circEXOC6B on the expression of proliferation and metastasis-related proteins (Fig. 5E–I), suggesting that circEXOC6B hampered the proliferation and motility of ovarian cancer cells through targeting miR-376c-3p.

CircEXOC6B-mediated influence in ovarian cancer cells is overturned by the addition of miR-376c-3p. Ovarian cancer cells were transfected with vector, oe-circEXOC6B, oe-circEXOC6B + miR-NC, or oe-circEXOC6B + miR-376c-3p.
MiR-376c-3p elevated the PTX resistance of ovarian cancer cells that was downregulated by the overexpression of circEXOC6B (Fig. 5J). The apoptosis of ovarian cancer cells was analyzed through detecting the apoptosis rate and the activities of caspase3 and caspase9. As mentioned in Figure 5K–M, the introduction of miR-376c-3p recovered the viability of ovarian cancer cells treated with PTX that was suppressed by the accumulation of circEXOC6B. Overall, circEXOC6B blocked the progression and PTX resistance of ovarian cancer cells through directly interacting with miR-376c-3p.
FOXO3 is a downstream target of miR-376c-3p in ovarian cancer cells
According to the results of StarBase software, it was noticed that FOXO3 possessed complementary sites with miR-376c-3p on its 3′UTR (Fig. 6A). They constructed FOXO3 3′UTR-WT and FOXO3 3′UTR-MUT reporter plasmids to verify whether FOXO3 could directly bind to miR-376c-3p. As shown in Figure 6B, FOXO3 3′UTR-WT or FOXO3 3′UTR-MUT were cotransfected with miR-NC or miR-376c-3p into ovarian cancer cells, and luciferase activity was dramatically reduced in miR-376c-3p and FOXO3 3′UTR-WT group, proved that FOXO3 was a direct target of miR-376c-3p in ovarian cancer cells.
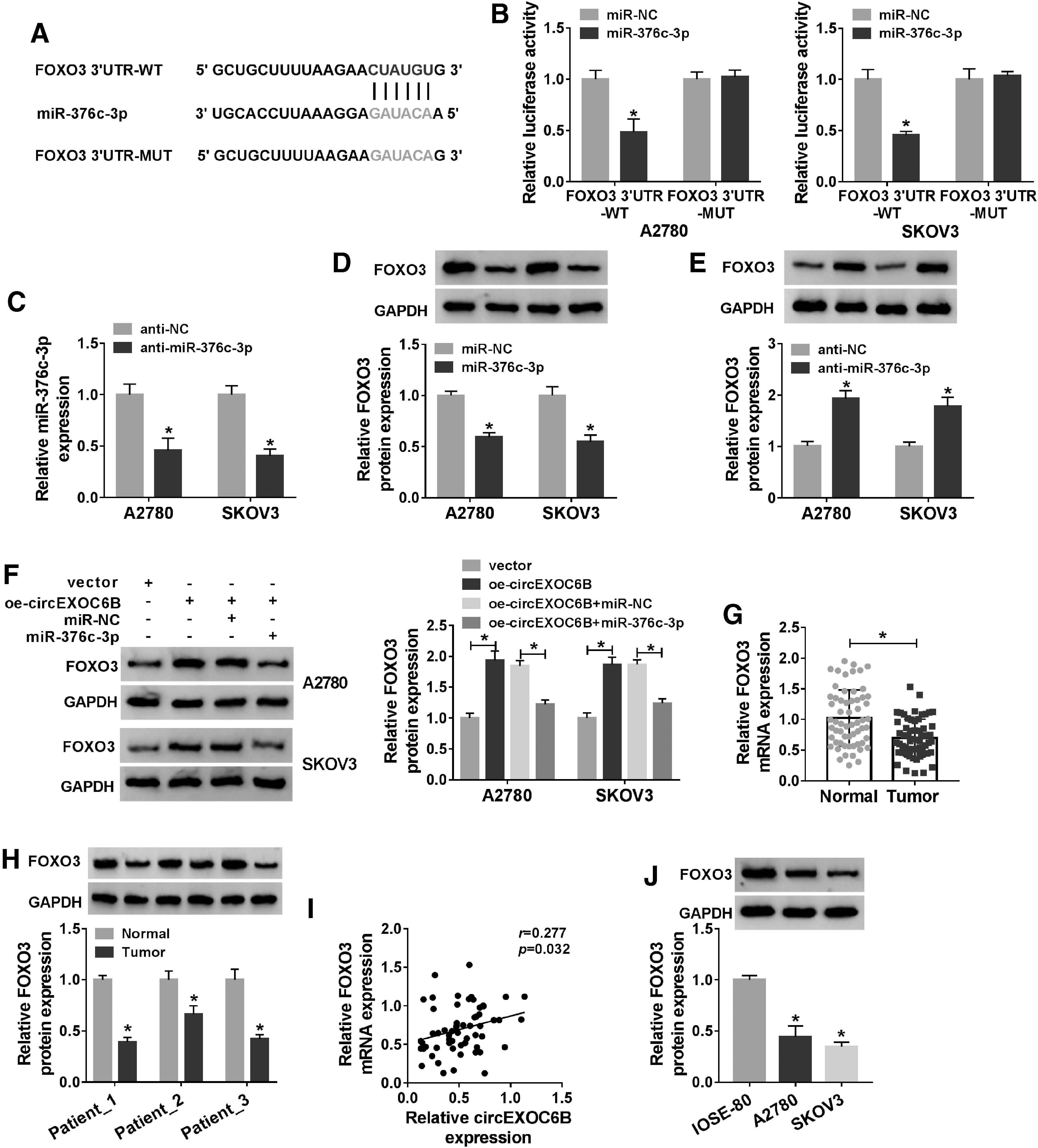
FOXO3 is a downstream target of miR-376c-3p in ovarian cancer cells.
The interference efficiency of anti-miR-376c-3p in ovarian cancer cells was high via qRT-PCR assay (Fig. 6C). MiR-376c-3p or anti-miR-376c-3p along with their negative controls was transfected into ovarian cancer cells to clarify the regulatory relationship between miR-376c-3p and FOXO3. As presented in Figure 6D, miR-376c-3p overexpression significantly decreased the level of FOXO3 in ovarian cancer cells. As shown in Figure 6E, miR-376c-3p silencing caused a significant elevation in FOXO3 protein expression in ovarian cancer cells. CircEXOC6B overexpression elevated the expression of FOXO3, while the addition of miR-376c-3p counteracted the above effect (Fig. 6F).
The expression pattern of FOXO3 in ovarian cancer was then explored. There was a notable downregulation in the mRNA and protein expression of FOXO3 in ovarian cancer tissues than that in adjacent normal tissues (Fig. 6G, H). The expression of FOXO3 was positively correlated with the level of circEXOC6B (Fig. 6I). Meanwhile, the expression of FOXO3 was markedly reduced in ovarian cancer cells than that in IOSE-80 cells (Fig. 6J). Collectively, miR-376c-3p could bind to FOXO3 and downregulate the level of FOXO3 in ovarian cancer cells.
CircEXOC6B overexpression enhances PTX sensitivity of ovarian cancer cells in vivo
To test whether circEXOC6B exerted a similar role in vivo, the authors conducted murine xenograft assay. They divided the nude mice into four groups: vector, oe-circEXOC6B, PTX + vector, and PTX + oe-circEXOC6B. CircEXOC6B overexpression or PTX treatment alone notably suppressed tumor growth through analyzing tumor growth curve (Fig. 7A) and tumor weight (Fig. 7B), and the cotreatment of circEXOC6B overexpression and PTX further inhibited the growth of ovarian tumors compared with PTX + vector group (Fig. 7A, B), suggesting that circEXOC6B accumulation sensitized ovarian cancer cells to PTX in vivo.

CircEXOC6B overexpression enhances PTX sensitivity of ovarian cancer cells in vivo.
Subsequently, the expression of circEXOC6B, miR-376c-3p, and FOXO3 was examined. As shown in Figure 7C and E, circEXOC6B and FOXO3 were prominently upregulated in oe-circEXOC6B and PTX + oe-circEXOC6B group in contrast to that in vector and PTX + vector group, respectively. Besides, the level of miR-376c-3p revealed an opposite trend to circEXOC6B and FOXO3 (Fig. 7D). Taken together, circEXOC6B elevated the PTX sensitivity of ovarian cancer cells in vivo.
Discussion
The dysregulation of circRNAs was involved in the pathogenesis of human cancers. 21,22 For instance, circ_100269 was reported to be downregulated in gastric cancer cells, and it hampered the growth of gastric cancer cells via miR-630. 23 CircPVT1 contributed to the growth and aggressiveness of nonsmall cell lung cancer (NSCLC) cells via miR-125b/E2F2. 24 Circ_0052112 accelerated the motility of breast cancer cells via miR-125a-5p. 25 Disclosing the potential mechanism between the abnormal expression of circRNAs and the progression of cancers is crucial to identify novel diagnostic and therapeutic targets.
The abundance of circEXOC6B was decreased in ovarian cancer, which was consistent with former article. 26 Besides, ovarian cancer patients with low expression of circEXOC6B possessed poor prognosis, implied that circEXOC6B might act as a tumor suppressor in ovarian cancer.
Overexpression experiments were conducted to disclose the effects of circEXOC6B. CircEXOC6B introduction inhibited the proliferation, migration, invasion, and PTX resistance of ovarian cancer cells. To explore the working mechanism by which circEXOC6B suppressed the development and drug resistance, the authors initially detected the distribution of circEXOC6B. CircEXOC6B mainly located in the cytoplasmic fraction of ovarian cancer cells.
CircRNAs could serve as crucial regulators in cancers through targeting miRNAs. 27 –29 For example, circUBA1 contributed to the progression of gastric cancer through sponging miR-375. 30 Geng et al. found that circ_0014130 suppressed the apoptosis of NSCLC cells through sponging miR-136-5p to upregulate BCL2. 31 MiR-376c-3p exerted a protumor role in many cancers. Lin et al. proved that Linc01278 hampered the progression of papillary thyroid carcinoma through targeting miR-376c-3p. 32 Wang et al. claimed that miR-376c-3p promoted the development of hepatocellular carcinoma. 33 Hung et al. found that miR-376c acted as an oncogene in gastric cancer. 34
Apart from these, the functional role of miR-376c in the cisplatin sensitivity of ovarian cancer cells has been explored before. Ye et al. demonstrated that miR-376c contributed to the cisplatin resistance of ovarian cancer cells via ALK7. 11 MiR-376c-3p was confirmed as a direct target of circEXOC6B. MiR-376c-3p overexpression promoted the proliferation, migration, invasion, and PTX resistance of ovarian cancer cells. Through conducting rescue experiments, the authors concluded that circEXOC6B suppressed the tumorigenesis and PTX resistance of ovarian cancer cells via targeting miR-376c-3p.
FOXO3 was verified as a downstream target of miR-376c-3p. Besides, circEXOC6B could upregulate the expression of FOXO3 through targeting miR-376c-3p. The role of FOXO3 in ovarian cancer was controversial. CDK6 was reported to protect epithelial ovarian cancer cells against platinum-mediated damage through phosphorylating and stabilizing FOXO3, 15 suggesting the oncogenic role of FOXO3 in ovarian cancer. Zhu et al. found that miR-148a suppressed the viability and motility in ovarian cancer cells through negatively regulating FOXO3 expression, 35 suggesting that FOXO3 exerted a protumor role in ovarian cancer.
FOXO3 was also reported to be a tumor suppressor in ovarian cancer. For instance, miR-590-3p was found to accelerate the progression of ovarian cancer through negatively modulating the expression of Cyclin G2 and FOXO3, 19 demonstrating the antitumor role of FOXO3 in ovarian cancer. In addition, the relationship between FOXO3 and chemoresistance of cancer cells has been explored in many cancers. For instance, Liu et al. proved that FOXO3 ectopic expression sensitized colorectal cancer cells to the exposure of 5-fluorouracil (5-FU) through Nrf2/TR1 pathway. 36 Lu et al. demonstrated that FOXO3 overexpression sensitized ovarian cancer cells to cisplatin. 37 Li et al. found that circ_0002483 blocked the development and Taxol resistance of NSCLC cells through upregulating GRB2, FOXO1, and FOXO3 via miR-182-5p. 38
Herein, they found that circEXOC6B exerted an antitumor role in ovarian cancer cells to enhance the PTX sensitivity through targeting miR-376c-3p to upregulate FOXO3 in vivo.
There are also some deficiencies in this study. The usage of two ovarian cancer cell lines (A2780 and SKOV3) did not fully recapitulate the common subtypes of human ovarian cancer (for example: high-grade epithelial ovarian cancer). The confirmation of the authors' conclusion in more comprehensive types of ovarian cancer cell lines is needed to verify if circEXOC6B/miR-376c-3p/FOXO3 functions in all subtypes of ovarian cancer.
In conclusion, their results revealed that circEXOC6B hampered the progression and elevated the PTX sensitivity of ovarian cancer through miR-376c-3p/FOXO3 axis, providing underlying targets to overcome the PTX chemoresistance of ovarian cancer cells.
Footnotes
Disclosure Statement
The authors declare that they have no financial conflicts of interest.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Original WB data revised
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.