
Abstract
Background:
Sevoflurane (Sev), a volatile anesthetic agent, is widely used in neurosurgery for anesthesia maintenance, accompanied with antitumor activity postanesthesia in multiple human cancers, including glioma. However, the molecular mechanism of Sev in glioma is largely unclear, including associated informative noncoding RNAs, such as long noncoding RNAs (lncRNA) and microRNAs (miRNAs).
Methods:
Expression of lncRNA KCNQ1 opposite strand/antisense transcript 1 (KCNQ1OT1), miRNA (miR)-146b-5p, and stanniocalcin-1 (STC1) was measured by real-time quantitative polymerase chain reaction and Western blotting. Cell proliferation, apoptosis, migration, and invasion in vitro were examined by 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay, fluorescence-activated cell sorting method, and transwell assays, respectively. Tumor growth in vivo was determined by xenograft models. The direct interaction between genes was confirmed by dual-luciferase reporter assay.
Results:
Sev enhanced apoptotic rate, but inhibited cell viability, migration, and invasion abilities of human glioma A172 and U251 cells in vitro, as well as tumor growth inhibition in vivo. The tumor-suppressive role of Sev in glioma was accompanied with downregulated KCNQ1OT1 and STC1, and upregulated miR-146b-5p. Overexpression of KCNQ1OT1 through transfection reversed, while KCNQ1OT1 silencing aggravated the antitumor role of Sev in A172 and U251 cells. Moreover, KCNQ1OT1-mediated tumor-promoting activity in A172 and U251 cells under Sev treatment was abrogated by miR-146b-5p restoration or STC1 deletion. Essentially, KCNQ1OT1 could positively regulate STC1 by acting as miR-146b-5p decoy.
Conclusion:
KCNQ1OT1 knockdown mediated the role of Sev in glioma cell proliferation, apoptosis, migration, and invasion both in vitro and in vivo through miR-146b-5p/STC1 pathway.
Introduction
Glioma is the leading malignant tumor in primary brain tumors, and its feature is highly aggressive, invasive, and destructive. 1 Surgery remains the paramount treatment for solid tumors including glioma. Nonetheless, metastatic recurrence always occurs after surgery, 2 thus serving as the major high hazard to mortality. In recent years, perioperative anesthetics have been proposed to affect tumor cell development, as well as cancer recurrence after surgery. 3,4 Sevoflurane (Sev) is a new type of volatile anesthetic agent for a wide use of inhalation anesthesia maintenance in neurosurgery. 5 It has been demonstrated that Sev exerts antiproliferation, migration, and invasion effects in multiple cancer cells, including glioma postanesthesia. 6,7 Moreover, the impact of Sev on long-term survival has been discussed in high-grade glioma 8 (World Health Organization [WHO] grades III and IV 9 ). However, the mechanism underlying Sev remains to be fully elucidated.
Emerging evidence suggests aberrant expression of long noncoding RNAs (lncRNAs) in response to Sev anesthesia. 10 LncRNAs are the transcripts longer than 200 nucleotides, and have been well documented in glioma. 11 KCNQ1 opposite strand/antisense transcript 1 (KCNQ1OT1) is a chromatin regulatory lncRNA, and plays essential role in malignant glioma. 12,13 However, the contribution of lncRNA to the role of Sev remains nearly blank in glioma. One famous type of molecular mechanism for lncRNAs is lncRNAs–microRNAs (miRNAs) interaction. MiRNAs are transcripts within 21–25 nucleotides. LncRNAs are serving competing endogenous RNAs (ceRNAs) to sponge miRNAs in glioma biology and pathology. 14 The expression profile of miRNAs has also been declared in glioma, 15 and miRNA (miR)-146b-5p is a tumor suppressor in glioma. 16 Very recently, this miRNA has been one molecular foundation for the application of Sev. 17
Stanniocalcin-1 (STC1) is a new biomarker of glioma progression partially being regulated by miRNAs. 18,19 In this study, we intended to explore the role and relationship among KCNQ1OT1, miR-146b-5p, and STC1 in glioma cells under Sev anesthesia.
Materials and Methods
Cells and cell culture
Human glioblastoma (WHO grade IV) cell line A172 (cat: 88062428) and astrocytoma (WHO grade I) cell line U251 (cat: 09063001) were obtained from European Collection of Authenticated Cell Cultures (Public Health England, United Kingdom), and cultivated in Dulbecco's modified Eagle's medium-high glucose (Sigma-Aldrich, Louis, MO) containing 10% fetal bovine serum (FBS; HyClone, Logan, UT). The cells were normally incubated in a humidified atmosphere containing 21% O2, 5% CO2, and 74% N2 in 37°C.
Sev treatment
Sev (Sevorane, Abbott Laboratories Queenborough, United Kingdom) was delivered from Sev vaporizer (Drage, Lubeck, Germany). The Sev machine was connected by the anesthesia vaporizer on the inlet port and a sealed glass chamber for cell treatment on the outlet port. The A172 and U251 cells were divided into four groups: control, 1.7% Sev, 3.4% Sev, and 5.1% Sev. All groups were incubated in a gas mixture containing 0–5.1% Sev, 21% O2, and 5% CO2, and balanced with N2 at 37°C. The gas monitor (Detex Ohmeda, Louisville, CO) monitored the gas fraction. Sev anesthesia was for 6 h, followed by normal cell culture for days.
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium assay
A MTS assay kit (Promega, Madison, WI) was employed to measure cell viability. In brief, cells in 96-well plate were further cultured for 0–3 d after Sev treatment, and 30 μL MTS solution was added to each well at indicated time points for another 4 h at 37°C. The optimal density value was read on a microplate reader BioTek Eon (BioTek Instruments, Winooski, VT) at a wavelength of 490 nm. Every group was set in at least three paralleled wells.
Fluorescence-activated cell sorting method
The Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis kit (BD Biosciences, Franklin Lakes, NJ) was used to detect apoptotic rate of A172 and U251 cells. In brief, cells in six-well plate were further cultured for 1 d after Sev treatment. The cells were collected, pelleted, and resuspended in the binding buffer, and then stained with Annexin V-FITC and PI reagent for 30 min in the dark. After adding ice-cold phosphate buffer solution, the cell suspension was analyzed on FACSCalibur flow cytometer (BD Biosciences).
Transwell assay
Cell migration and invasion abilities were measured using transwell chambers (Corning, Corning, NY). For cell migration assay, the system was established with the upper chamber filled with 100 μL of A172 or U251 cells in FBS-free DMEM, and the lower chamber loaded with 400 μL of DMEM supplemented with 20% FBS. The system was incubated in normal cell culture for 2 d, and then the lower surface was fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. Five randomly selected fields were captured under microscope at 100 × . The invasion assay was performed using chambers coated with Matrigel, and other operations were the same to migration assay.
Cell transfection
The miR-146b-5p mimics and the negative control (miR-NC) mimics, anti-miRNA against miR-146b-5p (anti-miR-146b-5p) and a scrambled anti-miR-NC, and siRNA target KCNQ1OT1 (si-KCNQ1OT1), si-STC1, and a scrambled si-NC were purchased from Ambion (Grand Island, NY). The pcDNA 3.1 vector was used to overexpress KCNQ1OT1. Cells in plate at 80% confluency were transfected using ViaFect™ Transfection Reagent (Promega) according to the manufacturer's protocol. The sequence of si-KCNQ1OT1 was 5′-GGGAAUCUGGUCUAAUGAATT-3′, 5′-UUCAUUAGACCAGAUUCCCTT-3′, and si-STC1 was 5′-GAGACAGCCUGAUGGAGAATT-3′ and 5′-UUCUCCAUCAGGCUGUCUCTT-3′, miR-146b-5p was 5′-UGAGAACUGAAUUCCAUAGGCU-3′, anti-miR-146b-5p was 5′-AGCCUAUGGAAUUCAGUUCUCA-3′, si-NC was 5′-UUCUCCGAACGUGUCACGUTT-3′ and 5′-ACGUGACACGUUCGGAGAATT-3′, miR-NC was 5′-UUUGUACUACACAAAAGUACUG-3′, anti-miR-NC was 5′-UCACAACCUCCUAGAAAGAGUAGA-3′.
Real-time quantitative polymerase chain reaction
The cellular total RNAs in tissues and cells were isolated by RNAiso Plus kit (Takara, Kusatsu, Shiga, Japan). The PrimeScript™ RT reagent kit with gDNA Eraser (Takara) and SYBR Premix Ex Taq (Takara) was utilized for the reverse transcription reaction and quantitative reaction, respectively. The primers including KCNQ1OT1 (5′-TGCAGAAGACAGGACACTGG-3′ and reverse 5′-CTTTGGTGGGAAAGGACAGA-3′), miR-146b-5p (5′-TGAGAACTGAATTCCATAGGCT-3′ and 5′-GTGCAGGGTCCGAGGT-3′), STC1 (5′-TTCACTCAAGCCAGGAGAGGGAAAG-3′ and 5′-AGGCATGCAAAAGCCCCGCAG-3′), U6 (5′-CTCGCTTCGGCAGCACA-3′ and 5′-AACGCTTCACGAATTTGCG-3′), and glyceraldehyde-phosphate dehydrogenase (GAPDH; 5′-AAGAAGGTGGTGAAGCAGGC-3′ and 5′-TCCACCACCCTGTTGCTGTA-3′) were synthesized by Riobio (Guangdong, China). The U6 and GAPDH were the endogenous controls. Melting curve analysis was used to confirm the specificity of amplification, and the threshold cycle (Ct) value was used to calculate RNA expression with the 2−ΔΔCt method. The real-time detection was conducted on StepOnePlus Real-Time PCR Systems (ABI, Foster City, CA). Every group cells were received the same procedures in at least three wells.
Dual-luciferase reporter assay
The putative binding sites of miR-146b-5p on KCNQ1OT1 and the 3′ untranslated region (3′UTR) of STC1 (STC1–3′UTR) were predicted according to starbase algorithm. Then, the fragments containing the putative binding sequences were mutated and cloned into psiCHECK-2 (Promega) as the mutant type (mut), compared with the corresponding wild type (wt). A172 and U251 cells in 24-well plate were cotransfected with wt/mut vectors and miR-146b-5p mimics or miR-NC mimics for 48 h. The dual-luciferase activities of A172 cells were measured on the Dual-luciferase Reporter Assay System (Promega). Every cotransfection group was repeated in three wells.
Western blotting
The cellular total proteins in tissues and cells were lysed in radioimmunoprecipitation buffer (Roche, Diagnostics, Mannheim, Germany). The isolated proteins were concentration determined using attached bicinchoninic acid assay kit, and 30 μg proteins were boiled in protein loading buffer before standard Western blotting procedures. The primary antibody including STC1 (20621-1-AP, 1:1000) and GAPDH (10494-1-AP, 1:25,000), and secondary antibody goat antirabbit (10285-1-AP, 1:10,000) were provided by Proteintech (Deansgate, Manchester, United Kingdom). The quantity of the proteins was analyzed in each group for three independent experiments on Image Lab™ software v5.2.1 (Bio-Rad, San Jose, CA).
In vivo experiment
The animal experiment was conducted in accordance with institutional guidelines for animal care and was approved by the Use and Care of Animals Committee of the First Affiliated Hospital of Xi'an Jiaotong University. A total of 10 male nude mice (BALB/c, 6–8 weeks old) were purchased from the HFK Bioscience Company, and randomly divided into two groups (n = 5): control and 5.1% Sev. The mice were subcutaneously injected with A172 cells (5 × 106) in 50% Matrigel into the right armpit. After cell transplantation for 7 d, the mice were anesthetized and administered 0 or 5.1% Sev for 6 h, as described above. Mice were excluded from the experiment if any signs of apnea or hypoxia were detected. After that, the mice were woken and reared in normal conditions for another 25 d. The length and width of tumors were monitored using a caliper every 5 d after Sev treatment, and the tumor growth curve was drawn according to the value of 0.5 × [the longest diameter] × [the widest diameter] 2 . The tumor weight was examined on electronic scale when the mice were euthanatized on the last day of animal experiment.
Statistical analysis
Data were expressed as mean (standard deviation). Student's t-test or one-way analysis of variance was performed to compare differences between two groups or among multiple groups using GraphPad Prism (GraphPad Software, La Jolla, CA). p-Values <0.05 were considered statistically significant, which is represented in the figures using asterisk.
Results
Functional analysis of Sev anesthesia on glioma cells in vitro
The functional analysis of Sev in glioma cells was performed. Fluorescence-activated cell sorting data showed that apoptotic rate of A172 and U251 cells after Sev anesthesia was promoted on 1 d (Fig. 1A); while cell viability on 3 d was significantly attenuated, as measured by MTS assay (Fig. 1B, C). Notably, the effect of Sev on cell viability and apoptotic rate was observed in a certain concentration-dependent manner, and 5.1% Sev exerted the optimal outcomes. Transwell assay evaluated cell migration and invasion ability, and the result was that migration cells and invasion cells were distinctively declined in A172 and U251 cells treated with 5.1% Sev (Fig. 2A, B). The above results indicated a suppressive role of Sev in malignant glioma cell development in vitro.
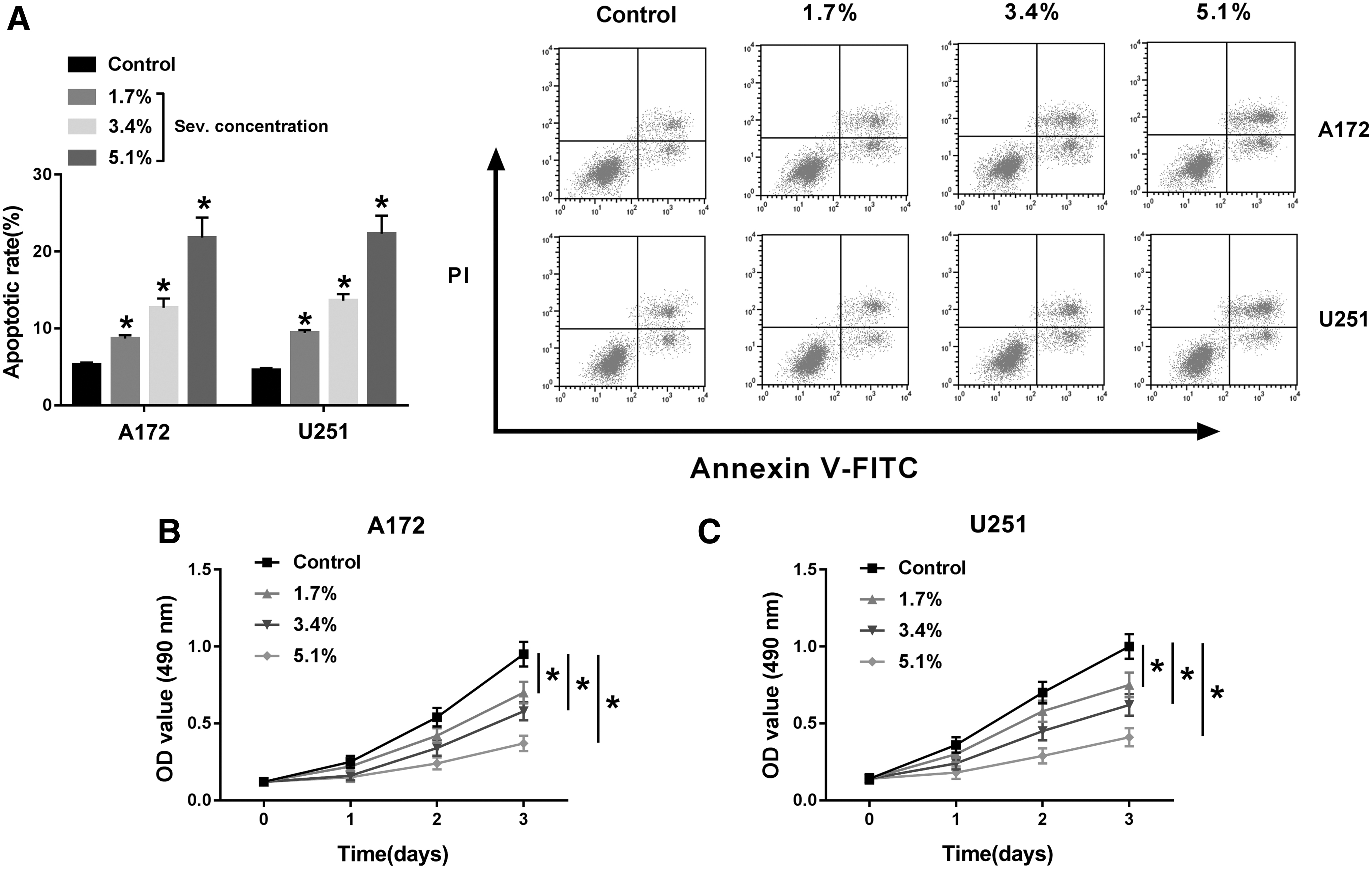
Functional analysis of Sev on glioma cell proliferation and apoptosis in vitro. A172 and U251 cells were treated with Sev (0–5.1%) for 6 h. After Sev exposure,

Functional analysis of Sev on glioma cell migration and invasion in vitro.
KCNQ1OT1 played oncogenic role in glioma cells postanesthesia of Sev in vitro
During Sev (5.1%) anesthesia for 6 h, relative KCNQ1OT1 expression was downregulated in A172 and U251 cells, paralleled with untreated cells (Fig. 3A). This finding suggested a potential impact of KCNQ1OT1 on the tumor-suppressive role of Sev in glioma postanesthesia. Thereby, KCNQ1OT1 was forcedly overexpressed or silenced in A172 and U251 cells through transfection before Sev (5.1%) anesthesia. And, real-time quantitative polymerase chain reaction (RT-qPCR) measured the transfection efficiency by analyzing KCNQ1OT1 expression. We found that KCNQ1OT1 level was extremely higher due to KCNQ1OT1 overexpression vector transfection, and was exceedingly lower in the presence of si-KCNQ1OT1 (Fig. 3B). Subsequently, gain-of-functional and loss-of-functional experiments were separately conducted. Ectopic expression of KCNQ1OT1 increased cell viability of A172 and U251 cells postanesthesia (Fig. 3C, D); contrarily, KCNQ1OT1 knockdown decreased that (Fig. 3H, I). Exogenous overexpression of KCNQ1OT1 diminished, whereas silencing of KCNQ1OT1 elevated the apoptotic rate of A172 and U251 cells administered with Sev (5.1%) (Fig. 3E, J). Transwell migration and invasion abilities of Sev-treated A172 and U251 cells were facilitated by KCNQ1OT1 overexpression, but inhibited by KCNQ1OT1 deletion (Fig. 3F–G, K, L). Collectively, these results demonstrated that KCNQ1OT1 negatively modulated the tumor-suppressive role of Sev in glioma cells in vitro.
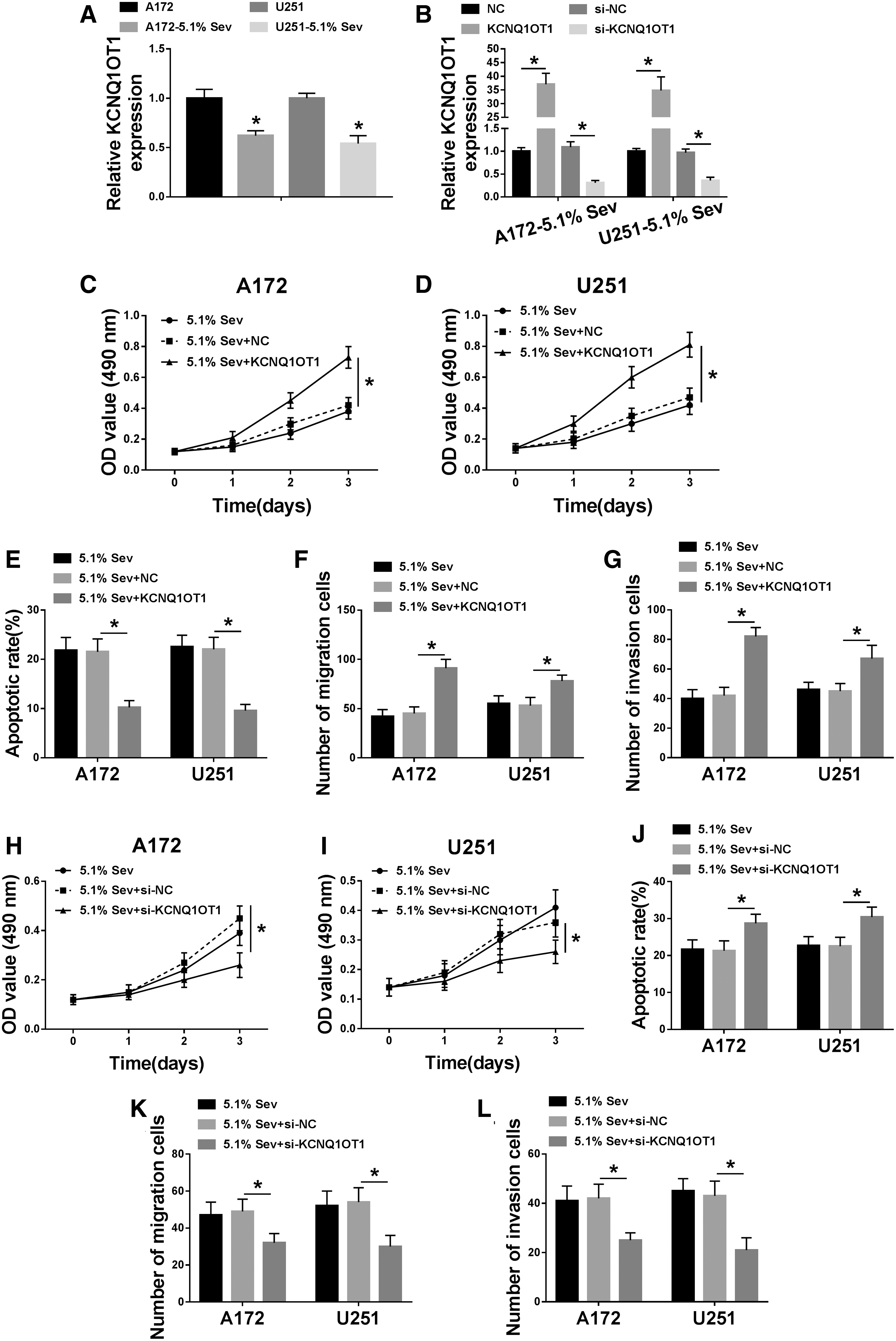
Role of long noncoding RNA KCNQ1 opposite strand/antisense transcript 1 (KCNQ1OT1) in Sev-treated glioma cells in vitro.
MiR-146b-5p silencing mediated the tumor-promoting effect of KCNQ1OT1 in glioma cells under Sev anesthesia through target binding
The target gene prediction software Starbase v.3.0 provided a putative binding sequence of miR-146b-5p on KCNQ1OT1, as presented in Figure 4A. Then, dual-luciferase reporter assay was utilized to validate this potential binding. Relative luciferase activity of KCNQ1OT1-wt was declined by miR-146b-5p mimics in A172 and U251 cells, compared with miR-NC mimics group (Fig. 4B, C). There was no significant difference of KCNQ1OT1-mut in A172 and U251 cells transfected with miR-146b-5p mimics or miR-NC mimics. Moreover, miR-146b-5p expression was inversely regulated by KCNQ1OT1 in A172 and U251 cells (Fig. 4D). In response to Sev anesthesia, miR-146b-5p was highly induced (Fig. 4E), and restoration of miR-146b-5p abolished the inhibitory effect of KCNQ1OT1 on miR-146b-5p expression in Sev-treated A172 and U251 cells (Fig. 4F). Functionally, introduction of miR-146b-5p mimics also counteracted the promoting effect of KCNQ1OT1 on cell viability of Sev-treated A172 and U251 cells (Fig. 5A, B). On the contrary, apoptotic rate was lower in KCNQ1OT1-overexpressed A172 and U251 cells under Sev anesthesia, which was further abrogated by miR-146b-5p upregulation (Fig. 5C). The enhancement of KCNQ1OT1 on transwell migration and invasion abilities was counteracted by miR-146b-5p upregulation as well (Fig. 5D, E). These data suggested that miR-146b-5p silencing might mediate the tumor-promoting role of KCNQ1OT1 in Sev-anesthetized glioma cells through target binding in vitro.

MiRNA (miR)-146b-5p was a direct target of KCNQ1OT1.

MiR-146b-5p mimics abrogated the effect of KCNQ1OT1 overexpression in glioma cells under Sev treatment. Sev-exposed A172 and U251 cells were pretransfected with KCNQ1OT1 alone or together with miR-146b-5p or miR-NC.
STC1 upregulation mediated the tumor-promoting activity of KCNQ1OT1 in glioma cells under Sev treatment through miR-146b-5p
The downstream functional gene of miR-146b-5p was followed to be identified. starbase3.0 provided a putative binding sequence of miR-146b-5p on STC1 (Fig. 6A). Dual-luciferase reporter assay depicted a decrease of luciferase activity of STC1–3′UTR-wt in miR-146b-5p mimics-transfected A172 and U251 cells (Fig. 6B, C). The overexpression of miR-146b-5p resulted in inhibited STC1 mRNA and protein expression in A172 and U251 cells, and miR-146b-5p knockdown led to raised STC1 expression (Fig. 6D–F). These findings prompted that STC1 was negatively regulated by miR-146b-5p through target binding in glioma cells. Furthermore, RT-qPCR and Western blotting analyzed downregulation of STC1 in Sev-anesthetized A172 and U251 cells (Fig. 6G, H), which was similar to KCNQ1OT1 (Fig. 3A). However, KCNQ1OT1 could induce STC1 upregulation in Sev-treated A172 and U251 cells, and this effect was further reversed by the presence of si-STC1 (Fig. 6I, J). Functionally, si-STC1 also counteracted the promoting effect of KCNQ1OT1 on cell viability, migration, and invasion abilities, and the inhibition on apoptotic rate of Sev-treated A172 and U251 cells (Fig. 7A–E). These data hinted that STC1 upregulation, similar to miR-146b-5p downregulation, could mediate the tumor-promoting role of KCNQ1OT1 in Sev-anesthetized glioma cells through miR-146b-5p in vitro.
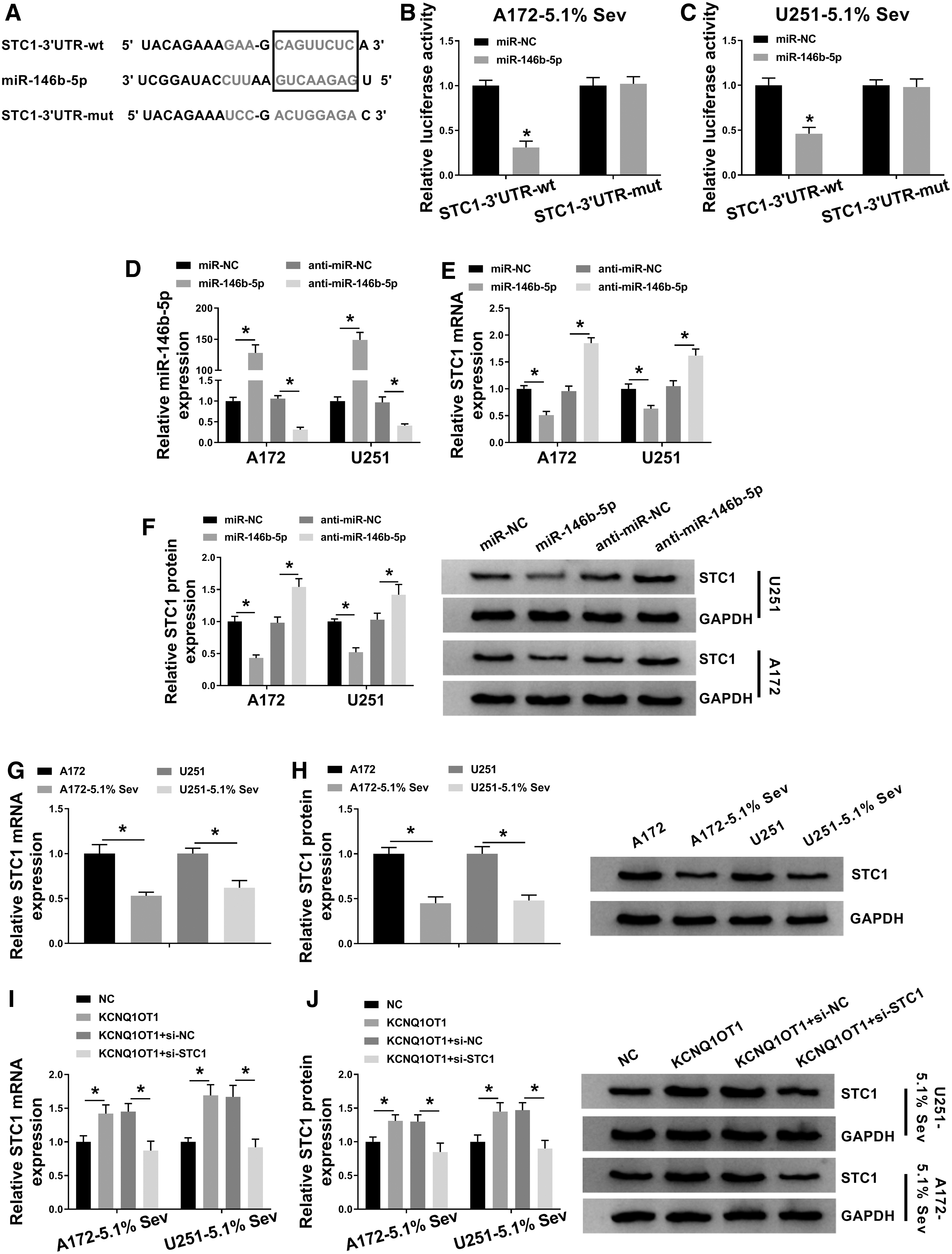
Stanniocalcin-1 (STC1) was a direct target of miR-146b-5p.
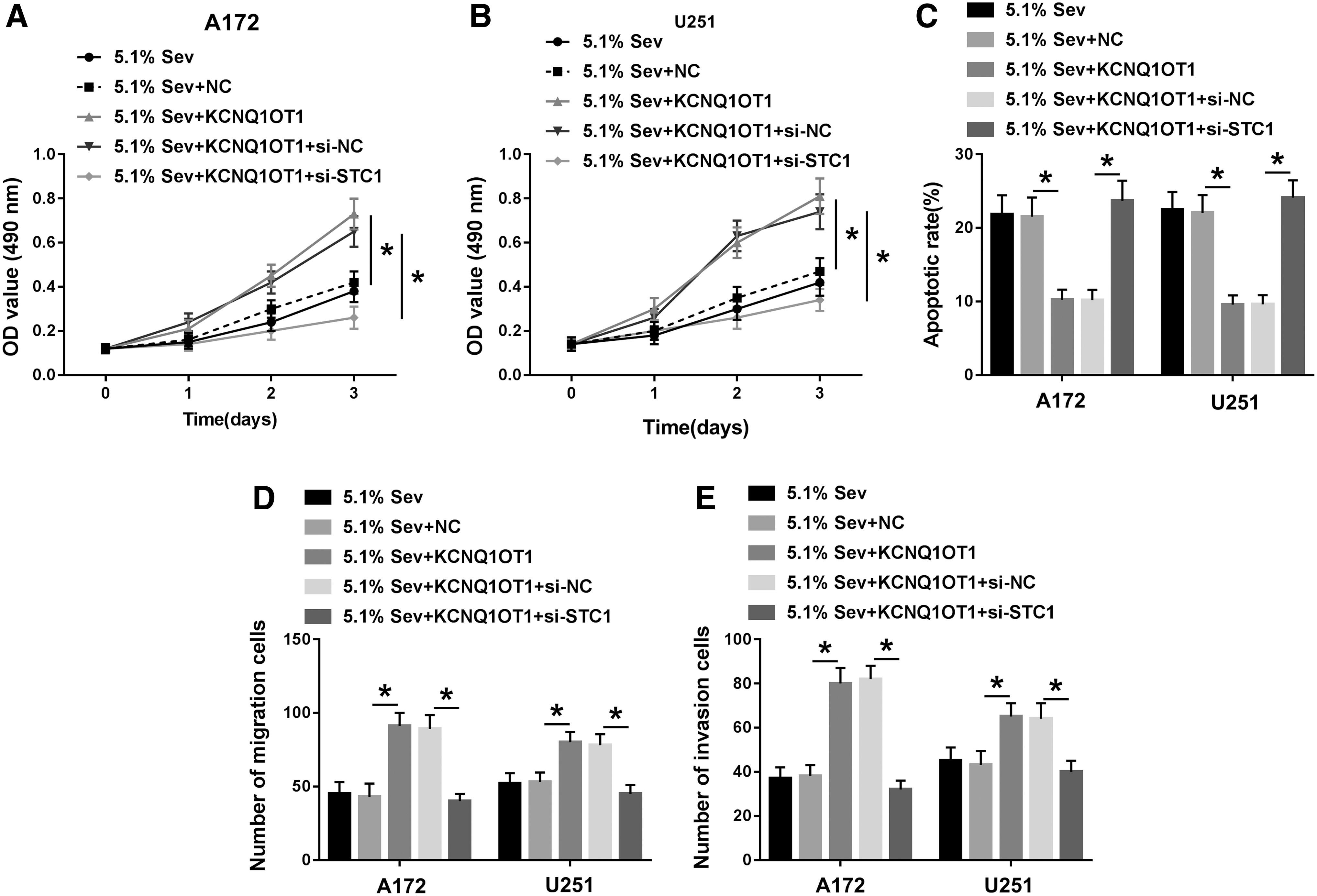
Si-STC1 abolished the effect of KCNQ1OT1 overexpression in glioma cells under Sev treatment. Sev-exposed A172 and U251 cells were pretransfected with KCNQ1OT1 alone or together with si-STC1 or si-NC.
Sev hindered tumor growth of glioma cells in vivo through downregulating KCNQ1OT1 and STC1, and upregulating miR-146b-5p
The role of Sev in tumorigenesis in vivo was also investigated. As shown in Figure 8A and B, the xenograft tumor growth of A172 cells in mice was hindered by Sev (5.1%) anesthesia, as evidenced by reduced tumor volume and tumor weight. Meanwhile, the KCNQ1OT1 and STC1 were less expressed (Fig. 8C, E, F), and miR-146b-5p was highly expressed (Fig. 8D) in the tumor tissues of mice. These results demonstrated that the tumor-suppressive role of Sev in tumor growth in vivo was partially regulated through KCNQ1OT1/miR-146b-5p/STC1 axis.

Sev hindered tumor growth of glioma cells in vivo. A172 cells were subcutaneously injected into nude mice, and 7 d later, the mice received 5.1% sevoflurane, 6 h.
Discussion
Perioperative anesthetic Sev exhibited direct effects on tumor cells and tumor stem cells, thus influencing tumor recurrence, metastasis, and long-term survival. 20,21 However, the effects of Sev inhalation in vitro studies seemed to be inconsistent across different cancer cell lines. 6 Although Sev probably played an antitumor role in most malignant tumors, such as glioma, hepatocellular carcinoma, and colorectal cancer, it was surprised to note that Lai et al. 22 claimed a promoting role of Sev in cell metastasis in glioblastoma both in vitro and in vivo. Except above research, Sev was widely documented to inhibit glioma proliferation, migration, and invasion. 7,17,23 By the way, still there were studies that concluded no effect of Sev on cell proliferation of glioma cells in vitro. 22,24 In consideration of the complicated role of Sev in glioma, we further confirmed the impact of Sev on cell proliferation, apoptosis, migration, and invasion in A172 and U251 cells, as well as the molecular mechanism focusing on informative RNAs.
Sev was associated with expression of aquaporin-4 and -9 that played pivotal role in cerebral edema, 25 a threat for patients who underwent gliomas resection. A cytoprotective property was conferred by Sev in glioma stem cells by inducing HIFs, which contributed to tumor initiation, metastasis, and recurrence. 20 Here, we observed that Sev induced inhibition on glioma cell viability, migration, and invasion abilities, and promotion on apoptotic rate, during which KCNQ1OT1 and STC1 were downregulated, and miR-146b-5p was upregulated. Knockdown of KCNQ1OT1 exerted tumor-suppressive function in Sev-treated glioma A172 and U251 cells, and similar antitumor role was discovered by Gong et al. 13 In that study, KCNQ1OT1 inhibition reduced cell proliferation, induced apoptosis, and weakened cell migration and invasion in vitro, but also impeded tumor growth, and prolonged survival time of mice carrying xenograft glioma tumors in vivo. Besides, miR-370/CCNE2 axis was directly regulated by KCNQ1OT1 during glioma malignant progression in U87 and U251 cells. In this study, we also identified a plausible KCNQ1OT1/miR-146b-5p/STC1 axis in Sev-treated A172 and U251 cells. However, we did not further validate the role of Sev/KCNQ1OT1/miR-146b-5p/STC1 pathway in the survival of glioma mice; while the both studies supported the notion that KCNQ1OT1 played a pivotal part in cancer development and cell fate determination. 26 By the way, several lncRNAs had been complicated in Sev-mediated cell toxicity in neurons and cardiomyocytes. This study was probably the first evidence about the interaction between Sev and lncRNA in glioma.
MiR-146b-5p was downregulated in glioma, and had been documented in glioma. 27,28 Moreover, it was observed to be highly expressed in recurrent glioblastoma tumors. 15,29 Overexpression of miR-146b-5p through transfection could restrain glioma cell proliferation, migration, and invasion by targeting MMP16, EGFR, and TRAF6, 16,30 –32 as well as stemness and radioresistance of glioma stem cells by targeting HuR/lincRNA-p21/β-catenin signaling pathway. 33 Furthermore, upregulation of miR-146b-5p by marrow stromal cell exosomes could reduce xenograft glioma tumor growth in rat. 34 Since miR-146b-5p had been implicated as a metastasis-inhibiting miRNA and immune-related miRNA in glioma, 29,30 the miRNA response elements of miR-146b-5p and other miRNAs were suggested to contribute to treat glioma by mediating tumor specific TRAIL delivery and oncolytic adenoviruses cytotoxicity. 28,35 Mechanically, miR-146b-5p was declared to be modulated by Sev in glioma, 17 as well as miR-124-3p and miR-637. 21,23 Here, we provided an upregulation of miR-146b-5p in glioma tumor tissues and cells under Sev anesthesia, thus endowing the tumor-suppressive activity in Sev-treated A172 and U251 cells. Furthermore, STC1 was disclosed as the novel target gene for miR-146b-5p.
STC1 was a secreted glycoprotein hormone in virtually all tissues. 36 STC1 served an oncogenic role in a number of different types of tumors, including glioma. 19 As a novel metastasis dissemination promoting factor, STC1 played promigration and invasion roles in glioma cells by regulating miRNAs in a hypoxia-dependent manner. 19,37 The stem-like traits of glioblastoma were augmented by STC1 in NOTCH1–SOX2 signaling pathway. 38 In clinic, the high STC1 level might predict a high-grade glioma and a worse outcome, thereby being a valuable biomarker in the diagnosis and prognosis of glioma. 18 Here, we discovered STC1 to be the downstream functional gene of Sev/KCNQ1OT1/miR-146b-5p axis in glioma. Expression of STC1 was downregulated in response to Sev both in vitro and in vivo, which was in consistence with findings in ovarian cancer from Zhang et al. 39 Moreover, upregulation of STC1 could lead to inhibitory effects on cell proliferation and metastasis in Sev-treated glioma cells, as well as ovarian cancer cells. 39
Conclusion
Collectively, we demonstrated a dysregulation of KCNQ1OT1, miR-146b-5p, and STC1 in response to Sev anesthesia in glioma, and indicated that a plausible mechanism for antitumor role of Sev entailed the depletion of KCNQ1OT1 and STC1, and the elevation of miR-146b-5p. This work suggested a KCNQ1OT1/miR-146b-5p/STC1 pathway underlying Sev anesthesia in glioma.
Footnotes
Disclosure Statement
The authors declare that they have no financial conflicts of interest.
Funding Information
This work was approved by the Clinical Research Award of the First Affiliated Hospital of Xi'an Jiaotong University (Grant No. XJTU1AF-CRF-2019-021), the National Natural Science Foundation of China (Grant No.81801388), and Natural Science Foundation of Shaanxi Province, China (2020JM388).