
Abstract
Background:
MicroRNA (miRNA) has been verified a significant factor to participate in the progression of colon cancer (CC). In this study, the authors investigated the mechanism and function of miR-548a-3p in CC.
Materials and Methods:
Bioinformatics analysis was used to analyze the mRNA expression profile and miRNA expression profile from GEO data series. The expression of miRNA and mRNA was analyzed by real-time quantification polymerase chain reaction in 43 pairs of CC clinical tissue samples and CC cells. The Western blot assay was used to detect the TPX2 protein. Then, SW480 and HCT116 cells were stably transfected with miR-548a-3p mimic, miR-548a-3p inhibitor, TPX2 overexpression, and TPX2 siRNA constructs to study the effects of miR-548a-3p and TPX2. Cellular functional experiments included cell counting kit-8 assay, BrdU incorporation assay, and wound healing assay. In addition, luciferase reporter assay was applied to detect the regulatory association between miR-548a-3p and TPX2.
Results:
TPX2 and miR-548a-3p were identified as the interested mRNA and miRNA by microarray analysis. In CC tissues and cell lines, miR-548a-3p with low expression and TPX2 with high expression were observed. What's more, exogenous overexpressed miR-548a-3p impaired the cell viability, cell proliferation, and cell migration, while TPX2 overexpression enhanced the malignancy phenotypes. However, the promotion effect of TPX2 on CC cells was impaired by miR-548a-3p.
Conclusion:
This study revealed that miR-548a-3p attenuated the development of CC by targeting TPX2.
Introduction
Colon cancer (CC) accounts for more than 70% of colorectal cancer (CRC), which has been the third most common and the second most deadly disease all over the world. 1,2 It is reported that CC has a high occurrence and mortality rate in North America, Australia, and Europe. 3 With the improvement of economic level, the occurrence of CC in East Asia has increased. 4 There has been reported that the median survival of the advanced CC patients with distant metastasis is less than 2 years. 5
At present, many molecular targeted therapies such as anti-VEG monoclonal antibody, VEGF tyrosine kinase inhibitor, and anti-EGFR monoclonal antibody have become a new treatment method for CC, thereby improving the life quality of CC patients. 6 –9 However, some patients do not respond to therapies that target these genes, thus it is necessary to find more diagnostic and treatment targets for CC.
During the last decade, it was demonstrated that a large number of small RNAs, including microRNAs (miRNAs), small interfering RNAs (siRNAs), and PIWI-related RNAs (piRNAs) at the length of 18–29 nucleotides, participated in the tumor progression. 10 –12 The endogenous miRNAs downregulated the target gene expression through translation repression or cleaving mRNAs. 13 It was reported that miRNAs controlled almost 30% of the expression of human proteins to promote or inhibit cellular processes, 14 and participated in diverse transcriptional regulation processes. 15 About a half of the miRNAs come from noncoding loci, while the rest are internally transcribed. 15,16 Tumor-associated miRNAs often act as promoters or suppressors in the development of cancer. 17
miR-548a-3p was a member of miR-548 family, which was proved to be upregulated in malignant pleural mesothelioma patients' sera. 18 Another reported that miR-548a-3p was significantly upregulated and acted as a diagnostic and prognostic biomarker for hepatocellular carcinoma. 19 However, whether miR-548a-3p was deregulated in CC tissues and how it might affect CC development remains unknown.
TPX2 is located on human chromosome 20q11.21 with 18 exons and encodes a microtubule-related protein. 20,21 At the early stage of mitosis, TPX2, a downstream effector of Ran-GTP, takes part in spindle formation. 22,23 Besides, TPX2 also strictly regulates all stages of the cell cycle, and it can be detected during G1-S transition and disappear at the completion of cytokinesis. 24 Therefore, the expression of TPX2 may provide an accurate evaluation for the proliferation of tumor cells. At present, many studies have proved that TPX2 is upregulated in tumor tissues such as lung cancer, pancreatic ductal adenocarcinomas, giant-cell tumor of bone, and CC. 25 –28 However, no researchers have identified whether TPX2 can be regulated by miRNAs in CC. By microarray analysis herein, the authors found that TPX2 was a key gene in CC participating in cell cycle biological process. How TPX2 affects CC cell phenotypes needs to be further explored.
They aimed to reveal the potential regulatory association between miR-548a-3p and TPX2 in CC. Together with the microarray analysis and previous studies, it was hypothesized that TPX2 might be the downstream target gene of miR-548a-3p and promote the development of CC. This study might provide a new method to diagnose or treat CC in the future.
Materials and Methods
Bioinformatics analysis
The mRNA microarray profile of GSE74604 downloaded from GEO DataSets (
The collection of clinical samples
The 43 paired clinical samples used in this study came from patients who were diagnosed with CC in West China Health Care Hospital of Sichuan University. Their CC tissues and corresponding adjacent normal tissues that were used in the experiments were approved by the Ethics Committee of West China Health Care Hospital of Sichuan University. The clinical characteristics of the patients are shown in Table 1.
The Clinical Characteristics of 43 Cases of Colon Cancer Patients
RNA extraction and real-time quantification polymerase chain reaction
Total RNA was obtained using the miRcute miRNA Isolation Kit (Cat. No.: DP501; Tiangen Biochemical, China). After using gel electrophoresis to check the purity of the RNA, the authors performed the RNA reverse transcription. They used the miRcute Plus miRNA First-Strand cDNA Kit (Cat. No.: KR211; Tiangen Biochemical) for miR-548a-3p reverse transcription, and the PrimeScript RT Reagent Kit (Cat. No.: RR037A; Takara, Japan) for TPX2 reverse transcription. Then, they ran the quantitative polymerase chain reaction (PCR) in ABI 7500 using the TB Green Premix Ex Taq II (Cat. No.: RR820A; Takara). GAPDH was used as the reference gene for TPX2 mRNA and U6 as the reference gene for miR-548a-3p. All the primers (sequences presented in Table 2) were purchased from GeneCopoeia (Rockville, MD). The Ct levels were obtained and the relative expression of miR-548a-3p and TPX2 mRNA was calculated using the 2−ΔΔCt method.
The Sequences of the Primer Used in This Study
Cell culture
The human CC cell lines HCT116, SW480, KM12, Caco-2, and SW620 and normal colonic epithelial cell HCoEpiC were purchased from BNCC (BeNa Culture Collection, Beijing, China). HCT116 and KM12 cells were cultured in RPMI 1640 supplemented with 10% FBS in humidified incubator with 5% CO2 at 37°C. SW480, Caco-2, SW620, and HCoEpiC cells were cultured in DMEM with other condition same with HCT116.
Cell transfection
miR-548a-3p mimic (Cat. No.: B01001), miR-548a-3p inhibitor (Cat. No.: B03001), TPX2 siRNA (si-TPX2, Cat. No.: A01004), overexpression TPX2 (OE-TPX2, Cat. No.: LV11), and negative control (NC, Cat. No.: not applicable) plasmids were designed from GenePharma (Shanghai, China). HCT116 and SW480 cell lines were added into six-well plates for transfection at the cell density of 2 × 104/mL. After incubation overnight, 60 nM miR-548-3p mimic, miR-548a-3p inhibitor, si-TPX2, OE-TPX2, and NC constructs were transfected using Lipofectamine 2000 reagent (Cat. No.: 11668027; ThermoFisher) on the basis of the protocol. After 48 h transfection, the transfection efficiency was measured by real-time quantification polymerase chain reaction (qRT-PCR).
BrdU assay
Cell proliferation was evaluated using BrdU incorporation assay. The transfected HCT116 and SW480 (2 × 104/mL) cells were seeded into a 96-well plate for a day. The cells were subsequently incubated with 10 μL BrdU (5-bromo-2′-deoxyuridine) and Thymidine analog (Cat. No.: ab142567; Abcam, UK) for 4 h. The optical density was measured by a scanning multiwell spectrophotometer at 450 nm.
Cell Counting Kit-8 assay
miR-548a-3p mimic, miR-548a-3p inhibitor, si-TPX2, OE-TPX2, or NC were stably transfected into the CC cell lines SW480 and HCT116. 3500/well cells were seeded into 96-well plate. Cell viability was measured using Cell Counting Kit-8 (CCK-8) (HY-K0301; MedChemExpress, China) at 0, 24, 48, and 72 h. Ten microliters of CCK-8 solution was added into every well. Next, the authors placed the plate at 37°C in the incubator for 4 h. Finally, the optical density was read at 450 nm under a microplate reader.
Wound healing assay
The transfected cells were seeded into a 12-well plate with the density of 4 × 105 per well and cultured until the cells covered the bottom of the well. The fused monolayer cells were scratched by a yellow micropipette tip in the center of the well. Washing the exfoliated cells by PBS for twice, the cells continued to grow in cell media with no serum for 24 h. The authors captured images at 0 and 24 h to observe the cell migration distance. The migration rate was calculated as (gap distance at 0 h−gap distance at 24 h)/gap distance at 0 h to represent the migration capability of cells in different groups.
Luciferase reporter assay
The authors constructed the vectors of wild strains and mutants of TPX2 (TPX2-3′UTR WT and TPX2-3′UTR Mut, respectively) that were bought from GeneCopoeia. Next, they transfected the TPX2-3′UTR WT and/or Mut vectors and miR-548a-3p mimic and/or NC into the target cells HCT116 and SW480 by Lipofectamine 2000 reagent (11668027; ThermoFisher). Cells were collected and lysed with lysis buffer after 48 h. The GeneCopoeia dual-luciferase reporter assay system (LF031) was applied for luciferase activity measurement.
Western blot assay
The total protein was extracted from HCT116 and SW480 cells using RIPA buffer with 1% protease inhibitor cocktail. After measuring the protein concentration, the protein was separated by SDS-PAGE and transferred onto PVDF membranes. Then, the membranes were incubated with anti-TPX2 (Cat. No.: ab252945; Abcam) and anti-GAPDH (Cat. No.: ab181602; Abcam) primaries overnight. The hybrids were then blocked in skim milk. Next, the membranes continued to incubate with the secondary antibody for 3 h. Finally, the protein blots were visualized using ECL Kit (Thermo Fisher Scientific) under a Bio-Rad gel imager.
Statistical analysis
The authors analyzed the results by SPSS 23.0 (SPSS, Inc., Chicago, IL). Data of every assay, exhibited as mean ± standard deviation, come from three independent experiments. Student's two-tailed t-test and one-way analysis of variance with Dunnett's post hoc were, respectively, used to evaluate the significance of the differences between two groups and multiple groups. p ≤ 0.05 was set as the threshold for statistical significance.
Results
TPX2 and miR-548a-3p were screened out as the interested mRNA and miRNA
With logFC more than 2, a total of 78 upregulated DEGs were selected from GSE74604 data series (Supplementary Table S1) and uploaded to String for biological process analysis (Fig. 1A). The regulation of cell cycle process involving 27 DEGs was the key biological process by String analysis (marked in red, Fig. 1B). The G2/M transition involving nine DEGs was considered as the key biological process by Metascape analysis (Fig. 1C). Because G2/M transition belongs to the regulation of cell cycle, they overlapped the DEGs from String results and Metascape results and confirmed eight DEGs, namely AUBKA, CCNA2, CCNB1, CCNB2, CDK1, CENPF, HMMR, and TPX2 (Fig. 1D). By uploading the eight DEGs to UALCAN, they found that AUBKA, CCNA2, and TPX2 were the significantly upregulated DEGs in colon adenocarcinoma (Fig. 1E). The authors then used TargetScan Human 7.2 to predict the targets of the three genes, and intersected them with the 729 downregulated miRNAs in CC tissues from GSE136020 with logFC <0 (Supplementary Table S2). Finally, they identified TPX2 and miR-548-3p as the interested mRNA and miRNA, respectively, for further investigation because miR-548-3p was both a downregulated miRNA in GSE136020 and a predicted target of TPX2 from TargetScan Human 7.2 (left panel of Fig. 1F).

The TPX2 and miR-548a-3p were the respective key genes and miRNA to be further investigated in CC.
miR-548a-3p was downregulated in CC and inhibited the malignancy phenotypes of CC cells
To explore the effect of miR-548a-3p on CC cells, they first measured the expression of miR-548a-3p in healthy and tumorous colon tissues, finding that the expression of miR-548a-3p was downregulated by 49.8% in CC tissues (Fig. 2A). Twenty-three of the 43 paired tissues were assigned to the training set, and the other 20 were assigned to the validation set. Data analysis of both training set and validation set showed that miR-548a-3p was significantly downregulated in CC tissues (Supplementary Fig. 1A, C). At the same time, they also used five CC cell lines (HCT116, SW480, KM12, Caco-2, and SW620) and normal colonic epithelial cell (HCoEpiC) to measure miR-548a-3p expression by qRT-PCR. It was shown that miR-548a-3p was significantly downregulated in five CC cell lines compared with the normal colonic epithelial cell line HCoEpiC (Fig. 2B).
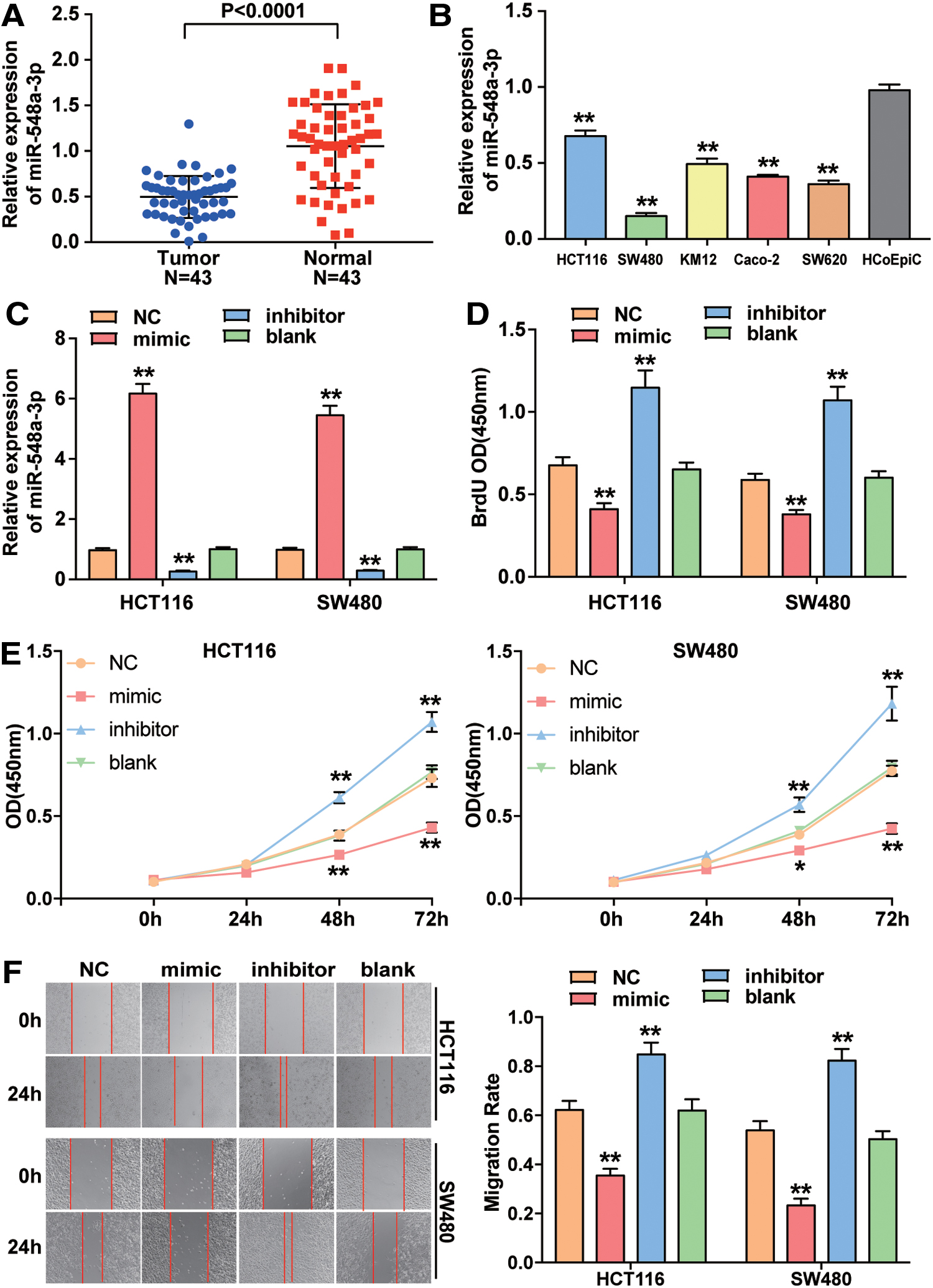
The expression level and function of miR-548a-3p in CC.
Next, HCT116 and SW480 cell lines were transfected with miR-548a-3p mimic, miR-548a-3p inhibitor, or negative control. The transfection efficiency was detected through qRT-PCR. In the miR-548a-3p mimic group, the miR-548a-3p expression increased by approximately sixfold, while the miR-548a-3p expression in the miR-548a-3p inhibitor group decreased by more than 70% compared with the control (blank group) (Fig. 2C). As shown in BrdU DNA incorporation assay, the optical absorbance (OD) of cells was the highest in miR-548a-3p inhibitor group among four groups and the lowest in miR-548a-3p mimic group in HCT116 and SW480 cell lines (Fig. 2D). The cell viability assay showed that the OD in miR-548a-3p inhibitor group was higher than that in blank group, while the cell viability in miR-548a-3p mimic group was weaker than that in blank group in both HCT116 and SW480 cell lines (Fig. 2E). Furthermore, it was observed that upregulating miR-548a-3p led to ∼50% decrease of migration rate, while downregulating miR-548a-3p led to a 1.36 and 1.64-fold increase of migration rate in HCT116 and SW480 cells, respectively (Fig. 2F).
miR-548a-3p directly targeted TPX2 mRNA
The binding site between TPX2 mRNA and miR-548a-3p was predicted by TargetScan Human 7.2 (Fig. 3A). The authors designed luciferase reporter assay to demonstrate whether miR-548a-3p combined with TPX2 mRNA in its' 3′-UTR. The fluorescence intensity was obviously reduced in cells transfected with miR-548a-3p mimic compared with the blank group in the presence of wild-type TPX2 mRNA 3′UTR that contains the binding site of miR-548a-3p. The fluorescence intensity increased in cells transfected with miR-548a-3p inhibitor compared with the blank group in the presence of wild-type TPX2 mRNA 3′UTR. In the presence of TPX2 mRNA 3′UTR mutant, the transfection of miR-548a-3p mimic and inhibitor did not cause a significant change in fluorescence intensity (Fig. 3B).
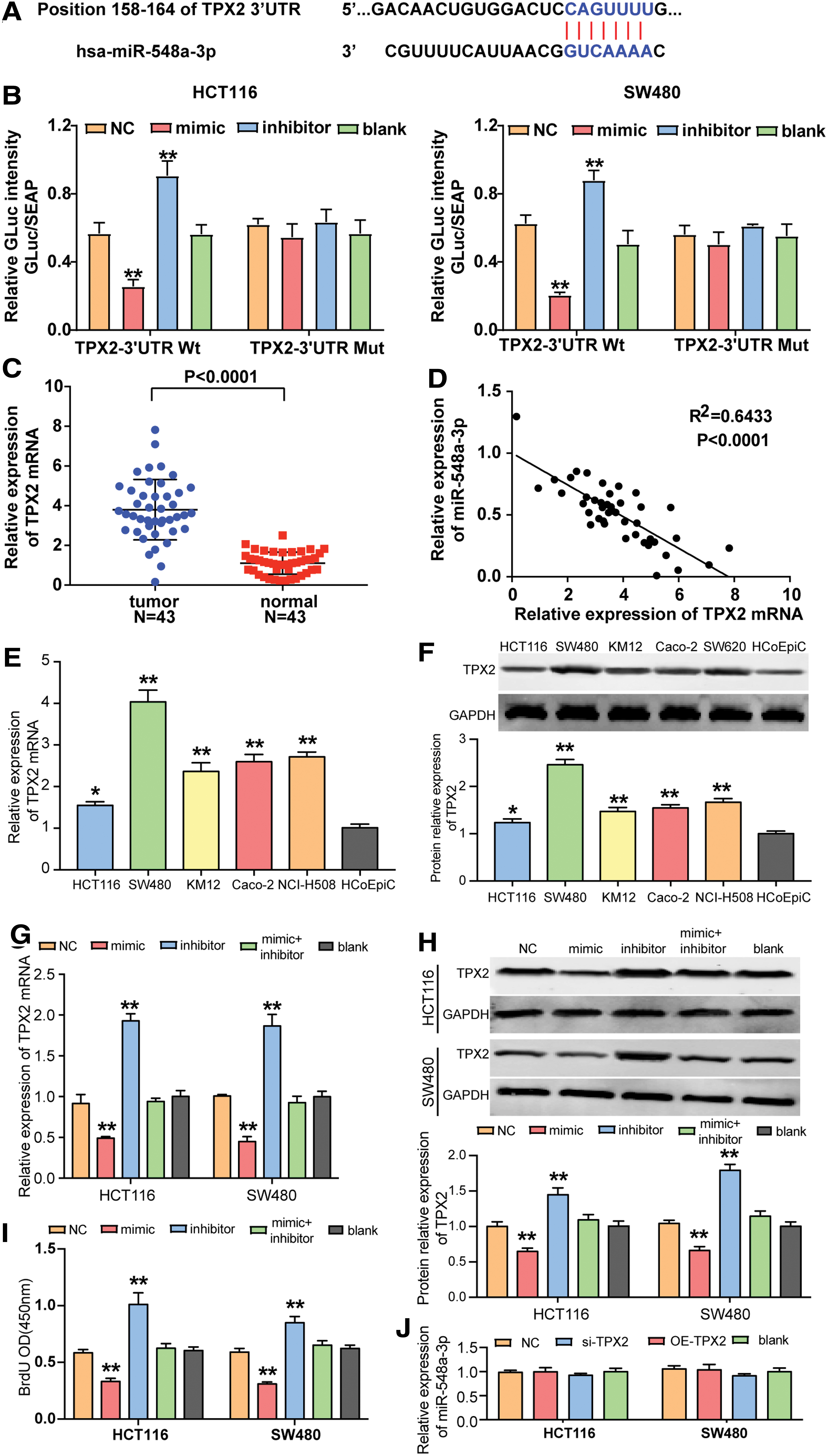
TPX2 was the downstream target gene of miR-548a-3p.
Then, it was found that the expression of TPX2 mRNA in CC tissues was 3.44-fold of that in adjacent healthy tissues (Fig. 3C). To avoid biased statistical test, the authors assigned 23 of the 43 tissues to training set and the other 20 to validation set. The analysis of both sets showed that TPX2 mRNA was significantly upregulated in CC tissues (Supplementary Fig. S1B, D). Besides, it was found that miR-548a-3p had a negative association with TPX2 (Fig. 3D). Similarly, the TPX2 with high expression was observed in five CC cell lines (HCT116, SW480, KM12, Caco-2, and SW620) compared to normal colonic epithelial cell line HCoEpiC through qRT-PCR (Fig. 3E). By detecting the expression of TPX2 protein, they observed that the expression of TPX2 protein significantly upregulated in CC cell lines (Fig. 3F).
Besides, the transfection of miR-548a-3p mimic caused a 1/2 decrease of TPX2 mRNA expression, the transfection of miR-548a-3p inhibitor caused a twofold increase TPX2 mRNA expression, and the cotransfection of miR-548a-3p mimic and inhibitor led to a decrease of TPX2 mRNA expression compared with the transfection of inhibitor alone (Fig. 3G). These results were confirmed at protein level: miR-548a-3p mimic led to almost 50% decrease of the expression of TPX2 protein, miR-548a-3p inhibitor caused almost 1.5-fold increase of TPX2 protein expression, and the cotransfection of mimic and inhibitor led to a decrease of TPX2 protein expression compared with the inhibitor group (Fig. 3H). Besides, BrdU assay results showed that the cotransfection of miR-548a-3p and miR-548a-3p inhibitor caused a reduced proliferation compared with the inhibitor group (Fig. 3I). In addition, the modulation of TPX2 expression did not cause significant changes in miR-548a-3p expression (Fig. 3J). These findings suggest that TPX2 is a downstream target gene of miR-548a-3p.
TPX2 promoted the malignancy phenotypes of CC cells that was regulated by miR-548a-3p
After, the authors confirmed miR-548a-3p′s CC suppressing role and the regulatory relationship between miR-548a-3p and TPX2. It was determined to further study whether miR-548a-3p suppresses the CC cells' phenotypes by regulating TPX2. They thus transfected HCT116 and SW480 cell lines with si-TPX2, OE-TPX2, or cotransfected OE-TPX2 with miR-548a-3p mimic. As shown in Figure 4A, TPX2 expression increased by approximately threefold in OE-TPX2 group, whereas the TPX2 expression decreased by more than 70% in si-TPX2 group in both HCT116 and SW480 cell lines. Besides, the TPX2 expression was markedly downregulated compared with OE-TPX2 group when the HCT116 and SW480 cells were cotransfected with miR-548a-3p mimic and OE-TPX2. Similar to the qRT-PCR results, the Western blot assay showed that the OE-TPX2 led to ∼1.7-fold increase of TPX2 protein level, whereas si-TPX2 led to 50% decrease of the expression of TPX2 protein in HCT116 and SW480 cells. At the same time, the positive effect of OE-TPX2 on the expression of TPX2 protein could be overturned by miR-548a-3p mimic (Fig. 4B).
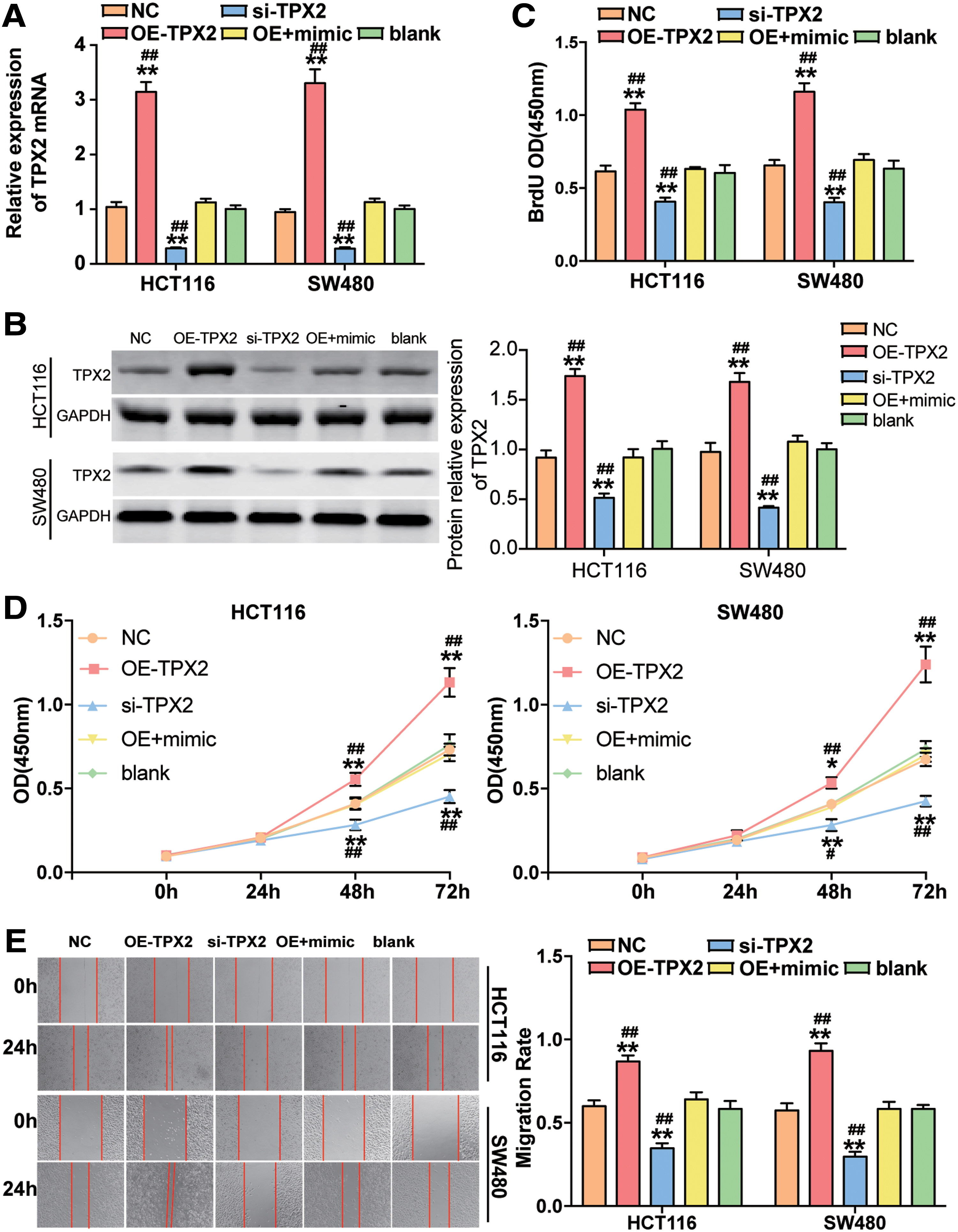
TPX2 promoted cell viability, cell proliferation, and cell migration, which was repressed by miR-548a-3p.
The BrdU assay results displayed that TPX2 overexpression contributed to the cell proliferation, while TPX2 knockdown by si-TPX2 suppressed cell proliferation in HCT116 and SW480 cells. When the authors cotransfected miR-548a-3p mimic and OE-TPX2 into CC cells, the cell proliferation capability was weakened compared with TPX2 overexpression group (Fig. 4B). The result of CCK-8 viability assay exhibited the same trend as the BrdU assay (Fig. 4C). Furthermore, they observed that the migration rate in OE-TPX2 group was enhanced by ∼1.49-fold and 1.6-fold in HCT116 and SW480 cell line, respectively, while the migration rate in si-TPX2 group was declined by ∼50%. When they cotransfected miR-548a-3p mimic and OE-TPX2 into CC cells, the migration rate was significantly declined compared with the transfection of TPX2 overexpression construct (Fig. 4D). These data showed that TPX2 was an oncogene in CC, and its promotion effect could be suppressed by miR-548a-3p.
Discussion
It was observed that miR-548a-3p was downregulated in CC tissues and CC cell lines, while TPX2 was upregulated in CC tissues and CC cell lines. In addition, miR-548a-3p inhibited the malignancy phenotypes of CC cells, while TPX2 acted as the oncogene to contribute to the aggression of CC cells. Furthermore, TPX2 had a negative relationship with miR-548a-3p. Moreover, miR-548a-3p had an ability to reverse the promotion function of TPX2 in viability, proliferation, and migration of CC cells.
miR-548a-3p has been proved to be increased in malignant pleural mesothelioma patients' sera, serving as a promising and novel diagnostic factor for malignant pleural mesothelioma. 18 What's more, another article covered that miR-548a-3p was upregulated in hepatocellular carcinoma patients' sera, acting as a diagnostic and prognostic biomarker for hepatocellular carcinoma. 19 However, a study on miR-548a-3p in breast cancer showed that miR-548a-3p was significantly downregulated in breast cancer tissues, inhibited cell proliferation, and promoted cell apoptosis via targeting ECHS1. 29 In addition, Li et al. 30 found that SIX1 was the oncogene in CC cell line (HCT116) to promote cell proliferation, and it was negatively regulated by miR-548a-3p.
These studies suggest that miR-548a-3p has controversial roles in diverse cancers, and miR-548a-3p could be a tumor suppressor factor in CC because the authors found it significantly downregulated in the collected colon tissue samples and cell lines. They also confirmed that miR-548a-3p significantly suppressed CC cell viability, proliferation, and migration. Thus, the authors' study may contribute to the comprehension of CC mechanism. They reported a novel miR-548a-3p that gets involved in CC pathology and confirmed its CC suppressing role.
TPX2, xklp2 movement binding target protein, is the first-identified cell cycle-associated protein with limited expression and high activity in a variety of malignant tumors. 31,32 TPX2 was once reported to be overexpressed in pancreatic cancer tissues and cells. TPX2 knockdown suppressed pancreatic cancer cell proliferation and tumor growth in nude mice but enhanced cell apoptosis. 26 Another article covered that TPX2 was the downstream target gene of miR-8075, and TPX2 overexpression in cervical cancer contributed to cell proliferation, migration, and invasion. 33 Moreover, TPX2 silence promoted esophageal carcinoma cell cycle arrest in S phase, thereby inhibiting tumor growth in the xenograft nude mice. 34
As for in CC, a study reported that TPX2 with high expression in CC tissues was observed, and TPX2 enhanced cell proliferation, migration, and invasion through enhanced AKT/MMP2 activity. 28 The previous studies suggest that TPX2 is probably an oncogene in several cancers, including CC, and it can be regulated by miRNAs in human cancers. In the study, it was found that upregulated TPX2 promoted the malignancy phenotypes such as migration and proliferation of CC cells, TPX2 knockdown by siRNA technique suppressed these phenotypes. Then, the authors also shed new light on a novel miRNA regulator of TPX2 in CC. Not only did they confirm the tumor facilitator role of TPX2 in CC but also they reported a novel interactome, miR-548a-3p-TPX2, in CC.
In the study, they proved their hypothesis in vitro due to the limited experimental conditions. The influence of TPX2 and miR-548a-3p on CC needs to be further validated in vivo. The bioinformatics analysis displays that TPX2 is the key gene involved in cell cycle especially G2/M transition, which remains to be further studied in CC cells. Besides, the occurrence of CC involves a plethora of factors, the specific signal pathway that TPX2 gets involved in is also worth further investigation.
Conclusion
In summary, the results verified that miR-548a-3p attenuated the development of CC in vitro by downregulating TPX2. miR-548a-3p and TPX2 interactome may be a new target for diagnosing or treating CC in the future.
Footnotes
Authors' Contributions
B.Y.D. designed the study. M.W. and M.Y. performed the experiments and wrote the article. All the authors read and approved the final article.
Ethics Approval
The present study was approved by the Ethics Committee of West China Health Care Hospital of Sichuan University (Wuhan, China). All the patients signed the written informed consent.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.