
Abstract
Background:
Circular RNA (circRNA) can regulate the progression of hepatocellular carcinoma (HCC). However, the role and potential mechanism of circ_0004913 in HCC are not explored.
Methods:
Circ_0004913 was identified from two GSE datasets (GSE94508 and GSE97322) as a differentially expressed circRNA between HCC and normal tissues. Levels of circ_0004913, microRNA-184 (miR-184), and hepcidin (HAMP) were determined by quantitative real-time polymerase chain reaction (qRT-PCR). Cell proliferation, migration, and invasion were estimated by methyl thiazolyl tetrazolium, colony formation, and Transwell assays, respectively. Levels of all proteins were examined by Western blot. Glucose consumption and lactate and ATP production were analyzed by the glucose, lactate, and ATP assay kits. Dual-luciferase reporter, RNA immunoprecipitation (RIP), and RNA pull-down assays were performed to verify the interactions among miR-184 and circ_0004913 or HAMP. The mice xenograft models were established to assess the effect of circ_0004913 on tumor growth in vivo.
Results:
Circ_0004913 was downregulated in HCC, and its expression impeded cell proliferation, migration, and invasion, EMT, and glycolysis in HCC cells. miR-184 was identified as a target miRNA of circ_0004913, and their expression levels were negatively correlated. miR-184 overexpression could reverse the inhibitory effect of circ_0004913 on HCC cell progression. Moreover, as a target gene of miR-184, HAMP expression was positively correlated with circ_0004913 expression in HCC tissues, and repression of miR-184 could inhibit the progression of HCC cells by increasing HAMP expression. Circ_0004913 could inhibit JAK2/STAT3/AKT signaling pathway and tumor growth in vivo by regulating the miR-184/HAMP axis.
Conclusion:
Circ_0004913 inhibited the tumorigenesis of HCC by sponging miR-184 to regulate HAMP expression in vitro and in vivo.
Introduction
Hepatocellular carcinoma (HCC) is a malignant tumor that accounts for more than 90% of primary liver cancers. 1 In 2016, HCC killed ∼110,730 Americans. 2 Currently, HCC treatment included liver resection, liver transplantation, radiofrequency ablation, and drug treatment, such as sorafenib and regorafenib. 3,4 Due to the strong invasion capacity of HCC and the rapid growth of tumor cells, the 5-year survival rate of HCC patients is still low. 5,6 Thus, it is of great significance to explore the pathogenesis of HCC and find effective diagnostic markers to improve clinical efficacy.
As a class of RNA molecules in the noncoding RNA family, circular RNAs (circRNAs) are stable and widely exist in mammalian cells due to their covalently closed-loop structures. 7 CircRNAs were found to play pivotal roles in biological behaviors of tumor cells, such as proliferation, apoptosis, metastasis, and glycolysis. 8,9 Accumulating data indicated that the abundance of circRNA was associated with the progression of HCC and played a biological role through sponging microRNA (miRNA). 10 For example, circRNA FOXP1 facilitated tumor growth in vivo by sponging miR-875-3p and miR-421, and its high expression was closely related to larger tumor size, advanced TNM stage, and poor prognosis. 11 Li et al. revealed that circMAT2B elevated the development of HCC by promoting glycolysis, by regulating the miR-338-3p/PKM2 axis under hypoxia. 12 Recently, Qiu et al. analyzed that circ_0004913 was downregulated in HCC liver tissues by GEO2R method. 13 However, the effect of circ_0004913 and its underlying mechanism in HCC progression are not investigated.
The mechanism by which interaction between circRNA and miRNA regulate cell development has been widely reported in HCC. 14 Accumulating evidence supported that abnormal miRNA expression was associated with the occurrence and development of HCC. For instance, miR-499a and miR-627-5p were significantly curbed in HCC, and could inhibit cell growth, migration, and invasion. 15,16 However, miR-17 could drive the carcinogenesis of HCC by targeting Smad3. 17 Multiple studies reported that miR-184 was raised in HCC and accelerated cell growth. 18 However, it is unclear whether circRNA can co-regulate the development of HCC with miR-184.
Hepcidin encoded through HAMP played a key role in tumor cell proliferation and metastasis. 19 Experiments indicated that maladjusted hepcidin signaling was associated with an augmented risk of HCC. 20 In addition, Shen et al. demonstrated that HAMP hampered cell proliferation and metastasis in HCC. 21 However, the correlation among circRNA, miRNA, and HAMP in HCC remains unclear. In short, the authors aimed to elucidate circ_0004913 expression in HCC and explore its effects on biological behaviors of HCC cells.
Materials and Methods
Samples and cell culture
Thirty HCC samples and their adjacent normal tissues were obtained from HCC patients at Longhua Hospital Shanghai University of Traditional Chinese Medicine. These samples were stored at −80°C until use. Written informed consents were acquired from all patients. This research was approved by the Ethics Committee of the Longhua Hospital Shanghai University of Traditional Chinese Medicine.
Two HCC cell lines (HCCLM3 and Huh7) were obtained from Procell (Wuhan, China), and normal liver cell line THLE-2 was obtained from American Tissue Culture Collection (ATCC, Manassas, VA). All cells were maintained in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS; Invitrogen, Carlsbad, CA) at 37°C under 5% CO2.
Transfection
The full length of circ_0004913 was cloned into the vector (Invitrogen) to form its overexpression plasmid (circ_0004913), and the vector was used as a control. The miR-184 mimics (miR-184), miR-184 inhibitor (anti-miR-184) and the corresponding NCs (miR-NC and anti-miR-NC), and small interference RNA against HAMP (si-HAMP) and its control (si-NC) were bought from RiboBio (Guangzhou, China). Transfection was performed by Lipofectamine 3000 (Invitrogen).
Quantitative real-time polymerase chain reaction
Total RNA from HCC tissues and cell lines at indicated time-points was extracted using Trizol (Invitrogen), and reverse transcription was performed by using PrimeScript RT Reagent Kit (Takara, Wuhan, China). The quantitative real-time polymerase chain reaction (qRT-PCR) was performed using SYBR Select Master Mix (Applied Biosystems, Foster City, CA), and circ_0004913, HAMP, and miR-184 were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and U6, respectively. The primer sequences were listed as follows: circ_0004913, F: 5′-GAAGATCCAAATGAAACTCAGCAA-3′, R: 5′-TCTGCTTTGGCTGTGACATGA-3′; GAPDH, F: 5′-TGATGACCCTTTTGGCTCCC-3′, R: 5′-GAAGCTTGTCATCAATGGAAAT-3′; miR-184, RT: 5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACCCTTAT-3′; miR-184, F: 5′-TGGACGGAGAACTGATAAGGGT-3′, R: 5′-CAGTGCGTGTCGTGGAGT-3′; U6, RT: 5′-ACGCTTCACGAATTTGCGTGTC-3′; U6, F: 5′-CTCGCTTCGGCAGCACA-3′, R: 5′-AACGCTTCACGAATTTGCGT-3′; and HAMP, F: 5′-GCTGCCTGTCTCCTGCTTCT-3′, R: 5′-CTGCAGAGCCGTAGTCTGTCTCGTC-3′. The 2−ΔΔct method was used to calculate the relative expression.
Methyl thiazolyl tetrazolium and colony formation assay
For methyl thiazolyl tetrazolium (MTT) assay, the transfected HCCLM3 and Huh7 cells were cultivated in 96-well plates. At 24, 48, and 72 h after transfection, 10 μL MTT (Beyotime, Shanghai, China) was added to the cells and cells were cultured for another 3 h. Cell proliferation was determined by spectrophotometry at 490 nm.
For colony formation assay, HCCLM3 and Huh7 cells were placed in six-well plates and incubated at 37°C. After 2 weeks, cells were fixed and stained with crystal violet (Solarbio, Beijing, China) for 20 min. Finally, the colonies were counted and photographed under an inverted microscope.
Transwell assay
Transwell assay was performed to detect cell migration and invasion. For invasion assay, Matrigel (Solarbio) was used to coat the upper chambers. For migration assay, the upper chamber was not coated with Matrigel (Solarbio). Briefly, HCCLM3 and Huh7 cells were resuspended in serum-free medium and 100 μL cell suspension was added to the upper chambers, and the lower chambers were added with 600 μL DMEM containing 10% FBS. Thirty-six hours later, cells were washed, fixed, and then stained with 0.1% crystal violet for 30 min. The migrated or invaded cells were counted using an inverted microscope.
Western blot
HCC tissues and cells were collected and lysed by RIPA (Solarbio). The proteins were separated with sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) before being transferred to the polyvinylidene difluoride (PVDF; Beyotime) membranes. The membranes were blocked with 5% skim milk for 2 h, and then incubated with primary antibodies against Ki67 (1:1000; Abcam, Cambridge, MA), proliferating cell nuclear antigen (PCNA, 1:1000; Abcam), Snail (1:1000; Thermo Fisher Scientific, Waltham, MA), vimentin (1:1000; Abcam), hexokinase 2 (HK2, 1:1000; Abcam), lactate dehydrogenase A (LDHA, 1:500; Abcam), glucose transporter 1 (GLUT1, 1:5000; Abcam), HAMP (1:500; Thermo Fisher Scientific), phosphorylation-JAK2 (p-JAK2, 1:1000; Abcam), JAK2 (1:5000; Abcam), p-STAT3 (1:2000; Abcam), STAT3 (1:2000; Abcam), p-AKT (1:1000; Abcam), AKT (1:2000; Abcam), or β-actin (1:2000; Abcam) overnight at 4°C. After washing, the membranes were incubated with the horseradish peroxidase-conjugated (0.3 μg/mL; Abcam) secondary antibodies for 1 h at 37°C. The band signals were visualized using a BeyoECL Plus kit (Beyotime).
Glucose consumption, and lactate and ATP production detection
HCCLM3 and Huh7 cells were seeded into 24-well plates. After 12 h, cells were transfected and then incubated for another 36 h. Next, the cell medium supernatant was collected and the concentrations of glucose and lactate were determined by the glucose and lactate assay kits (Sigma-Aldrich, St. Louis, MO). The level of ATP was detected by the ATP assay kit (Invitrogen).
Dual-luciferase reporter assay
Wild-type circ_0004913 (circ_0004913-WT), mutant circ_0004913 (circ_0004913-MUT), HAMP 3′UTR-WT, and HAMP 3′UTR-MUT were co-transfected into HCCLM3 and Huh7 cells with miR-184 or miR-NC, respectively. At 48 h after transfection, the luciferase activity was evaluated using a dual-luciferase reporter assay system (Promega Corporation, Madison, WI).
RNA immunoprecipitation assay
RNA immunoprecipitation (RIP) experiments were conducted by a Magna RIP RNA-binding Protein Immunoprecipitation Kit (Millipore, Billerica, MA). HCCLM3 and Huh7 cells were lysed in lysis buffer and incubated with the magnetic beads-antibody complex. The antibodies were Ago2 and IgG (control). The RNA from the magnetic beads binding complexes were extracted for subsequent qRT-PCR detection.
RNA pull-down assay
HCCLM3 and Huh7 cells were transfected with biotinylated miR-NC (Bio-miR-NC) or Bio-miR-184 and cultured for 48 h. Cells were lysed in lysis buffer for 10 min and then incubated with M-280 streptavidin beads (Sigma-Aldrich) overnight. Then, cells were washed by low and high salt buffer, and the RNA bound to the beads was extracted. Finally, the enrichment of circ_0004913 was analyzed by qRT-PCR.
Subcutaneous xenograft model
To establish xenograft models in nude mice, HCCLM3 cells stably transfected with circ_0004913 or vector were subcutaneously injected into the 4-week-old male nude mice, and the mice were divided into two groups (n = 4 each group): circ_0004913 and vector. Tumor volume was measured every other week. At 5 weeks after injection, all mice were euthanized and tumor weight was detected. Furthermore, tumors were excised for further qRT-PCR and Western blot analysis. The animal experiments were permitted by the Animal Care and Use Committee of Longhua Hospital Shanghai University of Traditional Chinese Medicine.
Statistical analysis
SPSS 19.0 and GraphPad Prism 7 were employed for data analysis. Data were shown as mean ± standard deviation (SD) from at least three times independently. Student's t-test and one-way analysis of variance (ANOVA) were, respectively, analyzed by the differences between the two groups and among multiple groups. The correlation was estimated by Pearson correlation coefficient. p < 0.05 was considered to have significant difference.
Results
Circ_0004913 was downregulated in HCC tissues and cells
To identify specific circRNAs in HCC, the authors analyzed two microarray datasets (GSE94508 and GSE97322) downloaded from the GEO database, and the first 10 downregulated and the last 10 upregulated circRNAs were selected from the two databases (Fig. 1A, B). Among the circRNAs, circ_0004913 was the only circRNA appearing in both GSE datasets (Fig. 1C). Thus, circ_0004913 was selected for further study. Then, they examined circ_0004913 expression in 30 pairs of HCC tissues and adjacent normal tissues by qRT-PCR. As shown in Figure 1D, circ_0004913 expression was remarkably decreased in HCC tissues compared with normal tissues. Circ_0004913 expression in HCC cell lines was also measured by qRT-PCR. The data showed that circ_0004913 was downregulated in HCC cell lines (HCCLM3 and Huh7) compared with normal cell line THLE-2 (Fig. 1E). These results supported that circ_0004913 might be an important regulator in HCC progression.

Circ_0004913 was downregulated in HCC tissues and cells.
Circ_0004913 overexpression inhibited the progression of HCC cells in vitro
To explore the biological role of circ_0004913 in HCC development, the authors overexpressed the expression of circ_0004913 in HCCLM3 and Huh7 cells by transfection of circ_0004913. As shown in Figure 2A, circ_0004913 transfection significantly increased circ_0004913. MTT results showed that circ_0004913 overexpression suppressed the proliferation of HCCLM3 and Huh7 cells (Fig. 2B, C). Colony formation assay demonstrated that circ_0004913 upregulation let to a distinct reduction of the colony number of HCCLM3 and Huh7 cells (Fig. 2D). Transwell assay revealed that overexpression of circ_0004913 decreased the number of migrated and invaded cells (Fig. 2E, F). In addition, they examined the expression levels of proliferation marker proteins Ki67 and PCNA as well as EMT-related proteins Snail and vimentin by Western blot. The data indicated that upregulation of circ_0004913 in HCCLM3 and Huh7 cells decreased the expression levels of these proteins (Fig. 2G), suggesting that the increased circ_0004913 could repress the proliferation and EMT of HCC cells. To examine the effect of circ_0004913 on cell glycolysis, the authors assessed the glucose consumption, and lactate and ATP production in HCC cells transfected with circ_0004913. As displayed in Figure 2H, highly expressed circ_0004913 reduced the glucose consumption. Consistently, lactate and ATP production were also decreased in HCCLM3 and Huh7 cells with overexpressed circ_0004913 (Fig. 2I, J). Meanwhile, Western blot demonstrated that the expression of glycolysis-related proteins HK2, LDHA, and GLUT1 was significantly degraded by overexpressing circ_0004913 in HCCLM3 and Huh7 cells. These findings suggested that circ_0004913 overexpression suppressed the proliferation, metastasis, and glycolysis of HCC cells in vitro.
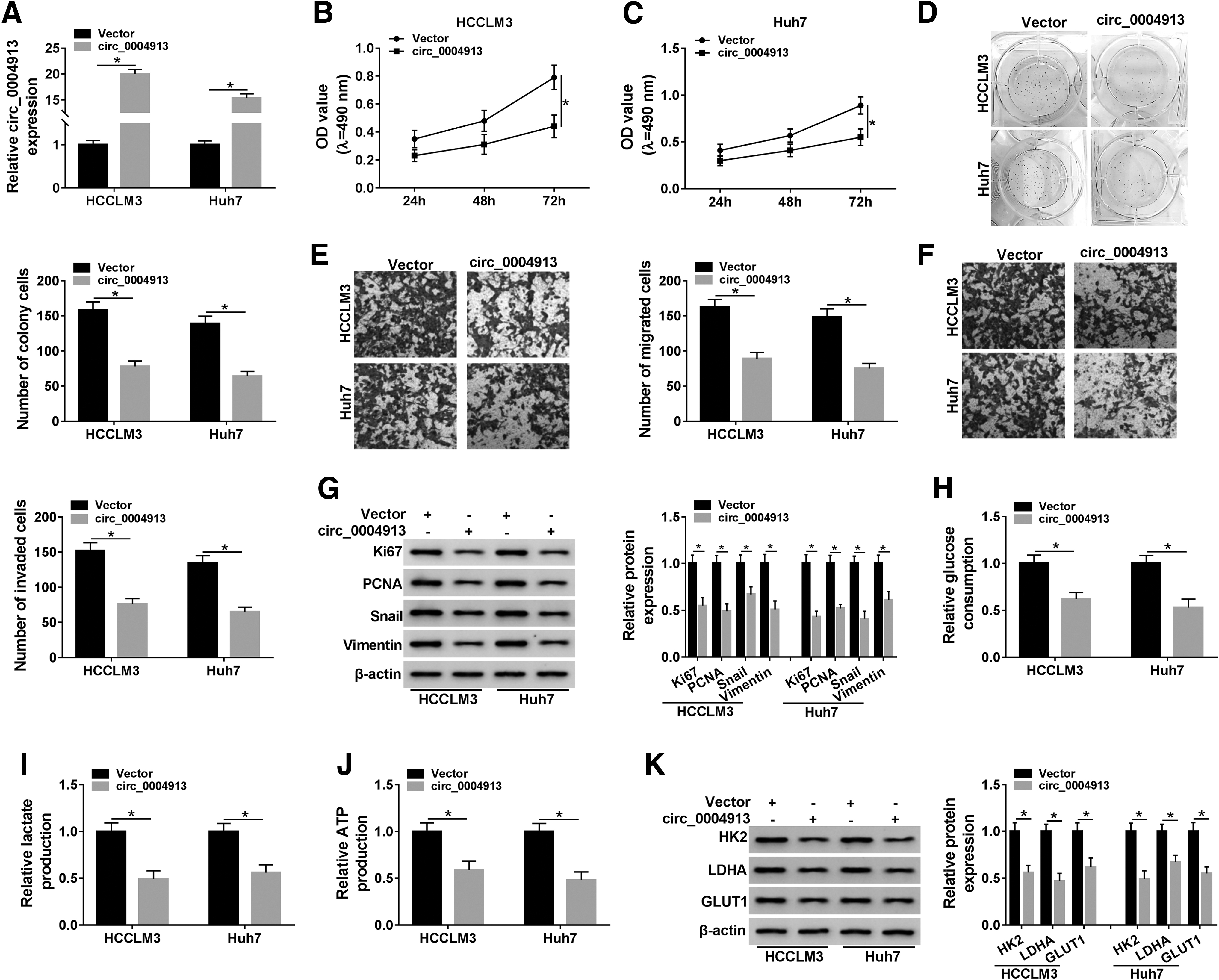
Circ_0004913 overexpression inhibited the progression of HCC cells in vitro. HCCLM3 and Huh7 cells were transfected with vector or circ_0004913.
Circ_0004913 could act as a sponge for miR-184 in HCC cells
CircRNA can function as the molecular sponge by affecting the expression of miRNAs. 22 Therefore, the authors analyzed the underlying targets of circ_0004913 through bioinformatics method CircInteractome and found that miR-184 has complementary sites with circ_0004913 (Fig. 3A). They therefore performed dual-luciferase reporter assay in HCCLM3 and Huh7 cells. Transfection of miR-184 significantly increased the expression of miR-184 (Fig. 3B). As shown in Figure 3C and D, miR-184 exerted the inhibitory impact on the luciferase activity in circ_0004913-WT group, while showed no significant effect on the luciferase activity in circ_0004913-MUT group. RIP assay validated that circ_0004913 and miR-184 were both enriched in HCCLM3 and Huh7 cells incubated with Ago2 antibody compared to the control IgG group (Fig. 3E, F). RNA pull-down also confirmed that the enrichment level of circ_0004913 in Bio-miR-184 group was higher than that in Bio-NC group (Fig. 3G), suggesting that miR-184 was the target of circ_0004913. Besides, the expression of miR-184 was found to be upregulated in HCC tissues and cell lines (Fig. 3H, I), and its expression was negatively correlated with circ_0004913 expression in HCC tissue samples (Fig. 3J). These data demonstrated that circ_0004913 could bind to miR-184 in HCC cells.
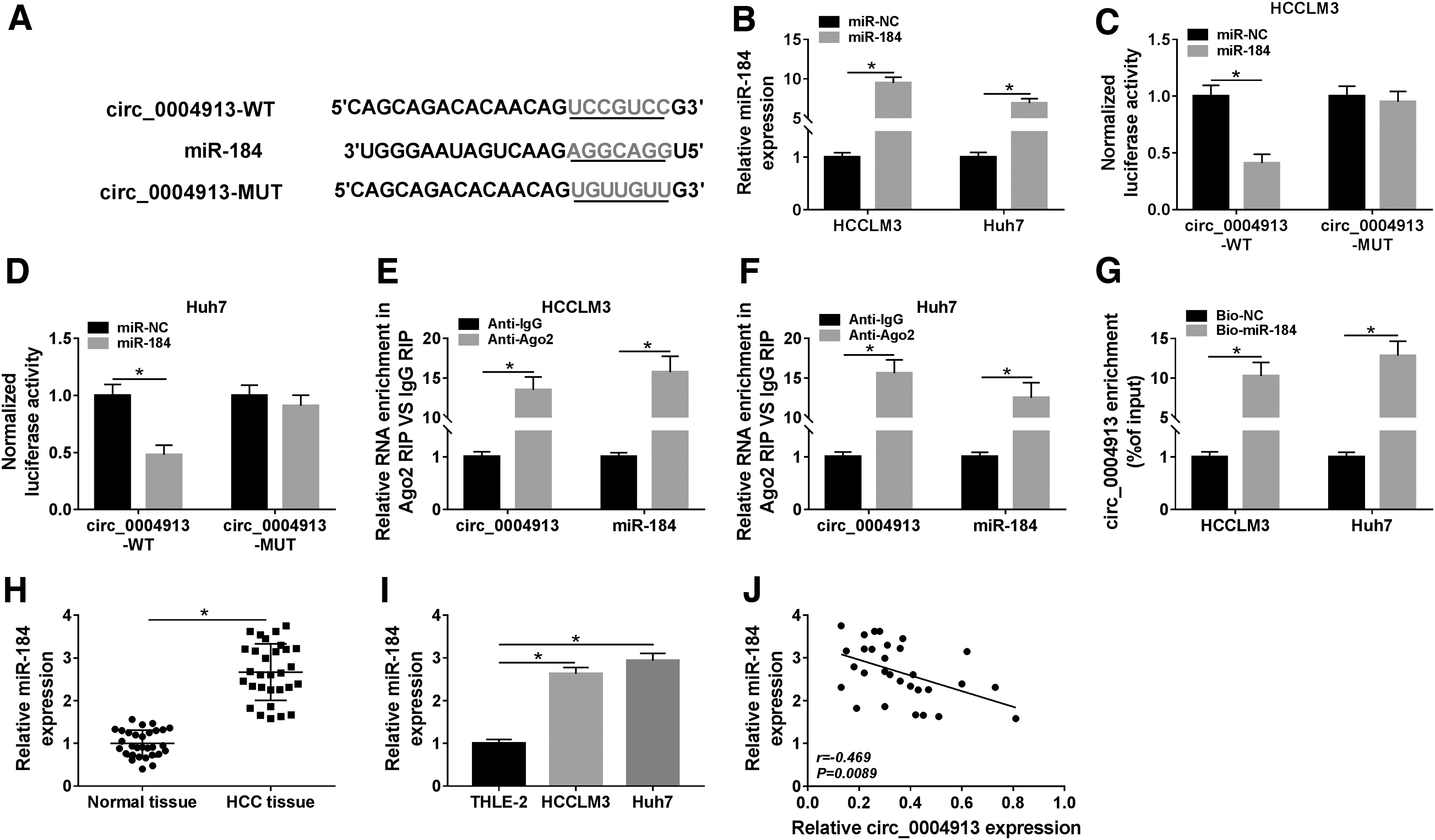
Circ_0004913 could act as a sponge for miR-184 in HCC cells.
Circ_0004913 restrained the malignant biological behaviors of HCC cells by suppressing miR-184
The authors employed the rescue experiments to investigate whether circ_0004913 regulated the biological behaviors of HCC cells by targeting miR-184. As appeared in Figure 4A, circ_0004913 overexpression distinctly suppressed miR-184 expression in HCCLM3 and Huh7 cells. Moreover, circ_0004913 overexpression repressed the proliferation (Fig. 4B, C), colony number (Fig. 4D), migration (Fig. 4E), and invasion (Fig. 4F) of HCCLM3 and Huh7 cells, while these effects could be reversed by co-transfection of miR-184. Furthermore, the inhibition effects of circ_0004913 on the expression of Ki67, PCNA, Snail, and vimentin were counteracted by miR-184 overexpression in HCCLM3 and Huh7 cells (Fig. 4G). Similarly, co-transfection with miR-184 in HCCLM3 and Huh7 cells could rescue the suppressive effects of circ_0004913 on the levels of glucose consumption, and lactate and ATP production (Fig. 4H–J). As expected, overexpression of miR-184 promoted the protein levels of HK2, LDHA, and GLUT1 in HCCLM3 and Huh7 cells transfected with circ_0004913 (Fig. 4K). Taken together, these results confirmed that circ_0004913 hampered HCC cell proliferation, metastasis, and glycolysis by decreasing miR-184 expression.

Circ_0004913 restrained the malignant biological behaviors of HCC cells by suppressing miR-184.
HAMP was the target of miR-184
To figure out the mechanism of circ_0004913/miR-184 in HCC progression, the authors filtrated the potential targets of miR-184 by GSE88839 and GSE101728 microarray datasets (ranking among the top 100 downregulated genes in HCC patient samples) as well as StarBase v3.0 tool, and the predicted results were presented in a Venn diagram (Fig. 5A). Only one gene HAMP appeared in the intersection of the three data sets, and the binding sites between miR-184 and HAMP are shown in Figure 5B. To prove their interaction, dual-luciferase reporter assay and RIP assay were carried out in HCCLM3 and Huh7 cells. The results showed that miR-184 remarkably reduced the luciferase activity of the HAMP 3′UTR-WT, while these was no significant change in the luciferase activity of HAMP 3′UTR-MUT group (Fig. 5C, D). Similarly, RIP results showed that miR-184 and HAMP were prominently enriched in HCCLM3, and Huh7 cells conjugated Ago2 antibody compared with the control IgG group (Fig. 5E, F). Importantly, miR-184 overexpression in HCCLM3 and Huh7 cells significantly degraded HAMP expression at mRNA and protein levels (Fig. 5G, H). Contrary to miR-184 expression in HCC, the mRNA and protein levels of HAMP were evidently declined in HCC tissues and cell lines (Fig. 5I–L), and a significant inverse correlation between miR-184 and HAMP was also observed (Fig. 5M). Moreover, HAMP expression was positively correlated with the circ_0004913 expression in HCC tissues (Fig. 5N). The above findings revealed that miR-184 directly targeted HAMP and regulated its expression in HCC cells.
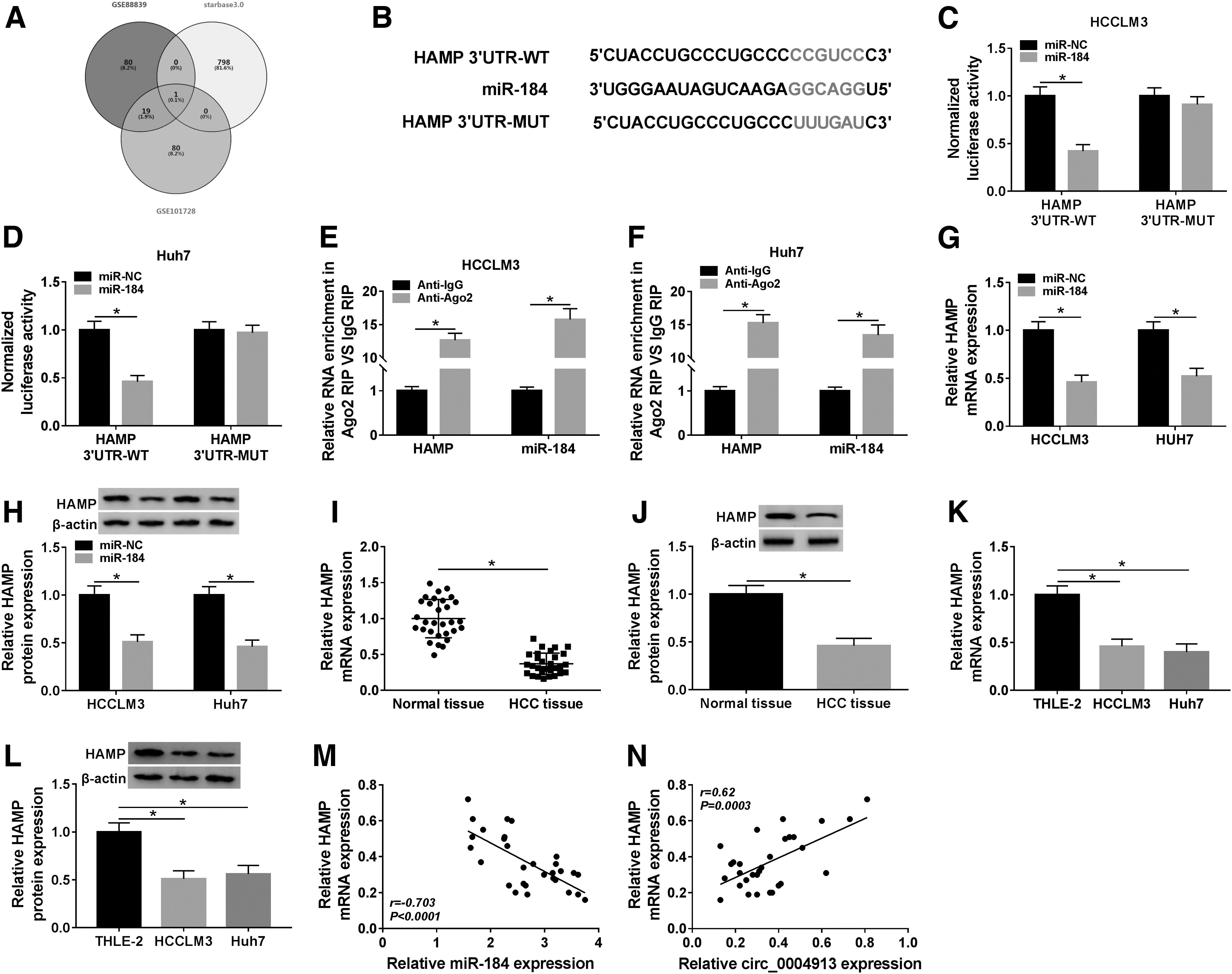
HAMP was the target of miR-184.
Silencing HAMP reversed the inhibitory effect of miR-184 knockdown on the progression of HCC cells
To demonstrate whether miR-184 regulated HCC cell progression by HAMP, the authors performed rescue assays. First, qRT-PCR and Western blot were used to detect the interference efficiency of HAMP by transfection of si-HAMP in HCCLM3 and Huh7 cells, and the results showed that HAMP expression was markedly decreased at mRNA and protein levels in cells transfected with si-HAMP (Fig. 6A, B). Subsequently, the authors found that repression of miR-184 retarded cell proliferation (Fig. 6C, D) and inhibited cell cloning (Fig. 6E), migration (Fig. 6F), and invasion (Fig. 6G) in HCCLM3 and Huh7 cells, while these effects were abolished by interfering with HAMP. Simultaneously, HAMP knockdown reversed the inhibition effects of anti-miR-184 on the expression of Ki67, PCNA, Snail, and vimentin (Fig. 6H, I). Additionally, glucose consumption (Fig. 6J), and lactate (Fig. 6K) and ATP production (Fig. 6L) were reduced by suppression of miR-184, and HAMP deficiency could invert these effects. In accordance with expectation, miR-184 knockdown decreased the expression levels of HK2, LDHA, and GLUT1 in HCCLM3 and Huh7 cells, while HAMP depletion enhanced their expression levels (Fig. 6M, N). To sum up, miR-184 knockdown impeded the progression of HCC cells by increasing HAMP expression.

Silencing HAMP reversed the inhibitory effect of miR-184 knockdown on the progression of HCC cells.
Overexpression of circ_0004913 could inhibit JAK2/STAT3/AKT signaling pathway through the miR-184/HAMP axis
To explore whether the JAK2/STAT3/AKT signaling pathway was involved in the inhibition of circ_0004913 on the progression of HCC cells, the protein expression implicated in the JAK2/STAT3/AKT signaling pathway was detected by Western blot. As shown in Figure 7A and B, overexpression of circ_0004913 in HCCLM3 and Huh7 cells could reduce the ratio of p-JAK2/JAK2, p-STAT3/STAT3, and p-AKT/AKT, and increase the protein expression of HAMP, while these reduction effects were reversed by overexpressing miR-184 or silencing HAMP, suggesting that circ_0004913 was able to inactivate JAK2/STAT3/AKT signaling pathway through the miR-184/HAMP axis.

Overexpression of circ_0004913 could inhibit JAK2/STAT3/AKT signaling pathway through the miR-184/HAMP axis. HCCLM3 and Huh7 cells were transfected with vector, circ_0004913, circ_0004913 + miR-NC, circ_0004913 + miR-184, circ_0004913 + si-NC, or circ_0004913 + si-HAMP.
High circ_0004913 expression hindered HCC tumor growth in vivo
To appraise the role of circ_0004913 in tumor growth in HCC in vivo, the authors constructed a mouse xenograft model. HCCLM3 cells transfected with circ_0004913 or vector (control) were subcutaneously injected into the nude mice. The body weight of the mice was shown in Figure 8A, and the data suggested that circ_0004913 had no effect on the body weight of the mice. The results showed that high expression of circ_0004913 inhibited tumor growth, reflecting smaller tumor size and weight compared to control group (Fig. 8A, B). Moreover, qRT-PCR results showed that the expression levels of circ_0004913 (Fig. 8C) and HAMP (Fig. 8E) were upregulated, while miR-184 was downregulated (Fig. 8D) in circ_0004913 group. Meanwhile, the protein expression of HAMP was also found to be increased in tumor tissues of circ_0004913 group (Fig. 8F). These data showed that circ_0004913 overexpression suppressed the growth of HCC tumors by regulating the miR-184/HAMP axis in vivo.

High circ_0004913 expression hindered HCC tumor growth in vivo. HCCLM3 cells transfected with circ_0004913 or vector (control) were subcutaneously injected into the nude mice.
Discussion
Due to the high recurrence and metastasis rates, the prognosis of HCC is worrisome. 23 Therefore, it is necessary to study the molecules and mechanisms associated with HCC metastasis. Accumulating evidence signified that circRNAs were involved in the growth and metastasis of human cancer cells, including HCC. 24 In this study, circ_0004913 was identified as a HCC-related circRNA in two GSE datasets by GEO2R tool, and the authors validated that circ_0004913 was downmodulated in HCC tissues and cells. Moreover, the results demonstrated that circ_0004913 overexpression could suppress cell proliferation, migration, and invasion. The degree of glycolysis can reflect the metabolic level of cancer cells. 25 Glucose consumption, and lactate and ATP production are three indicators of glycolysis, and HK2, LDHA, as well as GLUT1 are important enzymes in glycolysis pathway. 26 In these data, circ_0004913 retarded glycolysis by reducing glucose consumption, and lactate and ATP production. These data implied that circ_0004913 might be a tumor suppressor in HCC progression and a potential molecular marker for the treatment of HCC.
In general, circRNAs can bind to their targeted miRNAs and serve as ceRNAs to regulate mRNA expression. 27 For instance, circ_0067934 regulated bladder cancer progression through inhibiting miR-1304. 28 Li et al. indicated that circMYLK modulated miR-513a-5p/VEGFC axis to enhance cell growth and metastasis in renal cell carcinoma. 29 The authors' results suggested that miR-184 was a downstream target gene of circ_0004913. Besides, miR-184 was confirmed to be increased in HCC. Previously, Long et al. validated that miR-184 was upregulated in HCC and interference with miR-184 restrained cell growth and migration. 30 Wu et al. reported that miR-184 was aggrandized in HCC and elevation of miR-184 promoted cell proliferation and tumorigenesis. 31 In line with these results, the authors observed that overexpression of miR-184 could reverse the effects of circ_0004913 on HCC cells. Taken together, circ_0004913 acted as an anticancer factor in HCC by downregulating miR-184.
Moreover, the authors proved that HAMP was the target mRNA of miR-184 and negatively regulated by miR-184 expression. Importantly, they found a positive correlation between HAMP and circ_0004913. HAMP mainly encodes hepcidin, an antimicrobial peptide hormone involved in the metabolism of iron. High levels of iron in cells could interfere with the cancer cell cycle, 32 and silencing HAMP induced cell proliferation and migration. 21 In this research, HAMP knockdown could contribute to the progression of HCC cells. Chung et al. indicated that hepcidin expression was associated with JAK2/STAT3 signaling pathway. 33 The authors' findings showed that circ_0004913 could impair the expression of phosphorylated proteins JAK2, STAT3, and AKT, and these effects were abated by increasing miR-184 or silencing HAMP. Consistent with in vitro results, circ_0004913 overexpression hampered tumor growth in vivo by regulating the miR-184/HAMP axis.
Conclusion
In summary, the authors revealed that circ_0004913 served as an antitumor factor in HCC. In addition, circ_0004913 partially inactivated the JAK2/STAT3/AKT signaling pathway through miR-184/HAMP axis, thus restraining the proliferation, metastasis, and glycolysis of HCC cells.
Footnotes
Acknowledgment
The authors sincerely appreciate all members participated in this study.
Authors' Contributions
Conception and design: Mingyuan Wu; development of methodology: Tanlezi Sun; acquisition of data: Mingyuan Wu; analysis and interpretation of data: Tanlezi Sun and Lianjun Xing; writing, review, and revision of article: Mingyuan Wu; and administrative, technical, and material support: Lianjun Xing. All coauthors have reviewed and approved the article before submission.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary data of Western blots.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.