
Abstract
Background:
Various types of tumors are likely to acquire drug resistance over time. Hence, the development of novel therapies to overcome drug resistance is critical. Studies have demonstrated that drug resistance is closely associated with the dynamic regulation of mitochondria in tumor cells. The dynamin-related protein 1 (Drp1) is involved in the regulation of mitochondrial fission and plays an important role in maintaining mitochondrial morphology, function, and distribution. It is a key protein in mitochondrial quality control. Drp1 is a GTPase localized to the cytoplasm and is a potential target in cancer therapy. A variety of drugs targeting Drp1 have shown great promise in reducing the viability and proliferation of cancer cells. The dynamic regulation of Drp1-mediated mitochondria is closely associated with tumor development, and treatment.
Aim:
In this article, the authors reviewed the occurrence and progression of mitochondrial fission regulated by Drp1, and its influence on cell cycle, autophagy, apoptosis, migration, invasion, the molecular mechanism of tumor stemness, and metabolic reprogramming. Targeted inhibition of Drp1 and mitochondrial fission could reduce or prevent tumor occurrence and progression in a variety of cancers. Drp1 inhibitors could reduce tumor stemness and enhance tumor sensitivity to chemotherapeutic drugs.
Conclusion:
Research into identifying compounds that could specifically target Drp1 will be valuable for overcoming drug resistance in tumors.
Introduction
Mitochondrial quality control (MQC) is closely associated with aging, cancer, and neurodegenerative diseases. MQC refers to monitoring mitochondria at the molecular, organelle, and cellular level to ensure mitochondrial integrity and mitochondrial protein balance. The monitoring mechanism comprises the elimination of misfolded and damaged proteins at the molecular level by proteases in the mitochondria, maintaining protein levels in the mitochondria, and ensuring the synthesis and stability of the mitochondria. 1 At the organelle level, mitochondrial function is regulated by mitochondrial fission and fusion by kinesis-related proteins and mitochondrial fusion proteins.
Cells will eliminate irreversibly damaged mitochondria through a process termed mitophagy, which is an important mitochondrial control mechanism. In addition to organelle dynamics, the continual interaction of mitochondria with the ubiquitin-proteasome system has been identified as an important aspect of MQC. 2
Dynamin-related protein 1 (Drp1) is important for regulating mitochondrial fission. Coimmunoprecipitation assays have demonstrated that Fis1 and Drp1 bind with each other. The mitochondrial outer membrane proteins Fis1/Mdv2 recruits Drp1 from the cytoplasm and then associates with other proteins to form a large cleavage complex. The multimeric loop structure of Drp1 is gradually tightened causing the mitochondria to split. 3 Proteins involved in mitochondrial fusion include Mfn1/2 and Opa1. During mitochondria fusion, Mfn1/2 accumulates at the fusion site. Mfn1/2 proteins from two mitochondria then bind to each other through homologous or heteromultimers through their coiled-coil domains. This then guides the two mitochondria to gradually fuse into one. 4 The division and fusion of mitochondria work synergistically to maintain the dynamic regulation of mitochondria and achieve MQC. Drp1 is a key protein that regulates mitochondrial fission, and its abnormal expression is closely associated with tumor progression.
Drp1 and Its Involvement in Tumor Initiation and Development
Abnormal expression of Drp1 was observed during tumor progression. 5 Inhibition of Drp1 or augmentation of mitofusin-2 increases apoptosis and decreases proliferation in cancer cells. 5 The role of Drp1-mediated mitochondrial dynamics in the development of cancer is primarily associated with mitochondrial fission during mitosis. The coordination between mitochondrial fission and mitosis ensures equal distribution of mitochondria to daughter cells. 6 During the transition from the G1 to the S phase, mitochondria fuse and increase ATP production. DNM1L inhibition induces mitochondrial hyperfusion and triggers DNA replication and Cyclin E accumulation. 7
Drp1 mediates mitochondrial fission to accelerate mitosis and inhibit mitochondrial calcium influx to prevent calcium-mediated apoptosis. 8 Kashatus et al. demonstrated that phosphorylation of Drp1 at Ser616 regulated by mitogen-activated protein kinase (MAPK) is a major reason for early tumor progression. 9 Cancer cells often exhibit fragmented mitochondria. High expression or activated Drp1 has been shown to mediate lung cancer, 10 metastatic breast cancer, 11 glioblastoma, 12 neuroblastoma, 13 colon-rectal cancer cells, 14 melanoma 9 and other types of tumors.
Drp1-mediated mitochondrial fission is hypothesized to reduce the activity of mitochondrial complex I, enhance glycolysis, reduce reactive oxygen species (ROS) accumulation and response to cytotoxic therapies. 15 Studies have shown that MAPK signaling activates Drp1 via extracellular regulated protein kinases (ERK)-mediated phosphorylation, which is critical for Ras-induced malignant transformation of cells. 16 Mutations in the MAPK signaling pathway causes cancer cells to metabolize from oxidative phosphorylation (OXPHOS) to anaerobic glycolysis, that is, Warburg effect. Mitochondrial fission induced by elevated Drp1 activity is a consequence of carcinogenic signal RASG12V. This gives rise to mitochondrial dysfunction, metabolic reprogramming, increased glycolysis flux, and glutamate decomposition. 9
Progression of Antitumor Studies: The Relationship of Drp1 with Tumor Characteristics
Drp1 regulation of mitochondrial dynamics has been closely associated with 10 major tumor characteristics, 17 and were mainly involved in cell cycle progression, apoptosis, autophagy, migration and invasion, stemness, and metabolic reprogramming, all of which play significant roles in tumorigenesis.
Drp1 and the cell cycle in tumor cells
An imbalance in the cell cycle is a major feature observed in tumor cells. Drp1 plays an important role in regulating the cell cycle. During the G1/S phase, mitochondria are highly depolarized and fuse to form single giant tubular structures to preserve energy for DNA synthesis. It contributes to the recombination of mtDNA and maintains mitochondria between the daughter cells. During the S phase, mitosis-associated kinases phosphorylate Aurora A, which then causes RalA to localize to the mitochondria to recruit receptor RalBP1.
RalBP1 acts as a scaffold for CyclinB/cyclin-dependent kinases (CDK) 1 in the mitochondria. CyclinB/CDK1 then phosphorylates Drp1 at Ser616 to induce mitochondrial cleavage. 16 Drp1 inhibition causes sustained high-fusion mitochondria and induce a stress response to DNA replication. This in turn causes ataxia telangiectasia mutated–dependent DNA damage repair and arrests cell mitosis during the G2/M phase. This is one of the molecular mechanisms of Drp1 regulation of the cell cycle. 18 In addition, recent studies have shown that the cell cycle regulator Cyclin E1 could be recruited to the mitochondria. Reduction in Drp1 leads to the degradation of mitochondria-localized Cyclin E1, which results in cell cycle arrest. This further illustrates the important regulatory role of Drp1 in the cell cycle. 19
Drp1-mediated mitochondrial fission regulates the tumor cell cycle. In lung cancer cells, the activated form of Drp1, p-Drp1 Ser616, is increased compared with normal lung epithelial cells, while the expression of p-Drp1 Ser637 is decreased in the inactive form. The inhibition of p-Drp1 Ser616 results in S-phase arrest in lung cancer cells and a decrease in proliferation. This indicates that Drp1 plays an important role in the regulation of the tumor cell cycle. 10 PINK1 is an important kinase that phosphorylates and activates Drp1. Knockdown of PINK1 in breast cancer cells can inhibit Drp1 phosphorylation, reduce its activity, and reduce mitochondrial fission. This leads to G2/M arrest and demonstrates that Drp1 could exert antitumor effects by regulating the cell cycle. 20 Zhan et al. showed that high expression of Drp1 increases mitochondrial fission to promote the proliferation of hepatocellular carcinoma (HCC) cells in vitro and in vivo. This was mainly through increased G1/S progression through the cell cycle. Drp1 knockdown or treatment with a mitochondrial fission inhibitor-1 (Mdivi-1) significantly induced G1 arrest in HCC cells and inhibited tumor growth in an in vitro xenograft model. Chen et al. observed that PSII induced colorectal cancer (CRC) cell cycle arrest in the G1 phase by inhibiting the phosphorylation of ERK1/2 and mitochondrial translocation of Drp1 by dephosphorylating Drp1 at Ser616. 21 Additional studies have demonstrated that Drp1-mediated mitochondrial fission promotes the proliferation of hepatoma cells mainly through the p53/p21 and NF-κB cell cycle signaling pathways. Hence targeting Drp1-dependent mitochondrial fission provides a new strategy for inhibiting HCC tumor growth 22 (Table 1).
Dynamin-Related Protein 1 and Tumor Development
CRC, colorectal cancer; Drp1, dynamin-related protein 1; HCC, hepatocellular carcinoma; ROS, reactive oxygen species.
Drp1 and mitophagy in tumor cells
Tumor cells degrade damaged organelles and misfolded proteins in the cytoplasm by increasing autophagy. This is turn generates raw materials and energy required for cell metabolism. Hence autophagy is important for tumor cell survival. 23 Drp1 is mainly involved in mitochondrial autophagy. Low mitochondrial membrane potential and dysfunction leads to increased mitophagy, with Drp1-mediated mitochondrial fission being an important mechanism. 24 Recent studies have demonstrated that adenylate-activated protein kinase promotes mitochondrial translocation of Drp1 and induces increased mitochondrial fission leading to increased mitophagy. 25 Si et al. found that Silibinin-induced mitochondrial fission leads to Drp1-related mitophagy, which attenuates apoptosis in MCF-7 and MDA-MB-231 cells. 26 In addition, Jin et al. found that Drp1 induced mitophagy and suppressed apoptosis after irradiation with 0.5 Gy carbon ions in MCF-7 and MDA-MB-231 cells, 27 whereas treatment with Mdivi-1 significantly reduced the growth of HCC tumors in vitro. The main mechanism was through the inhibition of Drp1-mediated mitophagy. 28
Hence, the increase in mitophagy induced by Drp1 is one of the mechanisms by which tumor cells become less sensitive to chemotherapeutic drugs. However, some studies have demonstrated that Drp1-regulated mitophagy promotes cancer cell death and inhibits tumor proliferation.
A decrease in Drp1 levels accelerates mitochondrial fusion induced by mitophagy, which subsequently attenuates mitochondrial OXPHOS resulting in the inhibition of tumor growth in pancreatic ductal adenocarcinomas (PDAC). 29 Yadav et al. found that knockdown of scrib leads to elevated Drp-1 levels and enhanced mitochondrial fission, whereas reduced Parkin expression and defective mitophagy leads to tumorigenesis in the wing imaginal tissues of Drosophila (TWITD). 30 Thomas et al. indicated that HPV-E7 oncoprotein relieves E2F5, which then associates with Drp1 resulting in increased mitochondrial localization of Drp1 to induce mitophagy and tumor suppression in oropharynx head and neck squamous cell carcinomas (HNSCC). 31 In addition, BAY 87–2243 has been demonstrated to trigger mitophagy and lead to necroptosis and ferroptosis in melanoma cells 32 (Fig. 1).
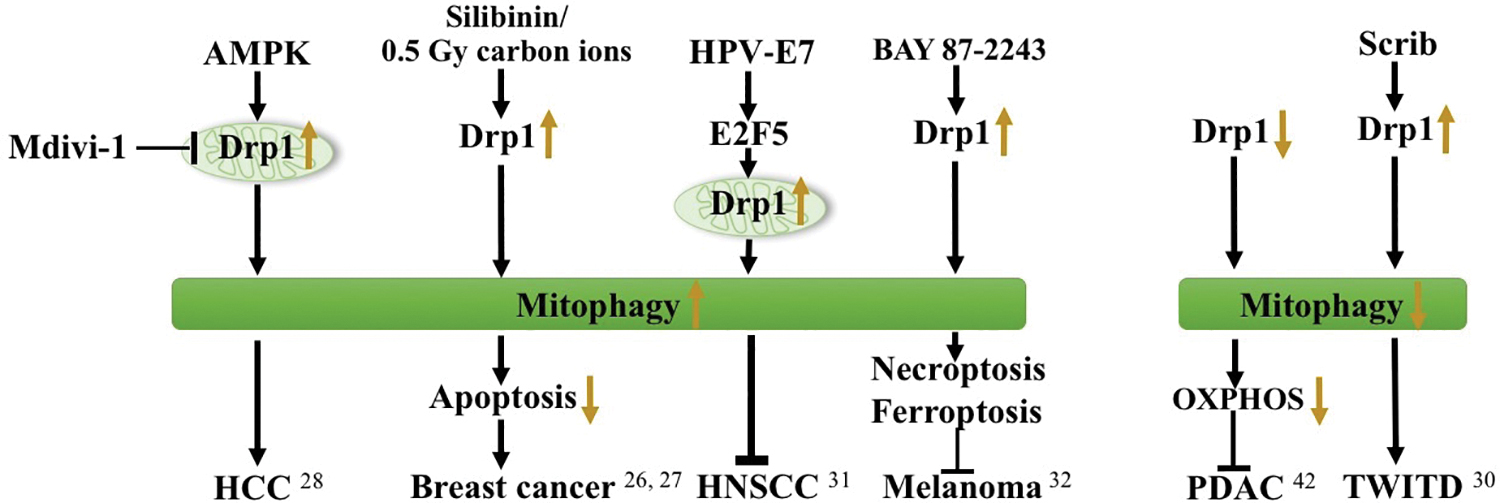
Mitophagy and Drp1 in tumors. Various factors induce or suppress mitophagy through Drp1 in different tumors. Drp1, dynamin-related protein 1. Color images are available online.
Drp1 and apoptosis in tumor cells
Drp1 is closely associated with cell apoptosis. As early as 2003, studies had demonstrated that mitochondria are fragmented during the early stages of apoptosis. Drp1 has been demonstrated to be involved in cell mitochondrial apoptosis. Apoptosis-related protein influences the expression of Drp1 and the morphology of mitochondria. 33 The process of apoptosis is accompanied by depolarization of mitochondria, and the expression of mitochondrial proteins, that is, cytochrome C, to activate caspases and induce mitochondrial apoptosis. Drp1 is primarily involved in the depolarization of the mitochondrial outer membrane during apoptosis. In colon cancer cells, Drp1 knockdown results in the disruption of mitochondrial membrane integrity, a decrease in mitochondrial membrane potential, and an increase in cytochrome C release, resulting in apoptosis. 14 Irradiation of MCF-7 and MDA-MB-231 cells with carbon ions at 3 Gy induces an increase in Drp1 expression levels resulting in apoptosis. 27
Salt-inducible kinase 2 (SIK2) promotes mitochondrial fission through the phosphorylation of Drp1 at Ser616 site. This subsequently results in the inhibition of mitochondrial OXPHOS and cell apoptosis in ovarian cancer cells. 34 Melittin has been demonstrated to have anticancer effects on 4T1 breast cancer cells. The mechanism was through the upregulation of Drp1 and Mfn1 mRNA expression levels to induce cell apoptosis. 35 [Cu (ttpy-tpp)Br2] Br (CTB) has also been demonstrated to strongly suppress the growth of liver cancer xenografts by increasing mitochondrial Drp1 levels and triggering p53-dependent apoptosis. 36 Hu et al. demonstrated that isorhamnetin (IH) mediated p-Drp1 Ser616 signaling, mitochondrial fission, and apoptosis in the triple-negative breast cancer (TNBC) and xenograft mouse models but not in estrogen-dependent breast cancer cells. 37 Additionally, Withaferin A (WA) was found to inhibit breast cancer development by inhibiting Drp1 expression to induce apoptosis. 38 Mammalian Ste20-like kinase 1 (Mst1) overexpression was shown to induce mitochondria-dependent cell apoptosis through DRP1-related mitochondrial fission in squamous cell carcinoma of the head and neck (SCCHN). 39 Large tumor suppressor (LATS) 2 overexpression was found to upregulate DRP1 expression through the Wnt/β-catenin pathway, which subsequently induced HCC apoptosis. 40 PTEN overexpression resulted in the inactivation of the AKT pathway, which enhanced Drp1-mediated mitochondrial fission to induce apoptosis in glioblastomas. 41 Xie et al. demonstrated that SMDT1 could induce the phosphorylation and translocation of Drp1 to the mitochondria to induce apoptosis in PDAC. 42 These previous findings suggest that Drp1 regulates apoptosis and plays an important role in tumorigenesis (Fig. 2).
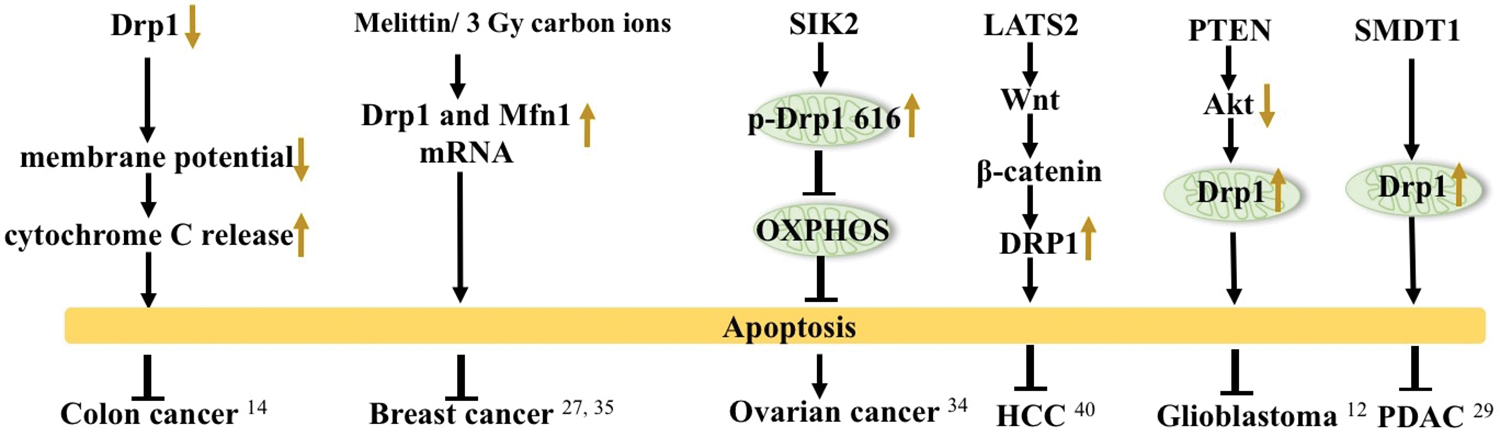
Apoptosis and Drp1 in tumors. Overexpression or reduction of Drp1 can induce apoptosis in cancers. Color images are available online.
Drp1 and tumor stemness
Mitochondria play an important role in tumor stemness, and hence could be a potential therapeutic target for tumor stem cells. 43 Recent studies have demonstrated that mitochondrial fission increases during cell stemness reprogramming. ERK-induced p-Drp1 Ser616 has been demonstrated to play an important role in cell reprogramming. 44 Inhibiting Drp1 has been shown to reduce the reprogramming of mouse fibroblasts, suggesting that Drp1 and mitochondrial fission plays an important role in the maintenance of stemness. 45 Xie et al. demonstrated that the active form of Drp1 was significantly higher in glioma stem cells compared with noncancer stem cells and was associated with increased fragmented mitochondria. The 5-year survival rate of glioma patients with low expression of p-Drp1 Ser616 was significantly higher compared with patients with normal expression levels. 46 Shen et al. found that increased levels of fragmented mitochondria played an important role in maintaining hypoxic phosphorylation levels and tumor stemness in nasopharyngeal carcinoma stem cells. 47 Zhou et al. demonstrated that cyclooxygenase (COX)-2 translocating into the mitochondria could induce mitochondrial p53 translocation, activate Drp1, increase p-Drp1 Ser616 levels, and stemness in nasopharyngeal carcinomas. Inhibition of COX-2 in the mitochondria significantly inhibited the activation of Drp1 and reduced the stemness of nasopharyngeal carcinomas. 48 Drp1-dependent mitochondrial fission is important for maintaining the stemness of mammary epithelial stem-like cells. 15 The mechanism of how Drp1-mediated mitochondrial fission promotes stem maintenance is unclear. In embryonic stem (ES) cells, mitochondrial fragmentation is significantly higher. Downregulation of growth factors leads to decreased Drp1 levels in ES cells, resulting in impaired pluripotency and increased apoptosis. 49 Recent studies have shown that Drp1 mediates mitochondrial fission as well as the asymmetric distribution of mitochondria during stem cell division. Interestingly, asymmetric distribution depends on mitochondrial quality. Aging mitochondria give rise to more differentiated daughter cells, whereas healthy mitochondria are retained in resulting stem cells after cytokinesis. Asymmetric separation of mitochondria helps maintain a homogenous and healthy stem cell population and is one of the mechanisms by which Drp1 maintains stemness. 50 In lymphocytic leukemia cells, ERK1/2 triggers Drp1-dependent mitochondrial fission to reduce ROS and enhance glycolysis. This protects cells from chemotherapeutic drugs. Constitutively active K-Ras mutations activate the ERK signaling pathway to promote tumor transformation and attain stem cell-like characteristics. 51 Additional studies have demonstrated that Parkin disrupts the stability of Drp1, and its loss upregulates Drp1, which contributes to stem cell maintenance. 52 Tumor stemness is an important part of Drp1-mediated mitochondrial regulation of tumor characteristics. Additional studies are necessary to identify specific targets of tumor stemness.
Drp1 and its role in tumor cell invasion and migration
Drp1 is involved in the regulation of invasion and migration of various tumor cells. Drp1 inhibition can reduce tissue infiltration and metastasis of tumor cells. The migration and invasion of tumor cells rely on the formation of pseudopods. The formation of pseudopods requires increased ATP production. Mitochondria, as the main organelle for ATP production, could affect the migration and invasion ability of tumor cells by affecting pseudopod formation. 11,53,54 In breast cancer cells, inhibition of Drp1 reduces mitochondrial fission and pseudopodia formation. This results in reduced cancer cell infiltration and invasion, and reduced tumor malignancy. 11 Epstein-Barr virus latent membrane protein LMP2A can increase the expression of Drp1 in gastric cancer and breast cancer cells. This results in increased metastasis and invasion and is associated with the activation of the Notch pathway. 55 Genetic and pharmacologic blocking of Drp1 activity has been shown to affect the migration/invasiveness of thyroid cancer cells and the characteristics of tumor malignancies. 56 Hatch et al. observed that actin interacts directly with Drp1, 57 and Ji et al. demonstrated that DRP1 regulates the migration and invasion of tumor cells through actin filaments. 58 In addition, Yin et al. demonstrated that downregulation of DRP1 could inhibit RhoA/ROCK1 signaling in human glioma cells to reduce cancer cell skeletal remodeling to prevent invasion and metastasis. 59 This suggests that Drp1-mediated mitochondrial fission promotes tumor invasion and metastasis. However, its specific mechanism remains to be deciphered (Table 1).
Drp1 and tumor metabolic reprogramming
Cells need a set of metabolic modes that are different from normal cells in the process of malignant transformation to quickly and efficiently provide the energy and raw materials needed for biosynthesis. This process is called metabolic reprogramming. Metabolic reprogramming mainly involves glycolysis, tricarboxylic acid cycle, OXPHOS, amino acid metabolism, fatty acid metabolism, and nucleic acid metabolism. Aerobic glycolysis is the first metabolic reprogramming process discovered by Otto Warburg. 60 Drp1 is closely related to metabolic reprogramming of cancer cells, especially glycolysis.
Prieto et al. suggest that mitochondrial fission regulated by an Erk-Drp1 axis constitutes an early and necessary step in the metabolic reprogramming process to pluripotent stem (iPS) cells. 44,61 Lee found that Androgen induced Drp1 upregulation is required for metabolic reprogramming of Prostate Cancer (PC) through VDAC-MPC2 complex pathway. 62 Cruz et al. found that changes in gene DNM1L and early Warburg-like metabolic changes may provide a favorable microenvironment for CRC. 63 SIK2 promotes reprogramming of glucose metabolism through PI3K/AKT/HIF-1α pathway in ovarian cancer. 34 Glucose starvation (GS) remarkably sensitized MDA-MB-231 to apoptosis and decreased its migratory potential by increasing mitochondrial ROS and upregulating Prohibitin 1 (PHB1), leading to its dissociation from Drp1, and triggering of the apoptosis cascade. 64 Li et al. observed that tumor cells under an energy stress condition exhibited a shift from glycolysis toward OXPHOS to survive by phosphorylating Drp1 at S637 site through NAD+-dependent SIRT1 activation. 65 During hypoxia, Kuo et al. found the existence of mtDNA elevated levels of HIF-1α, causing an increased expression of Drp1 in eliciting metabolic reprogramming, and mitochondrial biogenesis in neuroblastoma. 66 However, Dai et al. shows that mdivi-1 treatment impaired cell proliferation through limiting oxidative metabolism, rather than glycolysis, which was independent of Drp1 inhibition or mitochondrial fusion induction in H460, A549, and HCT116 cell lines. 67
Future Directions
Numerous studies have demonstrated that Drp1 promotes tumorigenesis. Drp1 has been associated with the proliferation, invasion, and metastasis of various cancers.
At present, studies on the regulatory mechanism of Drp1 have garnered increased interest. However, only mitochondrial fission Inhibitor, Mdivi-1, have been pharmacologically demonstrated to inhibit Drp1. Encouragingly, recent studies have discovered additional inhibitors that could target Drp1 to inhibit cancer development and progression.
Drp1-targeting drugs for cancer treatment
Preclinical studies have demonstrated that resveratrol 68 and other small molecules from plants could target Drp1 and have antitumor effects. These include P11069 and melatonin. 69 Blocking Drp1 mitochondrial translocation has been demonstrated to significantly sensitize breast cancer cells to MLN4924. 70 An ellipticine compound, Drpitor1, was identified as a Drp1 inhibitor by silico screening. It could reduce proliferation and induce apoptosis in five cancer cell lines (A549, SK-MES-1, SK-LU-1, SW 900, and MCF7). 71
In addition, Silibinin has been demonstrated to induce apoptosis and inhibit invasion and migration of MCF-7 and MDA-MB-231 cells by decreasing Drp1 expression levels. 26 Except for cisplatin, doxorubicin, and WA, 38 melittin has been demonstrated to have anticancer inhibitory effects on 4T1 cell lines and could be a good candidate for further research on breast cancer treatment. 35 Metformin has been shown to have antitumor activity by specifically targeting Drp1 to inhibit the glycolytic pathway of breast cancer stem cells. 72 Oral leflunomide, an FDA-approved arthritis drug, was demonstrated to reduce Drp1 expression and OXPHOS in tumors and was repurposed as a chemotherapeutic for the treatment of pancreatic cancer. 29
Drp1-targeting drugs in combination with chemotherapy
Chemotherapy drugs have been demonstrated to induce drug and apoptosis resistance in tumor cells. 73 Chemotherapeutic drugs have been shown to increase Drp1 expression levels in tumor cells, which consequently increases mitochondrial fission resulting in antiapoptotic effects and reducing drug sensitivity. Several studies have demonstrated that specific targeting of Drp1 could enhance apoptosis and sensitivity of tumor cells to chemotherapy drugs. 74 In ovarian cancer (OVCA), inhibition of Drp1-dependent mitochondrial fission significantly reduces cisplatin-induced apoptosis, resulting in decreased chemosensitivity. 75 Zhou et al. demonstrated that the natural phytochemical, Liensinine, could reduce lysosomal binding to mitochondria and inhibit late mitophagy. Liensinine has been demonstrated to significantly increase the sensitivity of breast cancer cells to chemotherapy drugs. 76 In addition, cisplatin has been demonstrated to significantly induce Drp1 expression leading to increased mitophagy. ABT737 could significantly increase tumor sensitivity to cisplatin for the treatment of cholangiocarcinomas 77 and OVCA. 78 The cellular mechanism involved the ability of ABT737 to inhibit cisplatin-induced Drp1 expression and mitophagy. Chloroquine has been shown to inhibit autophagy/mitophagy in TNBC cells by selectively enhancing IH-induced mitochondrial fission and apoptosis, but not in estrogen-dependent breast cancer cells. These findings suggest that IH could potentially be a novel chemotherapeutic drug. The combination of IH with classic autophagy/mitophagy inhibitors could represent a novel therapeutic strategy for the treatment of TNBC. 37 In addition, E2F5-mimetic peptides have been demonstrated to activate and induce Drp1 mitochondrial localization to enhance ceramide-mediated mitophagy leading to tumor suppression in HPV-negative HNSCC-derived xenograft tumor SCID mouse models in response to cisplatin 31 (Table 2).
Interventions Targeting Dynamin-Related Protein 1 and Coadministration with Chemotherapy
IH, isorhamnetin; HNSCC, head and neck squamous cell carcinoma; OVCA, ovarian cancer; TNBC, triple-negative breast cancer.
Although there is not yet Drp1-targeting drugs in combination with chemotherapy being used in clinical, the expression levels of Drp1 can be used as an early diagnostic marker and an indicator of tumor grade. For example, the expression levels of Drp1 were higher in cutaneous squamous cell carcinoma than in normal epidermis, and correlated positively with the advanced clinical stages. 79 The clinical data also show a strong inverse correlation between phosphorylation of Drp1 on Ser616 and poor survival of patients with glioblastoma, 46 whereas overexpression of Drp1 on Ser637 predicted poor prognosis of HCC patients. 67 In patients with lung cancer, increased Drp1 expression predicts higher rate of recurrence (3.4 times) and cisplatin resistance. 80 The information listed above provides the rationale for the use of Drp1 inhibitors for the clinical management of cancers. Additional studies are required to understand the mechanism of action of Drp1 and identify drugs and small molecules to inhibit Drp1.
Footnotes
Disclaimer
The work described has not been submitted elsewhere for publication, in whole or in part. The final version of this article has been approved by all authors listed in this review.
Authors' Contributions
P.H., X.R., and Y.M. drafted the article and critically revised it. X.Q. and P.H. drew the figures, and designed the tables. P.H. substantially contributed to conception and design. All the authors gave final approval and agreed to be accountable for all aspects of work.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.