
Abstract
Background:
Renal cell carcinoma (RCC) is one of the most common and malignant tumors in the urinary system. The aim of this research was to investigate the mechanism and clinical significance of miR-4461 in the RCC progression.
Materials and Methods:
Twenty-eight (28) paired RCC tissue samples and adjacent nontumor tissue samples, as well as RCC cell lines were used to measure the expression of miR-4461 and protein phosphatase 1 regulatory subunit 3C (PPP1R3C) transcript by real-time quantitative PCR. The target relationship between miR-4461 and PPP1R3C was predicted by TargetScan and further verified by dual-luciferase reporter gene assay and RNA pull-down assay. Cell Counting Kit-8 (CCK-8) assay and BrdU ELISA assay were performed to measure RCC cell viability and proliferation. In addition, caspase-3 activity assay and cell adhesion assay were implemented to measure RCC cell apoptosis and adhesion.
Results:
MiR-4461 was lowly expressed both in RCC tissues and cells, while upregulated PPP1R3C was tested in RCC tissues and cells. In addition, miR-4461 was validated to directly target PPP1R3C, thereby negatively regulating PPP1R3C. Particularly, miR-4461 exerted a clear inhibitory effect on the malignant phenotypes of RCC cells by binding and inhibiting PPP1R3C.
Conclusion:
MiR-4461, which served as a tumor suppressor, inhibited RCC progression by targeting and downregulating PPP1R3C.
Introduction
Renal cell carcinoma (RCC), one of the most common urogenital malignancies, stems from renal tubular epithelial cells and accounts for 2%–3% of all malignancies in adults. 1,2 RCC accounts for 80%–90% of all primary renal malignancies. 3
At present, although radical or partial nephrectomy has become an effective treatment for RCC, the overall survival rate is still not optimistic 4,5 because RCC is characterized by early absence of obvious symptoms, high risk of metastasis, and easy recurrence after surgery, as well as poor response to radiotherapy and chemotherapy. 5 –8 For advanced RCC, metastasis, and recurrence, there is currently no effective treatment. 9 –11 Therefore, the authors have great motivation and responsibility to research the pathogenesis and metastasis molecular mechanism of RCC, which would provide potential targets for early diagnosis, therapeutic intervention, and prognostic indicators.
MiRNA, one of the key members of the noncoding RNA family, currently the most studied noncoding RNA, are typically 19–24 nucleotides in length. 12,13 Countless evidence reveals the involvement of miRNAs in the development of cancer, in which by specifically combining with the downstream target gene 3′-UTRs, miRNAs exert their negative regulatory roles of gene expression. 14,15 In RCC, numerous miRNAs had been extensively studied, some presenting as cancer promoter, such as miR-183, 16 miR-106b, 17 and miR-142-5p, 18 while others presenting as suppressor, such as miR-30b-5p, 19 miR-28-5p, 20 miR-15a, 21 and miR-186. 22
MiR-4461, a mitochondrial related miRNA, 23 was reported to be the tumor suppressor in colorectal cancer. 24 Although miR-4461 has not been studied in RCC so far, according to previous research, miR-4461 may play a role in suppressing tumors. Therefore, it is necessary and significant for us to investigate the mechanism of miR-4461 in RCC.
Protein phosphatase 1 regulatory subunit 3C, a glycogen-targeting subunit for PP1, is able to regulate the activity of protein phosphatase 1 (PP1). 25 By activating glycogen synthase, PPP1R3C promotes glycogen biosynthesis, and by reducing glycogen phosphorylase activity, PPP1R3C restricts glycogen decomposition. Therefore, PPP1R3C can induce glycogen accumulation and storage. 25,26 During tumorigenesis, it will be accompanied by a variety of metabolic abnormalities, especially abnormal glucose metabolism, which provide energy for cancer cells to survive and proliferate. 27 –29
Mounting researches have uncovered that reprogramming glucose metabolism appeared in kinds of cancers, such as hepatocellular carcinoma, 30 breast cancer, 31 pancreatic cancer, 32 and bladder cancer, 33 and high glucose metabolism had been recognized as one of the hallmarks of cancer. 34 As an enzyme that regulated glucose metabolism, PPP1R3C was involved in the development of cancer by mediating glycogen accumulation or DNA methylation. 35,36 PPP1R3C had been shown to be abnormally expressed in a few of cancers, thus affecting the biological behavior of cancers, such as colon cancer 25 and cervical cancer. 37 However, whether PPP1R3C is involved in the development of RCC and its mechanism in RCC are worth exploring.
This study was designed to discuss the clinical and biological significance of miR-4461 in RCC, as well as the regulatory mechanism of miR-4461/PPP1R3C axis in the progression of RCC. In this study, the authors uncovered that downregulated miR-4461 and upregulated PPP1R3C appeared in RCC tissues and cells. Notably, they also confirmed that miR-4461 could downregulate the expression of PPP1R3C by binding to PPP1R3C as an upstream regulating miRNA, thus inhibiting the malignant phenotypes of RCC cells. Therefore, through this study, miR-4461 and PPP1R3C might become promising biomarkers and drug targets for clinic diagnosis and treatment of RCC.
Materials and methods
Microarray analysis
GSE781, GSE68417, and GSE117890 downloaded from GEO DataSets were the mRNA expression profiles. GSE781 included 9 RCC tissues and 8 adjacent nontumor tissues, GSE68417 included 29 RCC tissues and 14 adjacent nontumor tissues, and GSE117890 included 5 RCC tissues and 6 normal tissues. The upregulated differentially expressed genes (DEGs) were screened using Limma 3.26.8 with adj.p-value <0.05 and |logFC| ≥ 1.5. GSE95384 was the miRNA expression profile, including eight RCC tissues and eight normal tissues. The downregulated differentially expressed miRNAs were confirmed by Limma 3.26.8 with adj.p-value <0.05 and logFC <−1. Venny 2.1.0 was performed to select out the overlapping DEGs and miRNAs.
Clinical tissue samples
Twenty-eight (28) pairs of clinical samples, including RCC tumor tissues and adjacent nontumor tissues, were acquired from patients who were diagnosed with RCC in Wuhan Third Hospital. All RCC patients were not treated with any drug or other therapy before surgical excision. After surgical excision, the tissue samples were labeled and frozen for later analysis. The informed consent had been signed by all patients with RCC and the clinical research had been approved by the ethics committee of Wuhan Third Hospital. Clinical characteristics of 28 cases of RCC patients are displayed in Table 1.
Clinical Characteristics of 28 Cases of Patients with Renal Cell Carcinoma
Cell lines and cell culture
Five RCC cell lines, containing SW839 (Cat. No.: HTB-49), Caki-1 (Cat. No.: HTB-46), A498 (Cat. No.: HTB-44), 769-P (Cat. No.: CRL-1933), 786-O (Cat. No.: CRL-1932), as well as human normal kidney cell line HK-2 (Cat. No.: CRL-2190) were bought from American Type Culture Collection (ATCC, USA). Another RCC line, OSRC-2 (Cat. No.: BNCC100109), was bought from BeNa Culture Collection (BNCC, China). All these cells, except for HK-2 and A498 cell lines, were cultured in RPMI 1640 medium (Hyclone, USA). HK-2 and A498 cell lines were cultured in DMEM (Gibco, USA). In the medium, fetal bovine serum (Gibco, China) was added at the final concentration of 10% and streptomycin/penicillin (Invitrogen, USA) was added at 100 U/mL. All these cell lines were cultured under the same condition, which was 5% CO2 incubator at 37°C.
Cell transfection
MiR-4461 mimic, miR-4461 inhibitor, and miRNA negative control (NC), as well as small interfering RNA of PPP1R3C (si-PPP1R3C) in this study, were designed and bought from FulenGen (China). The pGL4 luciferase reporter vectors (Promega, USA) were used to construct dual-luciferase reporter plasmid vectors containing PPP1R3C 3′-UTR sequences or the corresponding mutation sequences. The target cells (SW839 and OSRC-2) were transfected using Lipofectamine RNAiMax (Life Technologies, USA) according to the product manual. Following 48 h of transfection, the cells were collected for real-time quantitative PCR (RT-qPCR) experiments to verify the efficiency of transfection. After successful transfection, the transfected cells were used for subsequent studies.
Real-time quantitative PCR
First, tissue samples or cultured cells were lysed by the TRizol reagent (Invitrogen, USA) to isolate and purify total RNA following the product manual. The spectrophotometer (METASH, China) was used to quantify the total RNA. Then, according to the product manual, cDNA was synthesized with 1g total RNA using SuperScript™ IV One-Step RT-PCR System (Cat. No.: 12594025; Invitrogen, USA). Subsequently, qPCR was implemented at least three times using SYBR™ Select Master Mix (Cat. No.: 4472903; Applied Biosystems, USA) on ABI 7500 Fast System (Applied Biosystems). U6 acted as an internal reference to normalize miR-4461, while GADPH acted as an internal reference to normalize PPP1R3C. The relative RNA expression of miR-4461 or PPP1R3C was analyzed by 2−▵▵Ct calculation. All the sequences of primers are listed in Table 2.
The Primer Sequences for Real-Time Quantitative PCR
Cell viability assay
The Cell Counting Kit-8 (CCK-8) assay evaluated the viability of SW839 and OSRC-2 cells according to the CCK-8 (Cat. No.: CK04-13, Dojindo, Japan). In short, both the transfected SW839 and OSRC-2 cells were planted into a 96-well plate at 5 × 103 cells per well, and placed at 37°C in a 5% CO2 incubator for 0, 24, 48, and 72 h cultivation. A group of cells was taken out every 24 h and 10 μL of CCK-8 solution was added to each well, followed by 2 h of incubation under the same condition. The absorbance at 450 nm was measured on a microplate reader to represent the viability of each group of cells.
Cell proliferation assay
The proliferation ability of SW839 and OSRC-2 cells 48 h post-transfection was evaluated using CytoSelect™ BrdU Cell Proliferation ELISA Kit (Cat. No.: CBA-251; Cell Biolabs, USA). In short, 5 × 104 transfected cells were suspended in 100 μL culture medium as described above, and plated into each well of the 96-well plates for 48-h cultivation in an incubator. Then, the cells were cultured for another 4 h in BrdU-containing medium. Next, the cells were immobilized and the cellular DNA was denatured. After the immune response with anti-BrdU antibody, absorbance at 450 nm was measured on a microplate reader to evaluate the incorporation of BrdU in cells.
Cell apoptosis assay
The apoptosis of SW839 and OSRC-2 cells was evaluated by measuring caspase-3 activity using a caspase-3 colorimetric assay kit (Cat. No.: ab39401; Abcam, USA). In short, 2 × 104 transfected cells were plated into each well of a 96-well plate for 48 h of cultivation in a 5% CO2 incubator at 37°C. Cells were collected and lysed on ice with lysis buffer for 1 h. Then, the supernatant was collected and aliquots of supernatant were mixed with the reaction buffer containing DEVD-p-NA substrate, and incubated in the dark at 37°C for 4 h. The absorbance at 405 nm was measured on a microplate reader for monitoring caspase-3 activity.
Cell adhesion assay
The adhesion of SW839 and OSRC-2 cells was evaluated by MTS reagent (Promega) using 96-well plates precoated with fibronectin (Sigma Aldrich, USA). In short, the plates were blocked with 1% bovine serum albumin for 1 h. Then, ∼2 × 104 transfected cells were plated into each well of the 96-well plates precoated with fibronectin for 1-h incubation at 37°C. After that, the unattached cells were removed by washing with phosphate buffered saline (PBS). Twenty microliters of MTS reagent was added to the adhesion cells in each well, and incubated at 37°C for 2 h. The absorbance at 490 nm was measured on a microplate reader to represent the adhesion ability of each group of cells.
Dual-luciferase reporter gene assay
The PPP1R3C 3′-UTR wild-type (WT) sequences or the corresponding mutation sequences (mutation 1 sequence, mutation 2 sequence, or their co-mutation sequence) were designed and bought from FulenGen (China). Then, the recombinant reporter plasmid vectors were inserted into the pGL4 Luciferase Reporter Vectors (Promega). Next, the recombinant reporter plasmid vectors and miR-4461 mimic were co-transfected into the SW839 and OSRC-2 cells, and then the cells were cultured in an incubator for 48 h. The activity of Renilla luciferase and firefly luciferase was tested using Pierce™ Renilla-Firefly Luciferase Dual Assay Kit (Cat. No. 16186; Thermo Scientific, USA) on a luminometer. Renilla luciferase was set as an internal reference to normalize firefly luciferase.
RNA pull-down assay
The RNA pull-down assay was performed on the SW839 and OSRC-2 cells using streptavidin magnetic beads (Life Technologies) and biotin-coupled miR-4461 probes and random oligo probes as NC group. In short, ∼1 × 107 SW839 and OSRC-2 cells were collected and lysed on ice with lysis buffer. Then, the whole supernatant lysate was collected and incubated with biotin-coupled miR-4461 probes or random oligo probes for 2 h at room temperature. After that, the biotin-coupled miR-4461 complex was pulled down by following incubation with streptavidin magnetic beads for another 4 h at room temperature. Finally, the beads were washed with elution buffer to extract the bound RNAs in the pull-down complex, and the pull-downed RNAs were analyzed by RT-qPCR.
Fluorescein phosphoramidate-siRNA transfection and quantification
Fluorescein phosphoramidate (FAM)-labeled si-PPP1R3C (FAM-si-PPP1R3C), miR-4461 mimic (FAM-miR-4461 mimic), and miR-4461 inhibitor (FAM-miR-4461 inhibitor) were designed and bought from Gene Pharma (Shanghai, China). For transfection and quantification, about 1 × 105 SW839 and OSRC-2 cells were seeded in coverslips, which were flatted on a six-well plate. After 24 h culture, the cell transfection of the FAM-si-PPP1R3C, FAM-miR-4461 mimic, and FAM-miR-4461 inhibitor was performed based on the procedure shown in cell transfection. After further culture for 24 h, the cells were gently washed with PBS and permeated with 0.1% triton. Next, the cells were stained with 4′,6-diamidino-2-phenylindole. Finally, the coverslips with cells were transferred onto microslides and the images were captured by the Flowview 1000 Laser Scanning Confocal Microscope (Olympus, Japan). The fluorescent cells and the fluorescence intensity were measured.
Statistical analysis
The authors employed the SPSS 20.0 software (SPSS, USA) to conduct data analyses in this article. And the results of the three independent experiments were expressed as mean ± standard deviation. The difference analysis between the two groups was evaluated by Student's t-test, while the difference analysis among the multiple groups was evaluated by analysis of variance (ANOVA). p < 0.05 was considered statistically significant.
Results
The identification of key gene and miRNA involving RCC
With adj.p-value <0.05 and |logFC| ≥ 1.5, 1107, 867, and 49 DEGs were selected from GSE781, GSE68417, and GSE117890, respectively (Fig. 1A). By Venny 2.1.0 analysis, a total of 6 overlapping DEGs were identified. Then, the authors detected the expression of 6 DEGs in normal and tumor tissues from RCC patients using RT-qPCR. The results showed that the expression of 6 DEGs mRNA was elevated in tumor tissues, especially PPP1R3C, whose expression increased by more than twofold in tumor tissues (Fig. 1B–G). Therefore, PPP1R3C was confirmed as the key mRNA to further explore its effect on RCC. Then, TargetScan predicted 403 miRNAs that could bind to the PPP1R3C 3′-UTR, as well as seven downregulated miRNAs were selected from GSE95383 with adj.p-value <0.05 and logFC <−1 (Fig. 1H). Finally, hsa-miR-4461 was overlapped from GSE95348 and TargetScan by Venny 2.1.0 analysis so that miR-4461 was confirmed to the key miRNA for further investigation.

PPP1R3C and miR-4461 were confirmed the key gene and miRNA to investigate in RCC.
MiR-4461 was lowly expressed in RCC tissues and cells
To investigate whether miR-4461 expression was different in RCC tissues and noncarcinoma tissues, RT-qPCR was applied to assess miR-4461 expression in 28 RCC tissue samples and adjacent noncarcinoma tissue samples. As the data showed in Figure 2A, miR-4461 expression in RCC tissues was reduced by ∼54% in contrast to noncarcinoma tissues. Then, the authors detected the expression of miR-4461 by RT-qPCR in various human RCC cell lines and human normal kidney cell line HK-2. As expected, low expression of miR-4461 was detected in six human RCC cell lines (SW839, A498, OSRC-2, 769-P, 786-O, and Caki-1) compared to HK-2 cell line. In particular, miR-4461 expression was found to be lowest in SW839 and OSRC-2 cells (Fig. 2B). Therefore, these two cell lines (SW839 and OSRC-2) were screened for subsequent in vitro functional experiments. Together, the data above revealed that the change of miR-4461 was inextricably bound up with RCC, showing prominently downregulated in RCC.

miR-4461 was lowly expressed in RCC tissues and cells.
MiR-4461 suppressed malignant phenotypes of RCC cells
Next, miR-4461 mimic or miR-4461 inhibitor was transfected into SW839 and OSRC-2 cells to better explore the effect of miR-4461 on the biological behavior of RCC cells. As shown by RT-qPCR results, in SW839 and OSRC-2 cells, miR-4461 expression was strongly raised by miR-4461 mimic transfection, while its expression was apparently decreased by miR-4461 inhibitor transfection, in which miR-4461 mimic caused miR-4461 expression to increase by ∼2.73 times, while miR-4461 inhibitor caused miR-4461 expression to decrease by ∼75% (Fig. 3A). The transfection efficiency of miR-4461 mimic, miR-4461 inhibitor and si-PPP1R3C was similar (∼80%) in SW839 and OSRC-2 cells by the detection of fluorochrome-labeling method (Supplementary Fig. S1).

miR-4461 suppressed malignant phenotypes of RCC cells.
Subsequently, CCK-8 assay was used to evaluate whether miR-4461 upregulation or downregulation had changed the viability of SW839 and OSRC-2 cells. The results are shown in Figure 3B; when transfected with miR-4461 mimic in SW839 and OSRC-2 cells, the viability was obviously lower than that in the control group. However, after transfection with miR-4461 inhibitor, the viability was markedly higher than that in the control group. Besides, BrdU ELISA assay was implemented and displayed that miR-4461 mimic resulted in a 28% decrease in absorbance at 450 nm of SW839 and OSRC-2 cells, and miR-4461 inhibitor resulted in a 47% increase in absorbance at 450 nm of SW839 and OSRC-2 cells (Fig. 3C), suggesting that miR-4461 played an inhibitory role in the proliferation of SW839 and OSRC-2 cells. Furthermore, caspase-3 activity was measured in SW839 and OSRC-2 cells to assess cell apoptosis. The results revealed that compared with control group, caspase-3 activity in SW839 and OSRC-2 cells was upregulated by 3.22 times by miR-4461 mimic transfection, while caspase-3 activity in SW839 and OSRC-2 cells was reduced by 68% by miR-4461 inhibitor transfection, indicating that miR-4461 mimic could accelerate RCC cell apoptosis, while miR-4461 inhibitor could inhibit RCC cell apoptosis (Fig. 3D). Finally, alterations in adhesive capacity of SW839 and OSRC-2 cells were measured using a cell adhesion assay. Results as displayed in Figure 3E, compared with control group, SW839 and OSRC-2 cells exhibited a 67% reduction in adhesive capacity after transfection with miR-4461 mimic. By contrast, the adhesive capacity of the cells was increased by 56% after transfection with miR-4461 inhibitor. In summary, these data illustrated that miR-4461 upregulation prominently accelerated cell apoptosis, but alleviated viability, proliferative capacity, and adhesive capacity of RCC cells, while miR-4461 inhibitor had an inverse effect.
PPP1R3C acted as a target gene of miR-4461
To unravel the underlying mechanism by which miR-4461 exerted its functional effects in RCC cells, TargetScan was employed to forecast possible binding sites between PPP1R3C 3′-UTR and miR-4461. As shown in Figure 4A, the 3′-UTR of PPP1R3C contained two potential specific binding sites for miR-4461.
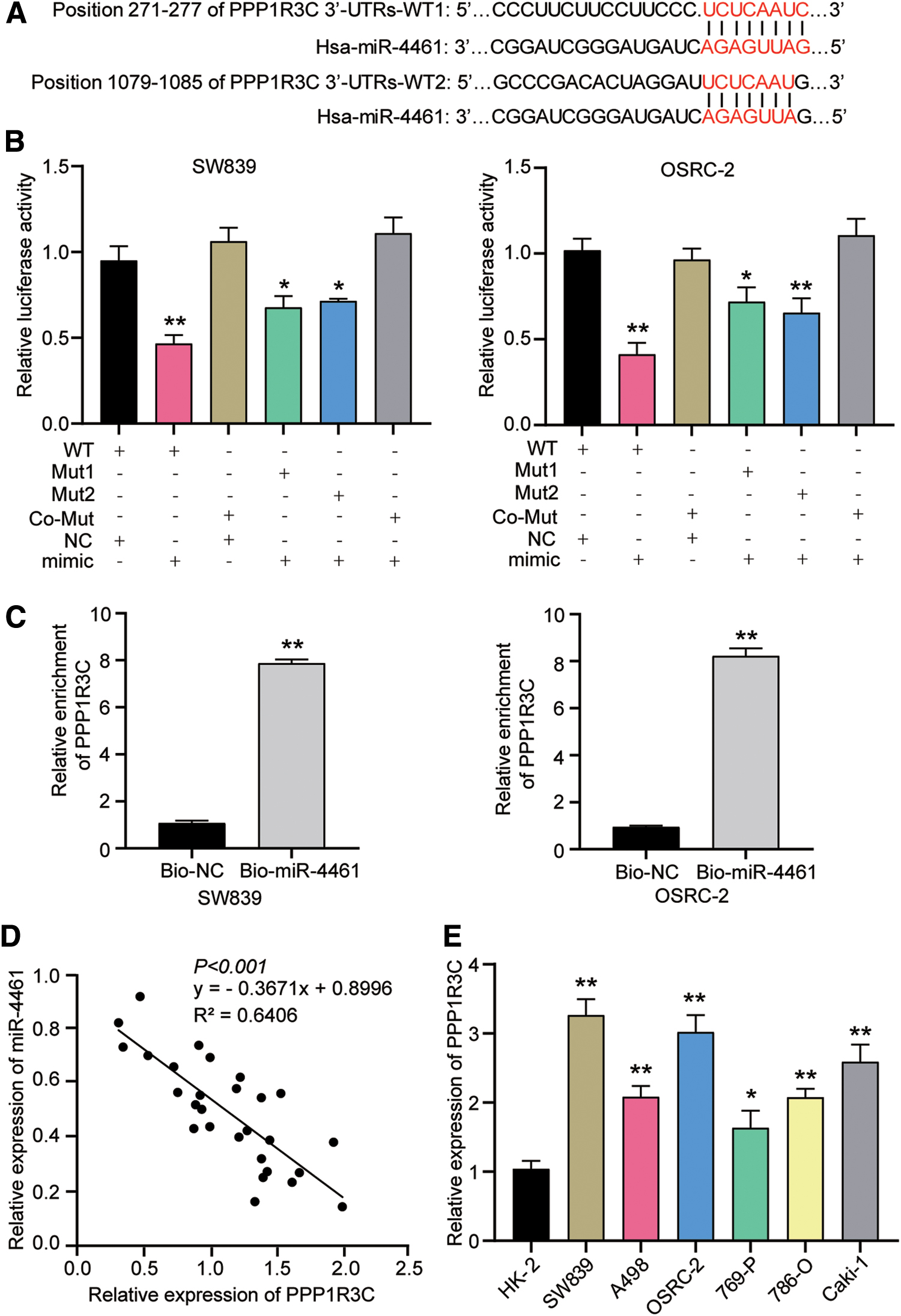
PPP1R3C acted as the target gene of miR-4461.
To verify whether these two sites in the PPP1R3C 3′-UTR region were miR-4461-specific binding sites, the authors conducted luciferase reporter vectors containing PPP1R3C 3′-UTR WT sequences or the corresponding mutation sequences (mutation 1 sequence, mutation 2 sequence, or their co-mutation sequence). As the results from dual-luciferase reporter assay, after transfection with miR-4461 mimic, the luciferase activity of reporter genes with a single-site mutation, including mutation 1 (Mut1) and mutation 2 (Mut2), was ∼1.6 times higher compared with WT reporter genes. However, co-mutated (Co-Mut) reporter gene with two mutation binding sites had a higher luciferase activity than other reporter genes, and its activity was 2.5 times compared with the WT reporter gene (Fig. 4B). This indicated that by complementary pairing with these two sites, miR-4461 could directly target PPP1R3C.
Furthermore, SW839 and OSRC-2 cells were subjected to RNA pull-down assay to further confirm this interplay between PPP1R3C and miR-4461. The outcomes as illustrated in Figure 4C, the enrichment of PPP1R3C in the pull-down complex was much higher in biotin-miR-4461 group than in biotin-NC group. That was to say, PPP1R3C could be plentifully gathered by miR-4461, which suggested that PPP1R3C could be combined with miR-4461. Besides, by Pearson correlation analysis, the authors found that there was a negative correlation between the expression level of PPP1R3C and miR-4461 in RCC tissues (Fig. 4D). To further understand the relationship between the two, they detected the expression of PPP1R3C in the same set of cell lines in Figure 2B by RT-qPCR analysis. As expected in Figure 4E, the higher expression of PPP1R3C was detected in the six RCC cell lines than that in the normal cell line HK-2.
In summary, these data in this section showed that PPP1R3C acted as the target gene of miR-4461, and its expression level was negatively correlated with the expression level of miR-4461.
MiR-4461 suppressed malignant phenotypes of RCC in vitro by inhibiting PPP1R3C
To uncover the mechanism of PPP1R3C in miR-4461-mediated inhibition of RCC progression, the siRNA targeting PPP1R3C was designed to downregulate the expression of PPP1R3C, and exogenous miR-4461 inhibitor was used to downregulate the expression of miR-4461; then, si-PPP1R3C, miR-4461 inhibitor, or both was transfected into SW839 and OSRC-2 cells to test the expression of PPP1R3C mRNA and miR-4461. As can be seen from Figure 5A, whether in SW839 or OSRC-2 cells, miR-4461 inhibitor decreased the expression of miR-4461 and increased the expression of PPP1R3C mRNA, and compared with control group, the expression of miR-4461 decreased by 69% and the expression of PPP1R3C mRNA increased by two times. However, si-PPP1R3C decreased the expression of PPP1R3C mRNA by 62%, but had no effect on the miR-4461 expression. The transfection efficiency of miR-4461 mimic, miR-4461 inhibitor, and PPP1R3C siRNA can also be seen by the fluorochrome-labeling method (Supplementary Fig. S1). Next, to further evaluate whether miR-4461 inhibited the RCC progression by regulating PPP1R3C, si-PPP1R3C was transfected to knockdown PPP1R3C and miR-4461 inhibitor was transfected to upregulate miR-4461 for further function experiments to explore phenotype changes of RCC cells. CCK-8 assay revealed that compared with the control group, the viability of SW839 and OSRC-2 cells was weaker in the si-PPP1R3C group, while in the miR-4461 inhibitor+si-PPP1R3C group, there was no significant change in the viability of SW839 and OSRC-2 cells (Fig. 5B). Besides, the BrdU ELISA assay exhibited that SW839 and OSRC-2 cells had a roughly 40% decrease on cell proliferation by si-PPP1R3C transfection. However, there was virtually no change in cell proliferation capacity of SW839 and OSRC-2 cells by miR-4461 inhibitor and si-PPP1R3C co-transfection (Fig. 5C). In addition, the authors tested caspase-3 activity to evaluate RCC cell apoptosis capacity. As could be seen from the Figure 5D, both in SW839 and OSRC-2 cells, PPP1R3C knockdown led to an approximately threefold increase in caspase-3 activity. More important, PPP1R3C knockdown could partially weaken the inhibiting effect of miR-4461 inhibitor on cell apoptosis when co-transfected with miR-4461 inhibitor and si-PPP1R3C, and compared with the control group, there was no significant change. Finally, cell adhesion assay was implemented in SW839 and OSRC-2 cells and revealed that compared with the control group, the adhesive capacity in si-PPP1R3C group was reduced by ∼57%. However, in the miR-4461 inhibitor+si-PPP1R3C group, the promotion effect of miR-4461 inhibitor on cell adhesion was partially impaired by PPP1R3C knockdown, and in contrast to the control group, the adhesive capacity did not change significantly (Fig. 5E). In summary, the above data revealed that miR-4461 suppressed malignant phenotypes of RCC cells by inhibiting PPP1R3C.
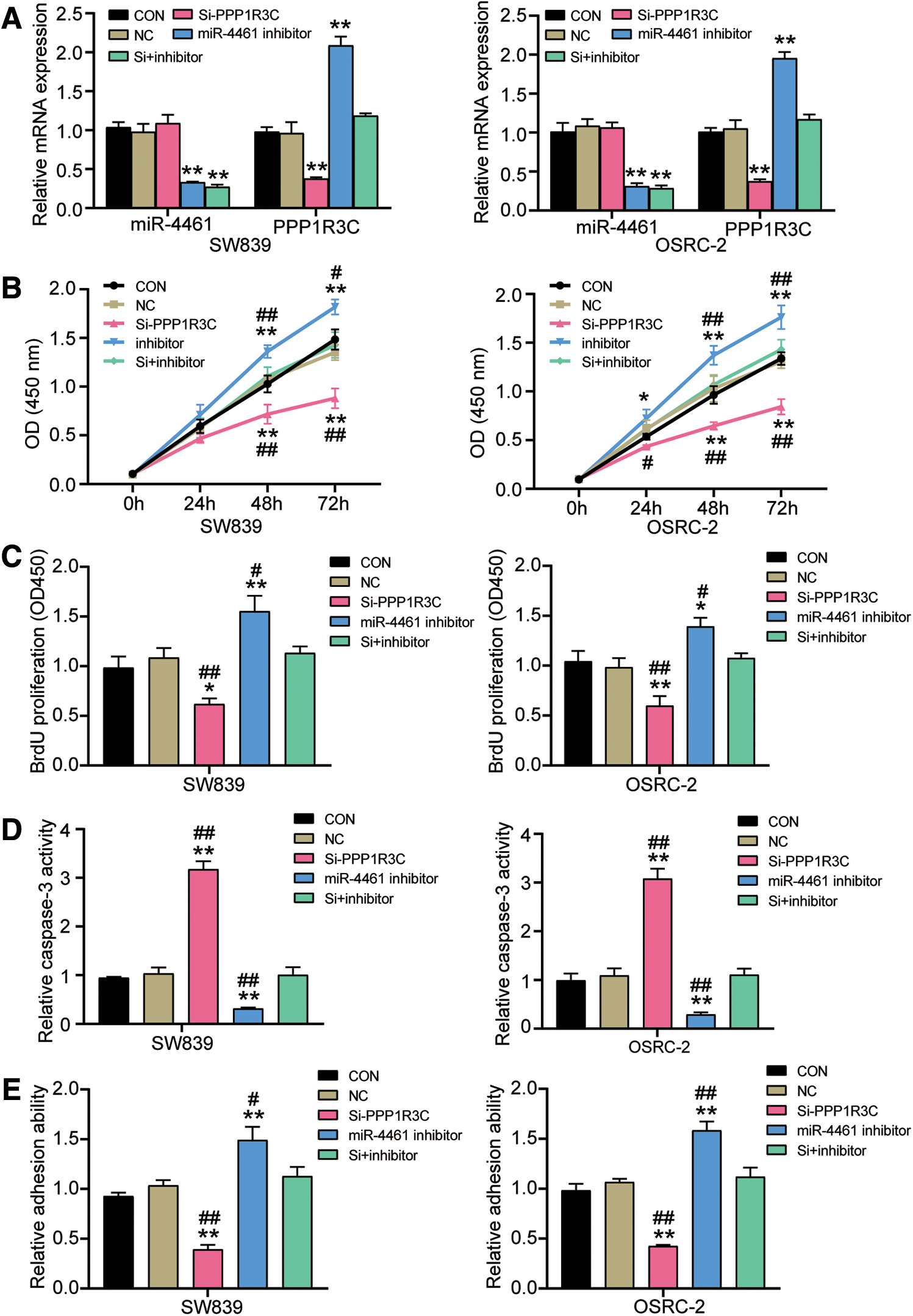
MiR-4461 suppressed malignant phenotypes of RCC in vitro by inhibiting PPP1R3C.
Discussion
RCC, the most deadly and common type of renal carcinoma in adult, is most likely to occur in people older than 40 years, and affect people older than 70 years. 11 In this study, a decreased expression of miR-4461 was observed not only in RCC tissues but also in RCC cell lines. Furthermore, the authors found that miR-4461 was an upstream modulator of PPP1R3C and could negatively regulate the expression of PPP1R3C in RCC cells. Furthermore, miR-4461 was proved to have a strong inhibitory effect on the malignant phenotypes of RCC cells, and this suppression was achieved by targeting and inhibiting PPP1R3C. These new data might provide a new insight into the mechanism of the occurrence and development of RCC. miR-4461 and PPP1R3C might serve as new biomarkers and treatment targets for RCC.
Dysregulated miRNA expression has been shown in carcinogenesis. 38 Coincidently, numerous miRNAs have been identified to function as either carcinogenic or tumor-suppressive miRNAs in the occurrence of RCC. 39 –42 For instance, miR-224, miR-193a-3p, and miR-543 emerged to be upregulated in RCC, 1,43 suggesting its tumor promoter role in RCC. However, other miRNAs like miR-193b and miR-106a-5p appeared to be downregulated in RCC and involved in the progression of RCC as suppressors. 44,45 Although miR-4461 has not been studied in RCC, its regulating function has been explored in several other cancer types. For example, 106 miRNAs were found significantly dysregulated after the CRT-MG cells (a glioblastoma multiforme cell line) were treated with necrotic ones, and miR-4461 was upregulated among the total 106 dysregulated miRNAs. 46 In addition, Chen et al. 24 published an article on colorectal cancer, in which miR-4461 was downregulated in bone marrow mesenchymal stem cell-exosomes, and had an inhibitory effect on the tumorigenesis of colorectal cancer by downregulating COPB2. Thus, it can be seen that miR-4461 possibly holds opposite effects on the progression of different cancer. Then, in this study, the authors found that miR-4461 showed an aberrantly low expression level in RCC tissues and cells, suggesting that miR-4461 could be an RCC suppressor. The cell functional experiments proved that miR-4461 could restrain RCC cell proliferation and adhesion and facilitate cell apoptosis, further highlighting the tumor-suppressive role of miR-4461 in RCC.
PPP1R3C, as an important regulatory protein of glucose metabolism, participates in the process of cancer by mediating glycogen synthesis or gene methylation. 35,36 For example, an article reported that the expression of PPP1R3C was inhibited in cervical cancer due to frequent DNA methylation, implying that PPP1R3C could act as a cervical cancer-related candidate gene to take part in the tumorigenesis and progression of cervical cancer. 37 However, Lee et al. uncovered that the knockdown of PPP1R3C inhibited the proliferation of colon cancer cells at high glucose levels, and the methylation of PPP1R3C was inhibited at high glucose levels. 25 This report seemed to be consistent with the authors' findings. In this study, the expression of PPP1R3C was upregulated in RCC. After transfection with si-PPP1R3C, the viability, proliferation, and adhesion of RCC cells were inhibited, while the apoptosis of RCC cells was accelerated. In other words, PPP1R3C played a key regulatory role in RCC as a cancer promoter. In addition, miR-4461 was identified as an upstream regulatory miRNA of PPP1R3C, and PPP1R3C expression was inhibited by miR-4461. Furthermore, miR-4461 exerted an inhibitory effect on the malignant phenotypes of RCC cells by targeting and inhibiting PPP1R3C.
More interestingly, although SW839 and OSRC-2 cell lines are different renal cancer cell lines, using FAM-labeling technique, the authors demonstrated that SW839 and OSRC-2 cell lines presented comparable transfection efficiency of PPP1R3C siRNA, miR-4461 mimic, and miR-4461 inhibitor. In the basis of this phenomenon, SW839 and OSRC-2 cell lines showed a striking similar cellular response in the loss and gain of function experiments. This finding might contribute to the selection of optimal renal cell lines in the future studies of RCC.
However, the research had certain limitations. Carcinogenesis was a very complex signaling pathway, and the authors did not involve the study of signaling pathways. In addition, this study did not further explore whether miR-4461 affected tumor growth and metastasis in vivo. Therefore, in the follow-up experiments, the role and regulatory mechanism of miR-4461 on RCC in vivo have to be confirmed, as well as the role of miR-4461 in cell signaling pathways. At the same time, PPP1R3C protein affects glycogen biosynthesis by activating glycogen synthase and limiting glycogen breakdown by reducing glycogen phosphorylase activity; thus, whether this can be regulated by miR-4661 in RCC in RCC is worth addressing in the future study.
Conclusions
Taking together, this study was the first to uncover that miR-4461, as a cancer inhibitor, was markedly linked with RCC progression in vitro. The authors' work illustrated that miR-4461 could be a new therapeutic target in the suppression of RCC progression, at least partially, by targeting PPP1R3C. These findings might help us to better understand the pathogenesis of RCC, and might provide new ideas for the treatment of RCC.
Footnotes
Ethics Approval
This study was approved by the Ethics Committee of Wuhan Third Hospital (Wuhan, China). All patients signed written informed consent.
Authors' Contributions
D.L. designed the study. Y.Y.Z. performed most of the experiments. G.Y. and Y.W. collected the data and wrote the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.