
Abstract
Background:
Mesenchymal stem cells (MSCs) could inhibit the proliferation of lung cancer cells. The authors' study investigated the effects of immunologically activated human umbilical cord (HUC)-MSCs on A549 lung cancer cells.
Materials and Methods:
HUC-MSCs were separated from the umbilical cord using the adherence method. Surface markers of HUC-MSCs were detected by flow cytometry for MSC identification. Imiquimod (TLR7 agonist) was incubated with HUC-MSCs for immune activation, and the expression of MSC-specific markers and immune inflammatory molecules was measured by quantitative real-time polymerase chain reaction. HUC A549 cells were cocultured with HUC-MSCs treated with imiquimod, siTLR7 (small interfering RNA for TLR7) or TLR7 overexpression, and then cell viability, proliferation, migration, and invasion, and the expression of phosphatidylinositol-3-kinase (PI3K)/Akt and NF-κB was investigated using MTT assay, clone formation assay, transwell assay, and Western blot, respectively.
Results:
HUC-MSCs were identified as positive for CD73, CD105, CD44, CD29, and CD90. Expression of MSC markers was inhibited, while those of immune inflammatory molecules expression except IL-6 (interleukin-6) was enhanced after MSCs were immunologically activated by imiquimod. After being cocultured with HUC-MSCs treated with imiquimod or overexpressed TLR7, cell viability, proliferation, and metastasis, and the phosphorylation of P65 and AKT in A549 cells were decreased, but apoptosis was increased, while siTLR7 showed the opposite effect HUC.
Conclusions:
Immunologically activated HUC-MSCs inhibited the growth and metastasis, yet, promoted the apoptosis of A549 lung cancer cells via regulating the PI3K/Akt and NF-κB pathways.
Introduction
Lung cancer is projected to be ranked as the sixth leading cause of death in the next decades, 1 and is the leading cause of cancer deaths in the United States. 2 Lung cancer is often diagnosed in a metastatic stage IV. 3 Traditional surgery, chemotherapy, and mechanism-based drugs are commonly used for lung cancer therapy; however, the survival rate of patients with lung cancer remains low and most therapies are accompanied by side-effects and drug resistance. 4,5 Thus, it is critical to develop new treatment methods for lung cancer.
Mesenchymal stem cell (MSC), a typical, representative and important member of the adult stem cell family, mainly exists in the connective tissue or organ stroma of the whole body and can further differentiate into mesenchymal cells, which can be used to repair different tissues and organs. 6,7 It has been reported that MSCs can also home to tumor sites and contribute to tumor growth and progression. 8 For example, Keramidas et al. showed that MSCs derived from human bone marrow could inhibit the proliferation of lung cancer cells in mouse tumor models. 9 Furthermore, MSCs derived from bone marrow in rats inhibited tumor growth in mouse lung cancer models. 10
In addition, compared with human bone marrow MSCs, human umbilical cord mesenchymal stem cells (HUC-MSCs) have obvious advantages, such as abundant source, simple sampling operation, low probability of pathogen contamination, strong ability of in vitro amplification, low immunogenicity, and strong immunosuppression ability. 11,12 The connection between HUC-MSCs and various cancers has been shown. For instance, Ciavarella et al. 13 found that HUC-MSCs had anti-inflammatory and antiproliferative effects on multiple myeloma (MM) in MM-bearing mice. Moreover, it has been reported that suppression of Burkitt's lymphoma cell proliferation is induced by HUC-MSCs, which emphasizes the native tumoricidal property of HUC-MSCs in hematological malignancies. 14
In addition, Zhang et al. 15 found that HUC-MSCs could be used as cell-based delivery vehicles for lung cancer therapy, indicating that interleukin (IL)-24-MSCs are able to inhibit the growth of A549 human nonsmall cell lung cancer cells in vitro and suppress xenograft lung tumor growth in vivo. In addition, on the basis of the study results of Liu et al., 16 UC-MSCs inhibited the cytotoxicity of activated Vγ9Vδ2 T cells against influenza virus-infected A549 cells. However, how enhanced immunogenicity of UC-MSCs influences the viability, proliferation, migration, and invasion of A549 cells remains inadequately clear.
Toll-like receptors (TLRs) are fundamental elements of immune system, which facilitate the understanding of innate and adaptive immunity. 17,18 TLR signal transduction pathway is involved in the occurrence and development of inflammatory diseases, 19 autoimmune diseases, 20 tumors, 21 and other immune-related diseases, which may aggravate immune damage of the body by initiating an abnormal immune response or maintaining chronic infection.
Furthermore, TLR7 agonist has also been reported to possess an immunostimulatory capacity and applied in the research of vaccines against several types of tumors, such as leukemia and breast cancer. 17 TLR7 agonist has been shown to induce UC-MSC immune activation by upregulating the expression of surface markers CD86 and HLA-E, promoting those of molecules that mediated immune inflammatory response and inhibiting MSC markers of UC-MSCs, such as CD29 and CD90. 22 Importantly, Jing et al. 23 found that UC-MSCs after immune activation had an antitumor effect on pancreatic tumor. However, the effect of immunologically activated UC-MSCs on lung cancer still needs further research.
AKT is acknowledged as the major downstream effector of phosphatidylinositol-3-kinase (PI3K), and overactivation of the PI3K/AKT signaling pathway is frequently present in human malignancies and cancer progression. 24 UC-MSC paracrine decreased the deposition of fibronectin by inhibiting cell proliferation mediated by the PI3K/Akt signaling pathways in mesangial cells. 25
This study introduced the TLR7 agonist imiquimod to induce immune activation of HUC-MSCs, and then investigated the effects of HUC-MSCs on the viability, proliferation, migration, and invasion of A549 cells, as well as the PI3K/Akt pathway.
Materials and Methods
Ethical statement
The study was approved by the Ethics Committee of The Sixth Affiliated Hospital of Wenzhou Medical University (Approval No. FCK20181124). Written informed consents were obtained from all the participants in any experimental work involving humans.
Clinical sample
The clinical samples of HUC-MSCs were collected from the umbilical cord (15 cm) of three full-term neonates who were born from healthy parturient women in The Sixth Affiliated Hospital of Wenzhou Medical University in 2018. All of these parturient women, who had signed informed consent, tested negative in HIV antibody test and Treponema Pallidum antibody test. All clinical samples were obtained at the initial resection, washed with phosphate-buffered saline (PBS), and stored at −80°C.
HUC-MSC separation and cell culture
The method used to isolate MSCs from the umbilical cord was based on published literature. 26 After being rinsed with PBS and then soaked in PBS containing penicillin–streptomycin solution (Lunan Pharmaceutical Group, China) for 15 min, the umbilical cord was diced into pieces (1 mm3). The small pieces of mesenchymal tissues were put into 10 × 35 mm plates, and then a small amount of L-DMEM (low-glucose Dulbecco's modified Eagle's medium; Gibco) containing penicillin–streptomycin solution was added to the plates and incubated at 37°C with 5% CO2. After about 15 d of culture, the HUC-MSCs showed a finger pattern or circinate growth, and after about 20 d of culture, the cells reached 80%–90% fusion. The first passage (P1) of HUC-MSCs was recorded in continuous culture. The morphological characteristics of HUC-MSCs on day 10 and 20 after culturing the mesenchymal tissues and those of P1 HUC-MSC were observed using a DMi8 optical microscope (Leica, Germany).
Human lung cancer cells (A549, Catalog No. CCL-185) were purchased from the American Type Culture Collection (ATCC, Manassas, VA). The cells were cultured in DMEM (Sigma) supplemented with 10% fetal bovine serum (FBS; Thermo Fisher Scientific, Waltham), and 100 U/mL penicillin and 100 μg/mL streptomycin at 37°C with 5% CO2.
Identification of HUC-MSCs
P4 (Passage 4) HUC-MSCs were prepared and then placed into 10 tubes at 2.5 × 105 per tube in PBS. Flow cytometry was used to identify the immune phenotypes of the fourth-generation HUC-MSCs, which were labeled with monoclonal antibodies such as FITC-conjugated CD73 antibody (ab239246; Abcam, Cambridge, United Kingdom), FITC-conjugated CD105 antibody (ab18278; Abcam), FITC-conjugated CD44 antibody (103021; BioLegend®), FITC-conjugated CD29 antibody (MCA1949T; Bio-Rad), FITC-conjugated CD90 antibody (ab11155; Abcam), FITC-conjugated HLA-DR antibody (361705; BioLegend), FITC-conjugated CD14 antibody (301803; BioLegend), FITC-conjugated CD34 (343603; BioLegend) and FITC-conjugated CD45 antibody (304005; BioLegend). IgG1-FITC antibody (ab97240; Abcam) was used as an isotype control. The cells and antibodies in each tube were mixed thoroughly and incubated at 4°C for 30 min. The cells were then rinsed in PBS and resuspended in 500 μL of PBS before detection by flow cytometry (MoFloAstrios EQ; Beckman Coulter).
Cell treatment
Imiquimod (TLR7 agonist; IMG-2207; Imgenex) was added into HUC-MSCs at a concentration of 10 μg/mL and incubated for 4, 12, 24, 72, and 120 h at 37°C.
Cell transfection
pcDNA3.1 plasmid was purchased (VT1010; YouBio, China) and the construction of the TLR7 overexpressed pcDNA3.1 plasmid was accomplished by Genomeditech. The small interfering RNA for TLR7 (siTLR7; sc-40266; Santa Cruz Biotechnology), scrambled siRNA (siNC), TLR7 overexpressed pcDNA3.1 plasmid (TLR7), and pcDNA3.1 plasmid (NC) were separately transfected into HUC-MSCs using Lipofectamine 2000 Transfection Reagent (Invitrogen, Carlsbad, CA) under the manufacturer's instructions.
HUC-MSCs were incubated into six-well plates at the density of 1 × 105 cells per well, and then incubated for 24 h. Then, 1.5 mL of medium without serum or antibiotics was added to each well, and a mixture of 500 μL siTLR7, siNC, TLR7, or NC was separately added in the wells with Lipofectamine 2000 Transfection Reagent. Later, the cells were incubated for 4–6 h at 37°C with 5% CO2. The siRNA sequences, which were used in the study were shown as follows: TLR7 siRNA: sense 5′-GAGGAAUUAGACAU CUCUAdTdT-3′, antisense 5′-GAGGAAUUAGACAU CUCUAdTdT-3′; si-negative control (siNC): sense 5′-AGAGTAATGGTACA CACTTdTdT-3′, antisense 5′-ATGAGTAGAGACTA CCATTdTdT-3′.
Quantitative real-time polymerase chain reaction
Total RNAs in HUC-MSCs treated with imiquimod for 4, 12, 24, 72, and 120 h were extracted to measure the mRNA expression of MSC-specific markers TP63, OCT4, Nestin, and Lin28, and those of immune inflammatory factors IL-6, IL-8, IL-12, tumor necrosis factor-α (TNF-α), transforming growth factor-β (TGF-β), and interferon-β (IFN-β). Total RNAs in HUC-MSCs transfected with siTLR7 or siNC, or treated with imiquimod, or without any treatment were extracted to measure the mRNA expression of immune inflammatory factors IL-6, IL-8, IL-12, TNF-α, TGF-β, and IFN-β.
Total RNAs were extracted using TRIzol reagent (T9424; Sigma) according to the instructions and then placed in the refrigerator at 4°C or −80°C. Later, a micro-UV-visible spectrophotometer (DR6000; Hach) was used to quantify RNA concentration. The extracted RNAs were reverse transcribed to cDNAs using a Prime Script RT reagent Kit (4368813; Thermo Fisher Scientific). The mRNA expression was determined using the standard SYBR-Green RT-PCR kit (Takara Biotechnology Co., Ltd., Dalian, China) and GAPDH was used as the internal control. The thermal cycle was set as follows: pretreatment at 94°C for 2 min, followed by at 94°C for 30 s, at 63°C for 30 s, 35 cycles of at 72°C for 1 min, at 72°C for 7 min, and finally kept at 4°C. 27 The primer sequences for the experiment are shown in Table 1. The gene expression was calculated using the 2−ΔΔCt method.
Primer for Quantitative Real-Time Polymerase Chain Reaction
Coculture of HUC-MSCs and A549 cells
A noncontact coculture system of HUC-MSCs and A549 cells was established by using a Transwell suspension culture chamber with a 6-pore plate (0.4 μm, Corning 3450; Corning, Inc., Corning, NY). HUC-MSCs transfected with siTLR7, TLR7, siNC, or NC, or treated with imiquimod or without any treatment, were incubated in the lower chamber at the density of 6 × 105 cells per pore, and a total of 3 × 105 A549 cells were cultured in the upper chamber for 24 h. Cell culture was terminated on day 7 of culture, and single cell suspensions were prepared for detection.
MTT assay
After coculture with HUC-MSCs with different treatments (transfected with siTLR7, TLR7, siNC, or NC, or treated with imiquimod, or without any treatment), A549 cells were cultured in a 96-well plate at the density of 5 × 105 cells per well for 48 h, and later 10 μL of MTT reagent (B7777; ApexBio) was added to each well. Then, the cells were incubated for 4 h at 37°C. The formazan product was dissolved in 1.5 mL of dimethylsulfoxide and then a multifunctional enzyme-linked analyzer (HBS-1096C; Detie, China) was used to detect for 4 h, and the absorbance value of each well was measured at 490 nm.
Clone formation assay
After coculture with HUC-MSCs with different treatments (transfected with siTLR7, TLR7, siNC, or NC, or treated with imiquimod or without any treatment) at the density of 1 × 102 cells per well were cultured in a 96-well plate at 37°C with 5% CO2 under saturated humidity. After being incubated for 2–3 weeks, the cells were carefully dipped and washed in PBS twice. Next, the cells were fixed with 5 mL of methanol for 15 min and subsequently stained with Giemsa for 30 min. Visible colonies to the naked eye were observed under a microscope (XSP-05; Zhanjing Optical Instruments Co., Ltd., Ningbo, China). The image of each colony was collected and the clone formation rate was calculated.
Flow cytometry
According to the manufacturer's protocols, A549 cell apoptosis was measured using an Annexin V-FITC Apoptosis Detection Kit (APOAF-50TST; Sigma). A549 cells were first cultured for 48 h and then after coculture with HUC-MSCs with different treatments (transfected with siTLR7, TLR7, siNC, or NC, or treated with imiquimod or without any treatment), the cells were washed with cold PBS twice. Then, the cells were processed with Annexin V and propidium iodide together in the dark at room temperature for 15 min. Cell apoptosis was then analyzed using the flow cytometer (NovoCyte; ACEA Biosciences), and NovoExpress Software (ACEA Biosciences) was used for data analysis.
Transwell
A Matrigel (BD, San Diego) was diluted and covered the upper surface of the polycarbonate film covering the bottom of the Transwell chamber, and then placed at 37°C for 30 min to make Matrigel polymerize into gel. To detect the invasion of A549 cells after coculture with HUC-MSCs with different treatments (transfected with siTLR7, TLR7, siNC, or NC, or treated with imiquimod or without any treatment), the cells at the density of 1 × 105 per well were collected and resuspended in serum-free DMEM in the upper chambers of the Transwell (8-μm pore size; Corning) insert, whose lower chambers were filled with DMEM containing 600 μL of 20% FBS that was used as a chemoattractant.
After a 24 h incubation, the noninvading cells in the upper chamber were wiped off gently with cotton swabs. Later, the upper chamber was fixed in 4% paraformaldehyde for 15 min and stained with 0.1% crystal violet for 20 min. The cells were selected and observed under a light microscope (XSP-05; Zhanjing Optical Instruments Co., Ltd.) and photographed.
The migration of A549 cells was detected in a similar manner after the cells were cocultured with HUC-MSCs with different treatments (transfected with siTLR7, TLR7, siNC, or NC, or treated with imiquimod or without any treatment), except that the upper side of the polycarbonate film was not spread with Matrigel. The cell migration rate was calculated by average migrated cells in Control group × 100%.
Western blot
A549 cells were collected after coculture with HUC-MSCs with different treatments (transfected with siTLR7, TLR7, siNC or NC, or treated with imiquimod or without any treatment). RIPA lysis buffer (P0013B; Beyotime, Shanghai, China) was used to extract the protein, and then protein concentration was determined with the bicinchoninic Protein Assay Kit (AR0146; Boster, Pleasanton, CA). Next 30 μg of the extracted protein samples were electrophoresed by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE; Zoonbio Biotechnology, China) and transferred to polyvinylidene fluoride membranes (Kynar 710; Arkema, France), 28 which were blocked with 5% fat-free milk for 1 h at room temperature.
The membranes were then incubated overnight at 4°C with the primary antibodies, including anti-phosphorylated-p65 (p-P65) antibody (rabbit, ab86299, 1:2000; Abcam), anti-P65 antibody (rabbit, ab16502, 0.5 μg/mL; Abcam), anti-Akt antibody (rabbit, ab8805, 1:500; Abcam), anti-p-Akt (phosphorylated-AKT) antibody (rabbit, ab38449, 1:500; Abcam), and anti-GAPDH antibody (mouse, ab8245, 1:500; Abcam). GAPDH was used as the internal reference. The membranes were then incubated with the secondary horseradish peroxidase (HRP)-conjugated antibodies: goat anti-rabbit IgG H&L (HRP) (goat, ab6721, 1:2000; Abcam), and goat anti-mouse IgG H&L (HRP) (goat, ab205719, 1:2000; Abcam) for 1 h at room temperature and washed with tris-buffer saline tween for three times.
The protein bands were analyzed using the enhanced Chemiluminescence Kit (ZD310-1; Zoman Biotechnology, China) and the gray values of the strips were scanned using ImageJ (version 5.0; Bio-Rad).
Statistical analyses
All statistical analyses were performed using SPSS ver. 18.0 software (SPSS, Inc., Chicago, IL). The data are presented as the mean ± standard deviation and the experiments have been carried out three times. One-way analysis of variance (ANOVA) was used to compare the differences between multiple groups followed by Dunnett's post hoc test. p < 0.05 indicated statistical significance.
Results
Identification of HUC-MSCs
As revealed in Figure 1A, after the umbilical cord tissues were incubated for 10 d, there were a few irregular cells and spindle-shaped adherent cells near the tissue. On day 20, cells grew denser and showed a finger pattern or circinate growth. P1 HUC-MSCSs were recorded in continuous culture (Fig. 1A). Flow cytometry was used to detect the surface markers of HUC-MSCs (P4). The results demonstrated that in HUC-MSCs, CD73 (93.7%), CD105 (95.6%), CD44 (96.6%), CD29 (92.9%), and CD90 (97.3%) were high expressed, but HLA-DR (0.918%), CD14 (2.21%), CD34 (2.50%), and CD45 (1.90%) were not expressed (Fig. 1B).

HUC-MSCs were identified using stemness markers identification after being isolated from umbilical cord tissues.
Effects of the TLR7 agonist imiquimod on stemness markers and inflammatory response-related genes in HUC-MSCs
HUC-MSCs were added with 10 μg/mL imiquimod (TLR7 agonist) and then the expression of stemness markers and molecules that mediate immune inflammatory responses in HUC-MSCs was detected by quantitative real-time polymerase chain reaction (qRT-PCR) after incubation for 4, 12, 24, 72, and 120 h. According to the results of qRT-PCR, the mRNA expression of specific markers TP63, OCT4, Nestin, and Lin28 in HUC-MSCs was significantly inhibited with time by the TLR7 agonist imiquimod (Fig. 2A–D, p < 0.05, vs. Control).

Effects of imiquimod on HUC-MSCs were evaluated by levels of stemness markers and inflammation factors.
As for the mRNA expression of molecules that mediates immune inflammatory response, IL-6 expression was increased with time by the TLR7 agonist imiquimod, while the expression of IL-8, IL-12, TNF-α, TGF-β, and IFN-β was upregulated at first but showed a decreasing tendency after 24 h under the treatment of the TLR7 agonist imiquimod (Fig. 2E–J, p < 0.05, vs. Control).
Expression of inflammatory factors in HUC-MSCs was promoted by imiquimod, which was reversed by siTLR7
In the Imiquimod group, HUC-MSCs treated with 10 μg/mL TLR7 agonist imiquimod were cocultured with A549 cells for 24 h. Also, in the siTLR7 group, HUC-MSCs treated with siTLR7 were cocultured with A549 cells for 24 h. The results of qRT-PCR showed that the mRNA expression of IL-6, IL-8, IL-12, TNF-α, TGF-β, and IFN-β in HUC-MSCs was increased in the Imiquimod group (Fig. 3A–F, p < 0.05, vs. Control), but decreased in the siTLR7 group (Fig. 3A–F, p < 0.05, vs. siNC).

Expression of inflammatory factors in HUC-MSCs under imiquimod or siTLR7 treatment after coculture with A549 cells was evaluated using qRT-PCR.
Imiquimod-treated HUC-MSCs inhibited the viability, proliferation, and metastasis but promoted the apoptosis of A549 cells through TLR7
A549 cells were collected, and MTT assay and clone formation assay were applied to measure the viability and proliferation of the cells. The results revealed that cell viability (Fig. 4A, p < 0.05 vs. Blank) and proliferation (Fig. 4B, p < 0.01 vs. Blank) in the Control group showed a downward trend. Interestingly, cell viability (Fig. 4A, p < 0.001, vs. Control) and numbers of colonies formed in the Blank group (Fig. 4B, p < 0.001, vs. Control) exhibited a further reduction. However, in the siTLR7 group, cell viability (Fig. 4A, p < 0.05, vs. siNC) and relative colony formation rates (Fig. 4B, p < 0.01. vs. siNC) were obviously enhanced, suggesting that siTLR7 in HUC-MSCs exerted the opposite function of imiquimod.
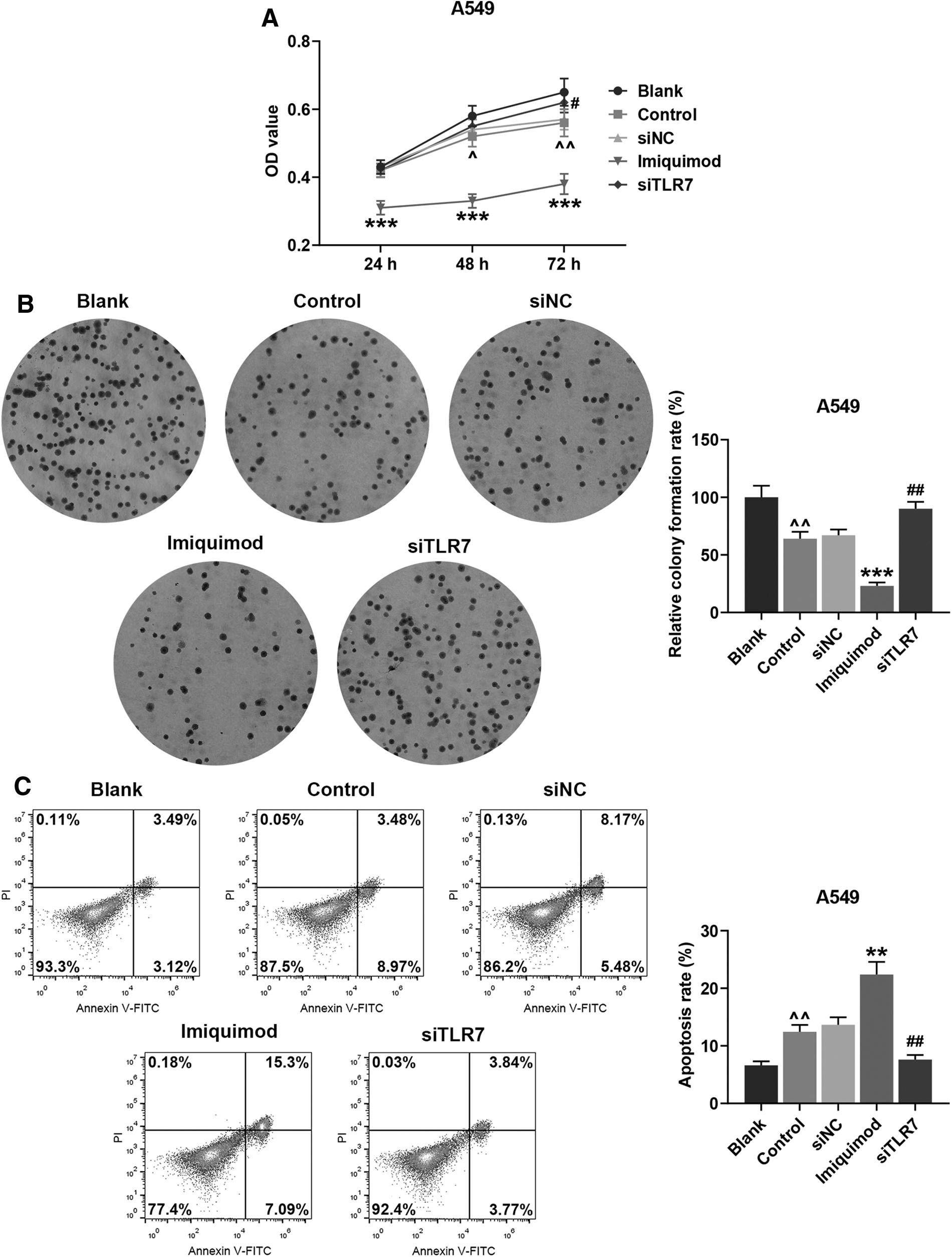
Effects of HUC-MSCs under imiquimod or siTLR7 treatment on the cell viability, proliferation, and apoptosis rate of A549 cells were evaluated.
At the same time, flow cytometry was used to detect A549 cell apoptosis and Transwell was used to measure cell migration and invasion rates. In light of the results, it was clear that the cell apoptosis rates in the Control group (Fig. 4C, p < 0.01, vs. Blank) and the Imiquimod group (Fig. 4C, p < 0.01, vs. Control) were raised, but that in the siTLR7 group (Fig. 4C, p < 0.01, vs. NC) was decreased. Meanwhile, after A549 cells were incubated in a small chamber for another 24 h, the cell migration and invasion rates in the Control group (Fig. 5A, B, p < 0.05, vs. Blank) and the Imiquimod group (Fig. 5A, B, p < 0.001, vs. Control) were decreased, while those in the siTLR7 group (Fig. 5A, B, p < 0.05, vs. siNC) were increased.

Effects of HUC-MSCs under imiquimod or siTLR7 treatment on A549 cells migration rate and invasion rate detection during coculture.
Imiquimod-treated HUC-MSCs suppressed the phosphorylation of P65 and AKT in A549 cells, which was reversed by siTLR7
In this phase, Western blot was applied to investigate the protein expression of PI3K/Akt and NF-κB. As revealed in Figure 6A, the protein expression of p-P65 and p-AKT was reduced in the Control group (p < 0.001, vs. Blank) and the Imiquimod group (p < 0.001, vs. Control) but promoted in the siTLR7 group (p < 0.001, vs. siNC). Furthermore, the ratios of p-P65 to P65 and p-AKT to AKT were decreased in the Control group (Fig. 6A, B, p < 0.05, vs. Blank) and the Imiquimod group (Fig. 6A, B, p < 0.001, vs. Control) but increased in the siTLR7 group (Fig. 6A, B, p < 0.01, vs. siNC). Based on these results, the authors could draw a conclusion that phosphorylation of P65 and AKT was suppressed in A549 cells when HUC-MSC immune activation occurred.
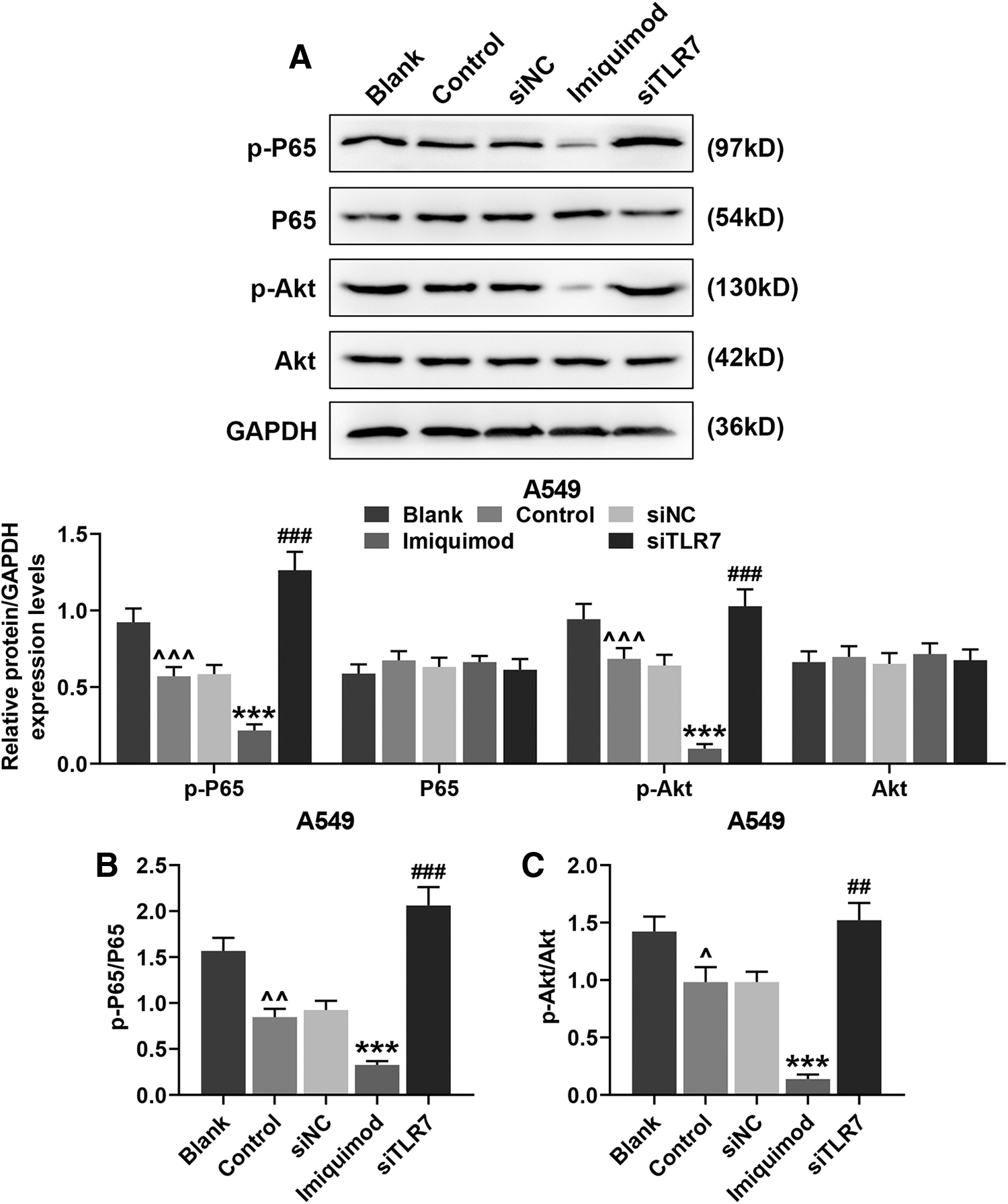
Effects of HUC-MSCs under imiquimod or siTLR7 treatment on phosphorylation of PI3K/Akt and P65 in A549 cells during coculture.
HUC-MSCs transfected with overexpressed TLR7 inhibited the viability, proliferation, and migration, yet, enhanced the apoptosis of A549 cells
To further confirm the role of TLR7 in the viability, proliferation, and apoptosis of A549 cells, the TLR7 level was upregulated in HUC-MSCs by the overexpressed TLR7 plasmid. In this phase, HUC-MSCs and A549 cells were cocultured for 24 h in the Control group, and HUC-MSCs transfected with overexpressed TLR7 and A549 cells were cocultured for 24 h in the TLR7 group. Through MTT and clone formation assays, the authors found that A549 cell viability and proliferation in the Control group (Fig. 7A, B, p < 0.05, vs. Blank) and the TLR7 group (Fig. 7A, B, p < 0.01, vs. NC) was decreased with time. On the contrary, the result of flow cytometry demonstrated that the apoptosis rates of A549 cells in the Control group (Fig. 7C, p < 0.01, vs. Blank) and TLR7 group (Fig. 7C, p < 0.01, vs. NC) were increased with time.

Effects of HUC-MSCs with overexpressed TLR7 on A549 cell viability, proliferation, and apoptosis during coculture.
HUC-MSCs transfected with overexpressed TLR7 suppressed the phosphorylation of P65 and AKT in A549 cells
Meanwhile, through Western blot, it was found that the protein expression of p-P65 and p-AKT in A549 cells was lowered in the Control group (Fig. 8A, p < 0.01, vs. Blank) and the TLR7 group (Fig. 8A, p < 0.001, vs. NC). Furthermore, the ratios of p-P65 to P65 and p-AKT to AKT were decreased in the Control group (Fig. 8B, C, p < 0.05, vs. Blank) and the TLR7 group (Fig. 8B, C, p < 0.01, vs. NC) but increased in the siTLR7 group (Fig. 8B, C, p < 0.01, vs. siNC). These results indicated that the phosphorylation of P65 and AKT was suppressed in A549 cells by overexpressed TLR7 in HUC-MSCs.

Effects of HUC-MSCs transfected with overexpressed TLR7 plasmid on phosphorylation of PI3K/Akt and P65 in A549 cells after coculture.
Discussion
MSC-derived extracellular vesicles have therapeutic effects on various preclinical models of lung diseases. 29 However, previous studies have shown that under certain conditions, transplanted MSCs are subject to a variety of molecularly mediated immunogenic responses in vivo. 30 Besides, in the authors' study, surface markers of P4 HUC-MSCs were detected by flow cytometry, and the results demonstrated that stemness surface markers either had high expression in HUC-MSCs, such as CD73, CD105, CD44, CD29, and CD90, or had no expression in HUC-MSCs, such as HLA-DR, CD14, CD34, and CD45, which testified that the phenotypes of MSCs derived from HUC were consistent with those from bone marrow, placenta, and other tissues. The results were in accordance with those of the prior study. 31
TLRs are a family of innate immune receptor recently discovered and widely studied. TLRs play a key role in initiating innate immune responses and also modulate acquired immunity by identifying pathogen-associated molecular patterns such as bacteria and viruses. 32,33 Among the 11 TLRs, TLR7 was located on the surface of endosome and could be recognized and activated by single stranded RNA of RNA viruses. 34,35 Activated TLR7 induces a cascade of reactions through signal transduction that eventually lead to the synthesis and secretion of type I IFN and inflammatory molecules, mediating an antiviral immune response that inhibits viral replication and plays an important role in protecting the organism. 36
Zhang et al. 26 found that the activation of the TLR7 pathway by specific agonist imiquimod increases the immunogenicity of UC-MSCs and suggested that the enhanced immunogenicity of UC-MSCs might cause a change of immune status in UC-MSCs-tumor environment. Furthermore, a previous study reported that TLR7 agonist CL264 could increase the expression of UC-MSC surface costimulatory molecules CD86 and HLA-E, and that multiple proinflammatory response factors (IL-1β, IL-6, IL-8, IL-10, IFN-β, IFN-γ, NF-κB, and TGF-β) could be significantly induced by CL264, and the expression of the stem cell markers (Klf4, Nestin, Sox2, and Lin28) of UC-MSCs was significantly inhibited, indicating that CL264 significantly increased the immunogenicity of UC-MSCs. 22
As for the study, the mRNA expression of stemness markers TP63, OCT4, Nestin, and Lin28 in HUC-MSCs was inhibited by the TLR7 agonist imiquimod. Previous studies have demonstrated that TP63 plays an important role in the development of many malignancies 37 ; inhibition of LIN28 may be useful in preventing immune evasion of cancer cells 38 ; and the expression of OCT4 and Nestin correlates with the growth, metastasis, and poor prognosis of aggressive tumors 39 ; these findings suggested that inhibited TP63, OCT4, Nestin, and Lin28 function as tumor suppressors.
Interestingly, as for the mRNA expression of molecules that mediate immune inflammatory response, IL-6 expression was increased with time by the TLR7 agonist imiquimod, and the expression of IL-8, IL-12, TNF-α, TGF-β, and IFN-β was upregulated at first, but showed a decreasing tendency after 24 h under the treatment of the TLR7 agonist imiquimod. To the best of the authors' knowledge, in inflammatory response, the expression of IL-6 increases earlier and for a longer time than those of other cytokines, 40 which was consistent with the results of the study. Importantly, it has been reported that UC-MSCs may have an antitumor effect on pancreatic tumor, given that they expressed murine IL15 in tumor-bearing mice. 23
Therefore, in light of the results of the studies above, they treated HUC-MSCs with the TLR7 agonist imiquimod and cocultured the cells with A549 cells. According to the results, the viability, proliferation, migration, and invasion of A549 cells were decreased, but their apoptosis was increased when HUC-MSCs were treated with the TLR7 agonist imiquimod. On the contrary, knockdown of TLR7 reversed the effect of imiquimod on A549 cells.
To the best of the author's knowledge, the PI3K/Akt and NF-κB pathways are important transcriptional regulators in the apoptotic signaling pathway and serve as central mediators to inhibit apoptosis. An imbalance in the PI3K/Akt pathway plays a key role in the formation and development of lung cancer. 41 Thus, it was speculated that the immune activation of MSC further affects the biological functions of lung cancer cells, which may be related to the PI3K/Akt and NF-κB pathways. 42,43 Therefore, after transfection and coculture, A549 cells were collected and detected by Western blot, and the results revealed that the phosphorylation of P65, one of the five transcription factors of NF-κB, as well as that of AKT, was suppressed in A549 cells. Their further investigation showed that the viability and proliferation of A549 cells were suppressed, but their apoptosis was increased by HUC-MSCs transfected with the overexpressed TLR7 plasmid. Furthermore, the phosphorylation of P65 and AKT was suppressed by HUC-MSCs with overexpressed TLR7.
Conclusions
In summary, the authors' study demonstrated the effective role of immunologically activated HUC-MSCs induced by the TLR7 agonist imiquimod in reducing the viability, proliferation, migration, and invasion of A549 cells and enhancing their apoptosis. These findings may provide a new therapeutic strategy for the treatment of lung cancer.
Footnotes
Authors' Contributions
Substantial contributions to conception and design: H.Y. and J.P. Data acquisition, data analysis, and interpretation: H.G., X.C., C.X., Y.L., H.Z., and Z.C. Drafting the article or critically revising it for important intellectual content: H.Y. and J.P. Final approval of the version to be published: all authors. Agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved: all authors.
Disclosure Statements
There are no existing financial conflicts.
Funding Information
This work was supported by the Major Project of Science and Technology Bureau of Lishui [Grant No. 2019ZDYF20]; the Basic Public Interest Research Project of Zhejiang Province [Grant No. LGF19H010006]. Thanks for the financial supports.