
Abstract
Background:
Increasing evidence enhanced the recognition of circular RNAs (circRNAs) implicated in cancer progression. CircRNA_0085616 (circ_0085616) is a novel circRNA with high expression in cervical cancer (CC); its function and action mechanism are explored during this study.
Methods:
The measurement of circ_0085616, microRNA-503-5p (miR-503-5p), and Ataxin-7L3 (ATXN7L3) was performed via quantitative real-time polymerase chain reaction. Next, 3-(4,5-dimethylthiazol-2-y1)-2, 5-diphenyl tetrazolium bromide (MTT) and colony formation assays were implemented to analyze the proliferative ability. Cell metastasis was evaluated by using transwell migration and invasion assays. Glycolysis was analyzed by using glucose consumption, lactate and adenosine triphosphate production. Western blot was employed for protein analysis. The target binding was performed by dual-luciferase reporter assay and RNA immunoprecipitation assay. An animal experiment in vivo was conducted by xenografts.
Results:
Circ_0085616 was heightened and its downregulation blocked cellular proliferation, metastasis, and glycolysis in CC. Then, we found that circ_0085616 had a sponge effect on miR-503-5p and the miR-503-5p inhibitor could ameliorate the effects of circ_0085616 knockdown on CC cells. In addition, miR-503-5p directly targeted ATXN7L3 to obstruct CC cell proliferation, metastasis, and glycolytic process. Further, circ_0085616 could enhance ATXN7L3 by sequestering miR-503-5p, and the miR-503-5p/ATXN7L3 axis was also responsible for circ_0085616 on promoting CC tumorigenesis in vivo.
Conclusion:
It was obvious that circ_0085616 facilitated the carcinogenic effect on CC via the activation of ATXN7L3 by sponging miR-503-5p. Our study may be likely to provide a novel molecular target for CC therapy.
Introduction
Cervical cancer (CC) is ranked as the fourth cancer regarding the incidence and mortality among females around the world. 1 The primary cause of CC is the persistent infection of human papilloma virus (HPV), and the key preventive tactics include cervical screening and HPV vaccination.2,3 In addition, surgical treatment and the concurrent chemoradiotherapy are usually used as the primary therapies for CC patients.4–6 However, the advanced-inoperable CC remains a severe challenge due to the increase of locoregional and distant recurrences. 7 Investigating the underlying pathomechanism of CC progression and finding out efficient biomarkers are quite necessary.
The noncoding RNAs, including circular RNAs (circRNAs) and microRNAs (miRNAs), have been involved in the initiative and progressive processes of CC. 8 CircRNAs are structurally characterized by the covalent closed-loop circuit and they act as “sponges” of miRNAs to be the crucial tumor-regulatory molecules. 9 Chen et al. found that circMTO1 contributed to the tumorigenesis and chemoresistance of CC by sponging miR-689310; Ma et al. identified circ_000284 as a promoter in proliferation and invasion of CC cells by acting as the sponge of miR-506. 11 hsa_circ_0085616 (circ_0085616) is overexpressed according to the heatmap of differentially expressed circRNAs in CC, 12 but it remains unknown regarding its function in CC. It has been reported that microRNA-503-5p (miR-503-5p) is a tumor inhibitor in CC 13 ; then, we wanted to analyze the potential relationship between circ_0085616 and miR-503-5p in CC.
As we all know, miRNAs can bind to the 3′ untranslated regions (3′UTRs) of messenger RNAs (mRNAs) to mediate the expression of genes in cancers. 14 Ataxin-7L3 (ATXN7L3) was overexpressed in CC as an oncogene, and it worked as a downstream gene of miR-877-5p. 15 Herein, whether ATXN7L3 can be a molecular target of miR-503-5p will be addressed. Meanwhile, the functional mechanism of circ_0085616 with miR-503-5p and ATXN7L3 is uncovered to exploit a novel signal-regulated pathway.
Materials and Methods
Tissue acquisition and cell culture
Seventy CC patients at the First Affiliated Hospital of Bengbu Medical College have enrolled in our study and signed the written informed consent to donate their tissues for medical research. Chemotherapy, radiotherapy, or immunotherapy has not been used for these patients and they finally suffered the surgical resection. In addition to obtaining CC tissues (n = 70), those distal tissues (n = 40; > 2 cm from cancer area to avoid the encroachment of cancer cells) were collected as normal controls. Promptly, these specimens were snap-frozen in liquid nitrogen and then saved in an ultra-low temperature refrigerator at −80°C. The sample collection was administrated in conformity with the guidelines of the Declaration of Helsinki, and this study was empowered by the Ethical Committee of the First Affiliated Hospital of Bengbu Medical College.
Four human CC cell lines (HeLa, SiHa, C33A, and CaSki) and normal ectocervical cell line ECT1/E6E7 purchased from American Type Culture Collection (ATCC, Manassas, VA, USA) were all cultivated in the complete medium of Dulbecco's modified eagle medium (DMEM; Gibco, Carlsbad, CA, USA), 10% fetal bovine serum (FBS; Gibco), and the antibiotic solution (100 U/mL penicillin and 100 μg/mL streptomycin; Gibco) under the conditions of 37°C, 5% CO2, and 95% air.
Cell transfection
Small interfering RNA (siRNA) targeting circ_0085616 (si-circ_0085616#1, si-circ_0085616#2, and si-circ_0085616#3) and siRNA negative control (si-NC), short hairpin RNA (shRNA) targeting circ_0085616 (sh-circ_0085616) and shRNA NC (sh-NC), miR-503-5p mimic (miR-503-5p) and miRNA mimic NC (miR-NC), miR-503-5p inhibitor (anti-miR-503-5p) and miRNA inhibitor NC (anti-miR-NC) were synthesized from GenePharma (Shanghai, China). The overexpression vector pcDNA-ATXN7L3 was formed by cloning the sequence of ATXN7L3 into the empty vector pcDNA (Invitrogen, Carlsbad, CA, USA). According to the manufacturer's specification, transfection of oligonucleotides or vectors in SiHa and CaSki cells was conducted by Lipofectamine 3000 (Invitrogen).
The quantitative real-time polymerase chain reaction
RNAiso Plus (Takara, Beijing, China) was used for isolating total RNA, followed by the acquisition of the complementary DNA (cDNA) via PrimeScript™ RT Master Mix (Takara). Then, the quantification of circ_0085616, miR-503-5p, and ATXN7L3 was implemented on the Applied Biosystems 7500 Fast Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) by TB Green® Premix Ex Taq™ II (Takara) with the specific primers as given later: circ_0085616 (forward, 5′-CACCGAGGACTACAACTCGC-3′ and reverse, 5′-GCCTCAGTGAAAACCATCTGC-3′), miR-503-5p (forward, 5′-CCTATTTCCCATGATTCCTTCATA-3′ and reverse, 5′-GTAATACGGTTATCCACGCG-3′), and ATXN7L3 (forward, 5′-CAGGCACTTCTCCAGATGGG-3′ and reverse, 5′-CGGGCTTGGACATCTTTGGA-3′). Besides, β-actin (forward, 5′-GACCTGTACGCCAACACAGT-3′ and reverse, 5′-AGTACTTGCGCTCAGGAGGA-3′) acted as the endogenous control for circ_0085616 or ATXN7L3, as well as U6 (forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′) for miR-503-5p. The comparative cycle threshold (2−ΔΔCt) method 16 was used to calculate the fold changes of gene expression.
3-(4, 5-dimethylthiazol-2-y1)-2, 5-diphenyl tetrazolium bromide assay
Cell culture was performed in 96-well plates with 5 × 103 cells/well for 24 h. After adding 20 μL MTT (Sigma-Aldrich, St Louis, MO, USA) and discarding the supernatant, we added dimethyl sulfoxide (Sigma-Aldrich) with 200 μL/well. Then, the absorbance of 490 nm was determined via a microplate reader.
Colony formation assay
Five hundred cells in DMEM medium containing 10% FBS were seeded into six-well plates and incubated for 12 d in the incubator. Subsequently, the colonies were photographed by a camera and counted by using a microscope after fixation and staining in 4% paraformaldehyde and crystal violet (Beyotime, Shanghai, China).
Transwell assay
Serum-free DMEM medium was employed for suspension preparation of transfected cells. Then, cell suspension was seeded into the top chamber of a transwell chamber (Corning, Inc., Corning, NY, USA) with or without Matrigel (Corning, Inc.) package to determine the invasive or migratory ability, respectively. After the addition of DMEM medium with 10% FBS into the bottom chamber for 24 h, the membranes were fastened and dyed by 4% paraformaldehyde and crystal violet (Beyotime). Eventually, these migrated and invaded cells in randomly selected observational field were photographed with the magnification of 100 × and counted under the inverted microscope (Olympus, Tokyo, Japan).
Glycolytic analysis
To perform glycolytic analysis, the glucose uptake colorimetric assay Kit (Biovision, Milpitas, CA, USA) and lactate colorimetric assay kit (Biovision) were exploited for the detection of glucose consumption and lactate production. In addition, adenosine triphosphate (ATP) production was assessed by using ATP cell viability assay kit (Biovision) as per the provided operating manual.
Western blot
Proteins were obtained by lysing tissue samples; cells were cultured by using RIPA buffer (Beyotime); and finally, ATXN7L3 and the standard markers associated with proliferation, metastasis, and glycolysis were examined by Western blot following the previous description. 17 The specific primary antibodies (Abcam, Cambridge, United Kingdom) contained Ki67 (ab92742; 1:1000), proliferating cell nuclear antigen (PCNA; ab152112; 1:1000), Vimentin (ab137321; 1:1000), E-cadherin (ab15148; 1:1000), lactic dehydrogenase A (LDHA; ab84716, 1:1000), glucose transporter 1 (GLUT1; ab15309, 1:1000), ATXN7L3 (ab99947; 1:2000), and β-actin (ab8227; 1:3000). After the incubation of anti-Rabbit IgG H&L (HRP) (Abcam; ab205718, 1:5000) and protein coloration by ECL Substrate Kit (Abcam), the relative protein levels were analyzed on ImageLab software version 4.1 (Bio-Rad, Hercules, CA, USA) with β-actin as a housekeeping gene.
Bioinformatics analysis
The bioinformatics analysis between circ_0085616 and the possible miRNAs was performed by online circinteractome, circBank, and starbase 3.0. Also, starbase 3.0 software was used for the prediction of potential target genes of miR-503-5p.
Dual-luciferase reporter assay
SiHa and CaSki cells were transfected with the constructed luciferase reporter plasmids and miR-503-5p or miR-NC for 48 h. After cells were centrifuged at 14,000 rpm for 5 min and lysed in 1 × passive buffer (Promega, Madison, WI, USA), the relative luciferase detection (firefly/renilla) was conducted via the dual-luciferase assay kit (Promega). The luciferase reporter plasmids of wild-type (circ_0085616-WT and ATXN7L3 3′UTR-WT, containing the miR-503-5p binding sites) and mutant type (circ_0085616-MUT and ATXN7L3 3′UTR-MUT, containing the mutated sites of the miR-503-5p binding region) were constructed by using the pMIR-REPORT vector (Promega).
RNA immunoprecipitation assay
Imprint® RNA Immunoprecipitation Kit (Sigma-Aldrich) was applied for RNA immunoprecipitation (RIP) analysis in this article. Next, 2 × 106 SiHa and CaSki cells were lysed in RIP lysis buffer and incubated with Protein A magnetic beads precoated with anti-Argonaute-2 (anti-Ago2) or anti-immunoglobulin G (anti-IgG) at 4°C overnight. After RNA isolation, the expression levels of circ_0085616 and miR-503-5p were assayed by using quantitative real-time polymerase chain reaction (qRT-PCR).
Animal experiment
Overall, 4 × 106 SiHa cells with stable transfection of sh-circ_0085616 or sh-NC via lentiviral vector were, respectively, injected into the female BALB/c nude mice (5-week-old; Shanghai Animal Experimental Center, Shanghai, China) in a subcutaneous manner, with five mice per group. Every week postinjection, the length and width of tumors were accurately monitored by using a digital caliper, followed by the calculation of tumor volume via the formula as given next: length × width 2 × 0.5. At the end of the fifth week, all mice were sacrificed by using the CO2 asphyxia method and tumors were excised for weighing. Then, the levels of circ_0085616, miR-503-5p and ATXN7L3 in tumors were measured by using qRT-PCR or Western blot analysis after isolation of total RNA or protein. This mice experiment was authorized by the Animal Ethical Committee of the First Affiliated Hospital of Bengbu Medical College, and all operated procedures strictly followed the Guideline of Care and Use of Laboratory Animals.
Statistical analysis
SPSS 24.0 (IBM Corp., Armonk, NY, USA) and Graphpad Prism 7 (GraphPad, Inc., La Jolla, CA, USA) were exploited to process data in this study. Data were presented as the mean ± standard deviation according to the three independent experiments with N = 3. Spearman's correlation coefficient was applied for the linear analysis among circ_0085616, miR-503-5p, and ATXN7L3 in CC tissues. The difference analysis was performed through Student's t-test or one-way analysis of variance followed by Tukey's test. p < 0.05 indicated the significant difference at the statistical level.
Results
Circ_0085616 was dysregulated with a high level in CC tissues and cells
Regarding the expression level of circ_0085616 in CC, we performed the qRT-PCR to analyze it in tissues and cells. As per the description in Figure 1A, 70 CC tissues manifested the upregulation of circ_0085616 level by comparison to the corresponding 70 normal tissues. Similar to that in CC tissues, circ_0085616 expression was also significantly increased in CC cells (HeLa, SiHa, C33A, and CaSki) compared with ECT1/E6E7 cells (Fig. 1B). SiHa and CaSki cells with a higher level than HeLa and C33A cells were used as the research objects in the following experiments.
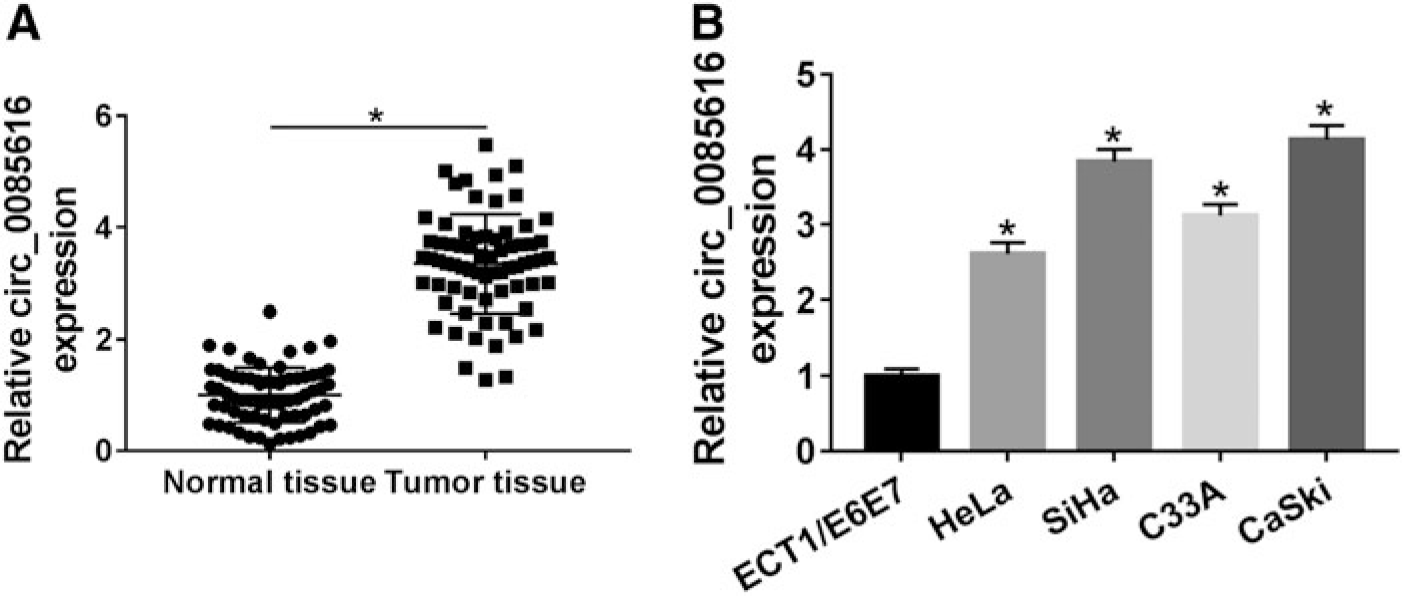
Circ_0085616 was dysregulated with a high level in CC tissues and cells.
Knockdown of circ_0085616 blocked CC cellular proliferation, metastasis, and glycolysis
After the specific siRNA targeting of circ_0085616 (si-circ_0085616#1, si-circ_0085616#2 and si-circ_0085616#3) to interfere with the circ_0085616 expression (Fig. 2A, B), we selected si-circ_0085616#2 with the best efficiency to perform the research in the cellular behaviors of SiHa and CaSki cells. As a consequence of circ_0085616 downregulation, cell proliferation was distinctly reduced in SiHa and CaSki cells through the MTT (Fig. 2C, D) and colony formation (Fig. 2E) assays. Also, the migrated (Fig. 2F) and invaded (Fig. 2G) cells in si-circ_0085616#2-transfected SiHa and CaSki cells were fewer than those in si-NC-transfected cells. Glycolysis is a well-known metabolic pathway with the consumption of glucose and the production of lactate and ATP, and it is meanwhile accompanied by the involvement of abundant enzymes. 18 After circ_0085616 expression was hampered, we found there were inhibitory effects on glucose consumption (Fig. 2H), lactate production (Fig. 2I), and ATP production (Fig. 2J) in SiHa and CaSki cells. Western blot exhibited the downregulation of Ki67 and PCNA (pro-proliferation markers), Vimentin (epithelial–mesenchymal transition [EMT]-promoting marker) inhibition, and E-cadherin (EMT-inhibitory marker) upregulation; the decrease of LDHA and GLUT1 (glycolytic enzymes) (Fig. 2K). These results affirmed that knockdown of circ_0085616 obstructed the cancerous process of CC.

Knockdown of circ_0085616 blocked CC cellular proliferation, metastasis, and glycolysis.
Circ_0085616 generated the sponge effect on miR-503-5p
Through the Venn diagram analysis of miRNA targets of circ_0085616 predicted by three online softwares (circinteractome, circBank, starbase 3.0), miR-503-5p was only mutual miRNA among three softwares (Fig. 3A). MiR-503-5p was then used for the target analysis of circ_0085616 in this study. The binding sites of miR-503-5p were shown in the sequence of circ_0085616 in circinteractome, as depicted in Figure 3B. Subsequently, the binding between circ_0085616 and miR-503-5p was affirmed by dual-luciferase reporter assay and RIP assay. In SiHa and CaSki cells that overexpressed miR-503-5p, the luciferase activity of circ_0085616-WT vector was declined whereas that of circ_0085616-MUT had no significant change (Fig. 3C, D). Significantly, circ_0085616 and miR-503-5p were largely enriched in the Anti-Ago2 group compared with the Anti-IgG group (Fig. 3E), indicating the interaction between circ_0085616 and miR-503-5p in CC cells. Transfection of si-circ_0085616#2 caused the promotive effect on the level of miR-503-5p, relative to si-NC transfection (Fig. 3F). The qRT-PCR also validated the downregulation of miR-503-5p in CC, including tissues and cells (SiHa and CaSki) (Fig. 3G, H). Hence, circ_0085616 was able to sponge miR-503-5p.
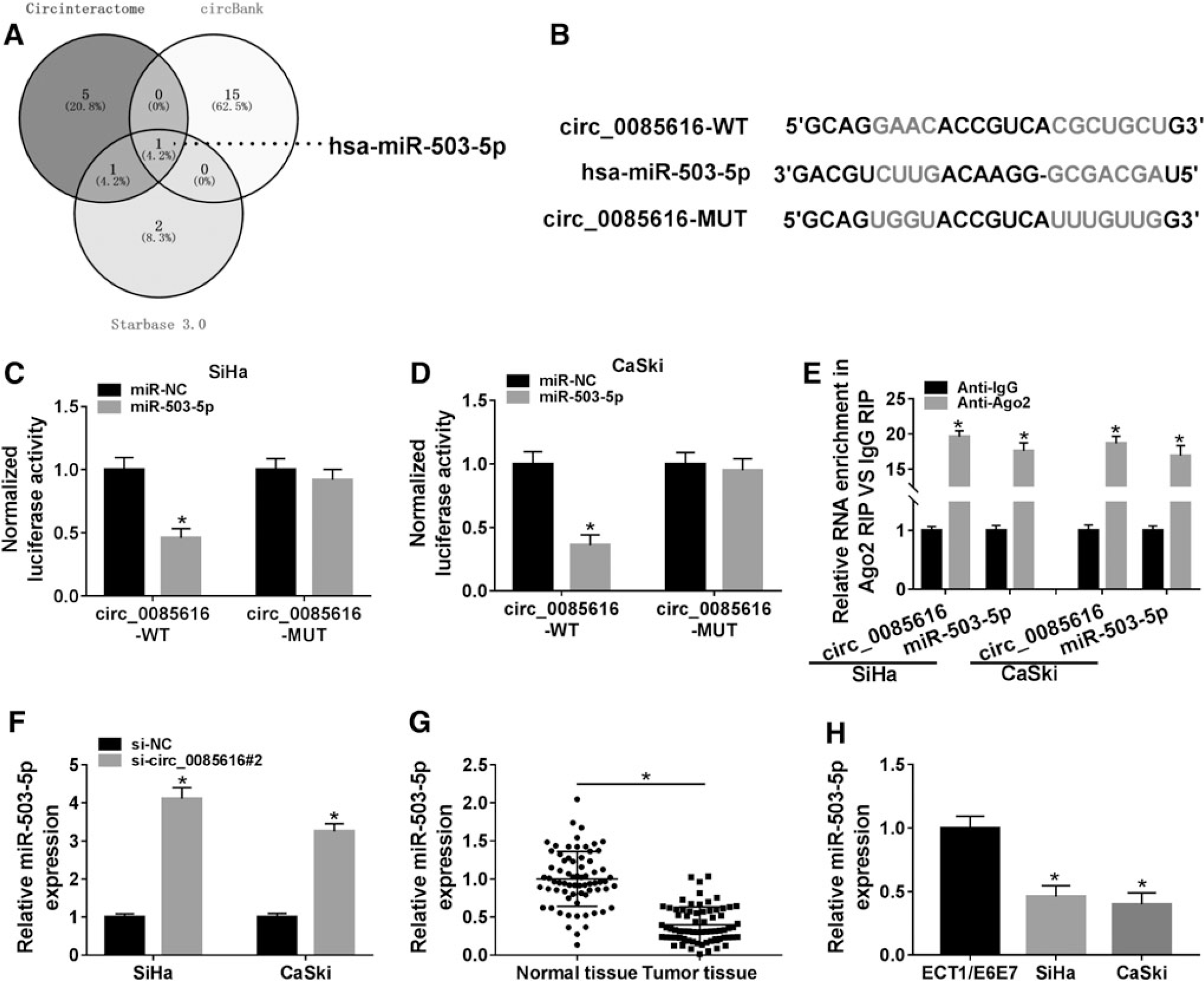
Circ_0085616 generated the sponge effect on miR-503-5p.
Inhibition of miR-503-5p returned the effects of circ_0085616 knockdown on CC cells
Based on the effective inhibition of miR-503-5p by anti-miR-503-5p transfection (Fig. 4A, B), the rescued transfection of anti-miR-503-5p to si-circ_0085616#2 was performed in SiHa and CaSki cells. Subsequent cellular analyses demonstrated that the si-circ_0085616#2-induced suppressive effects on proliferation (Fig. 4C–E), metastasis (Fig. 4F, G), and glycolytic metabolism (Fig. 4H–J) in SiHa and CaSki cells were all reversed by downregulating miR-503-5p. The reversal of anti-miR-503-5p to si-circ_0085616#2 on the protein changes of proliferation, metastasis, and glycolysis-associated markers (Fig. 4K) also implied that miR-503-5p inhibition neutralized the effects on CC cellular processes evoked by circ_0085616 knockdown.
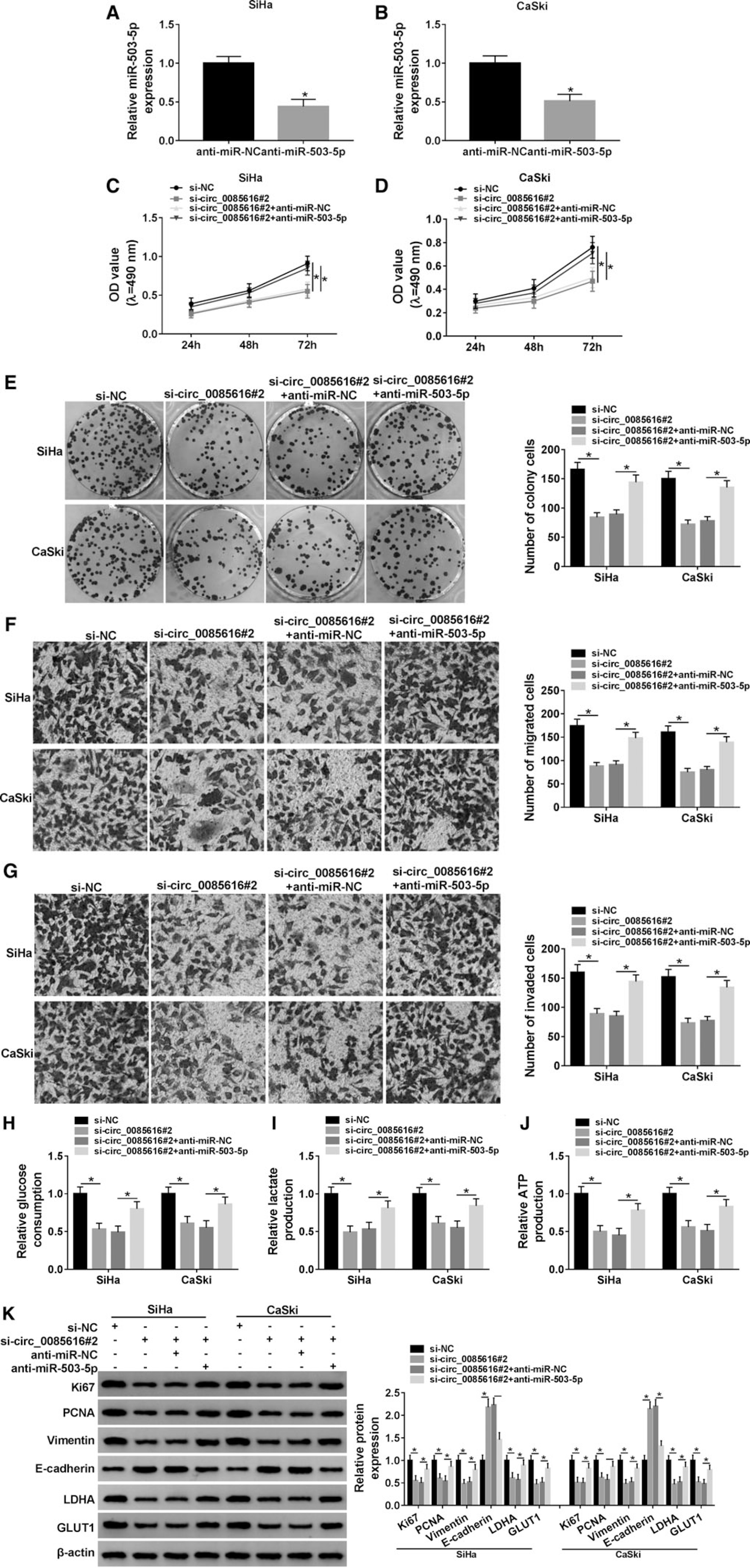
Inhibition of miR-503-5p returned the effects of circ_0085616 knockdown on CC cells.
MiR-503-5p directly targeted ATXN7L3
Regarding the molecular target of miR-503-5p, we detected the expression of 7 genes (RAB10, SOX4, ZNF217, IGF1R, LETM1, BACP31, ATXN7L3) predicted by starbase 3.0 after transfection of miR-NC or miR-503-5p. As shown in Figure 5A, the downregulated effect of miR-503-5p overexpression on ATXN7L3 was the most significant in SiHa cells. Thus, ATXN7L3 was considered a candidate target of miR-503-5p in CC during this study. Starbase 3.0 presented the binding sites between miR-503-5p and ATXN7L3 3′UTR (Fig. 5B). Dual-luciferase reporter results revealed that miR-503-5p combined with ATXN7L3 3′UTR to inhibit the luciferase intensity of the WT group, not the MUT group (Fig. 5C, D). In contrast with miR-NC transfection, ATXN7L3 protein expression was notably decreased after miR-503-5p transfection in SiHa and CaSki cells, showing that miR-503-5p directly inhibited ATXN7L3 level in CC cells (Fig. 5E). Besides, both CC tissues (Fig. 5F, G) and cells (Fig. 5H, I) displayed the overexpression of ATXN7L3 mRNA and protein levels that are different from normal tissues and ECT1/E6E7 cells. Altogether, miR-503-5p was shown to target ATXN7L3 directly.

MiR-503-5p directly targeted ATXN7L3.
Overexpression of miR-503-5p inhibited the proliferation, metastasis, and glycolysis of CC cells by decreasing ATXN7L3 expression
Through the analysis of Western blot, as depicted in Figure 6A and B, ATXN7L3 protein level was successfully increased by twofold after the introduction of pcDNA-ATXN7L3. Then, SiHa and CaSki cells were transfected with miR-NC, miR-503-5p, miR-503-5p+pcDNA, or miR-503-5p+pcDNA-ATXN7L3 to analyze the regulatory relationship between miR-503-5p and ATXN7L3 on different biological processes. Prominently, miR-503-5p acted as a tumor inhibitor to repress the proliferation (Fig. 6C–E), migration (Fig. 6F), and invasion (Fig. 6G) in SiHa and CaSki cells but ATXN7L3 upregulation eliminated these effects. It was similar that miR-503-5p played an inhibitory role in the process of glycolysis (repression of glucose consumption, lactate, and ATP production), which was counterbalanced after the addition of pcDNA-ATXN7L3 (Fig. 6H–J). By performing the protein detection using Western blot (Fig. 6K), the results again clarified that miR-503-5p targeted ATXN7L3 to hinder cell proliferation (downregulation of Ki67 and PCNA), metastasis (Vimentin inhibition and E-cadherin upregulation), and glycolysis (decline of LDHA and GLUT1) in CC cells.
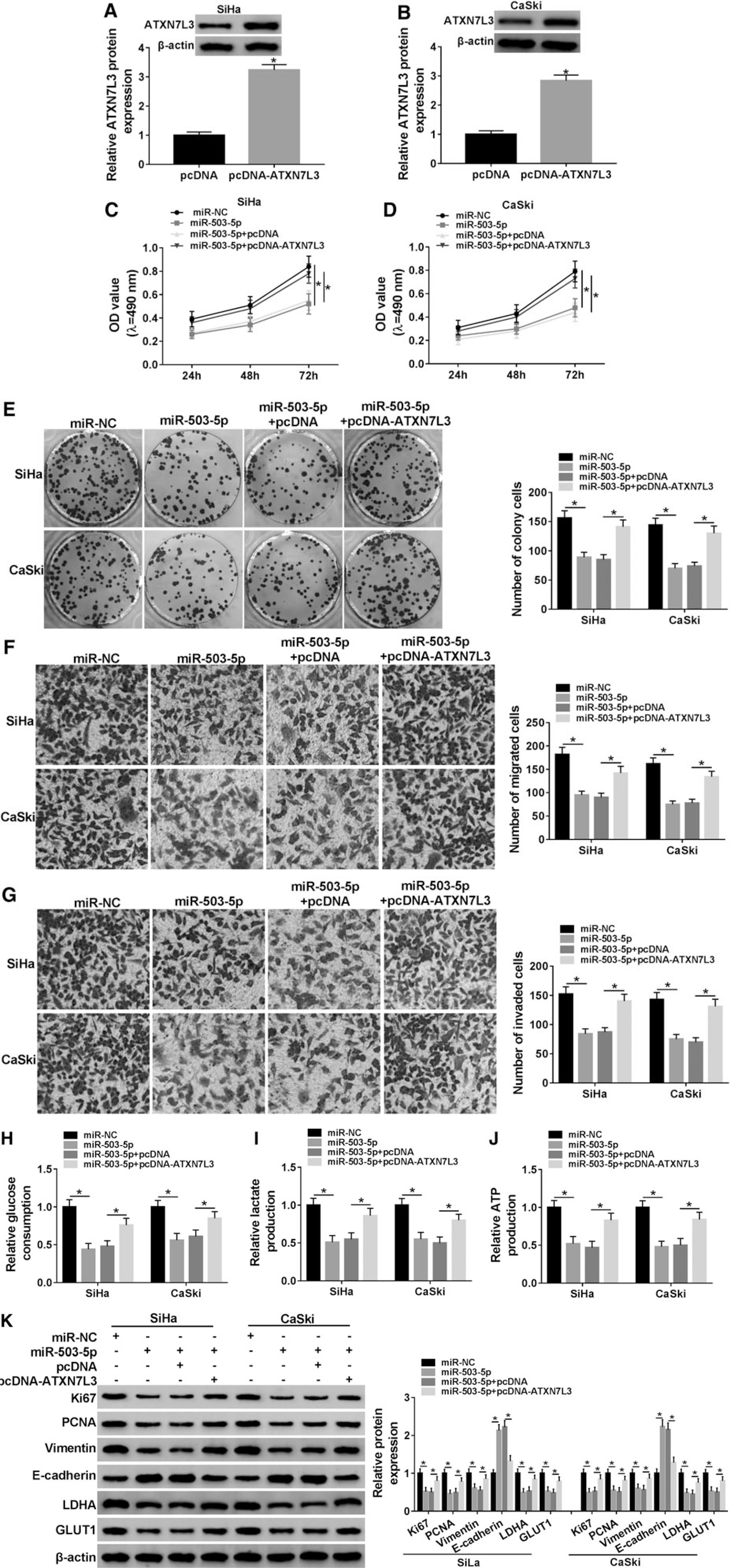
Overexpression of miR-503-5p inhibited the proliferation, metastasis, and glycolysis of CC cells by decreasing ATXN7L3 expression.
Circ_0085616 elevated ATXN7L3 via sponging miR-503-5p
Interestingly, the protein expression of ATXN7L3 was restrained by transfection of si-circ_0085616#2, whereas ATXN7L3 was then increased in the si-circ_0085616#2+anti-miR-503-5p group in contrast with the si-circ_0085616#2+anti-miR-NC group (Fig. 7A). Spearman's correlation coefficient suggested that there was a significantly negative relationship between the expression of circ_0085616 and miR-503-5p (r = −0.414, p = 0.0004) (Fig. 7B), as well as between miR-503-5p and ATXN7L3 (r = −0.455, p < 0.0001) (Fig. 7C). However, circ_0085616 was positively correlated to ATXN7L3 (r = 0.371, p = 0.0016) (Fig. 7D). These data indicated the positive regulation of circ_0085616 on ATXN7L3 via sponging miR-503-5p.

Circ_0085616 elevated ATXN7L3 via sponging miR-503-5p.
Circ_0085616 promoted tumorigenesis of CC in vivo by the mediation of miR-503-5p/ATXN7L3 axis
Through the animal experiment in vivo, tumor volume (Fig. 8A) and weight (Fig. 8B) were shown to be reduced by sh-circ_0085616, in contrast to sh-NC. Regarding the expression of three molecules, sh-circ_0085616 overtly suppressed the circ_0085616 level (Fig. 8C), enhanced miR-503-5p expression (Fig. 8D), and downregulated ATXN7L3 protein level (Fig. 8E). The sh-circ_0085616-mediated CC growth inhibition was achieved through increasing miR-503-5p to inhibit ATXN7L3, implicating that circ_0085616 promoted CC tumorigenesis in vivo by modulating the miR-503-5p/ATXN7L3 axis.
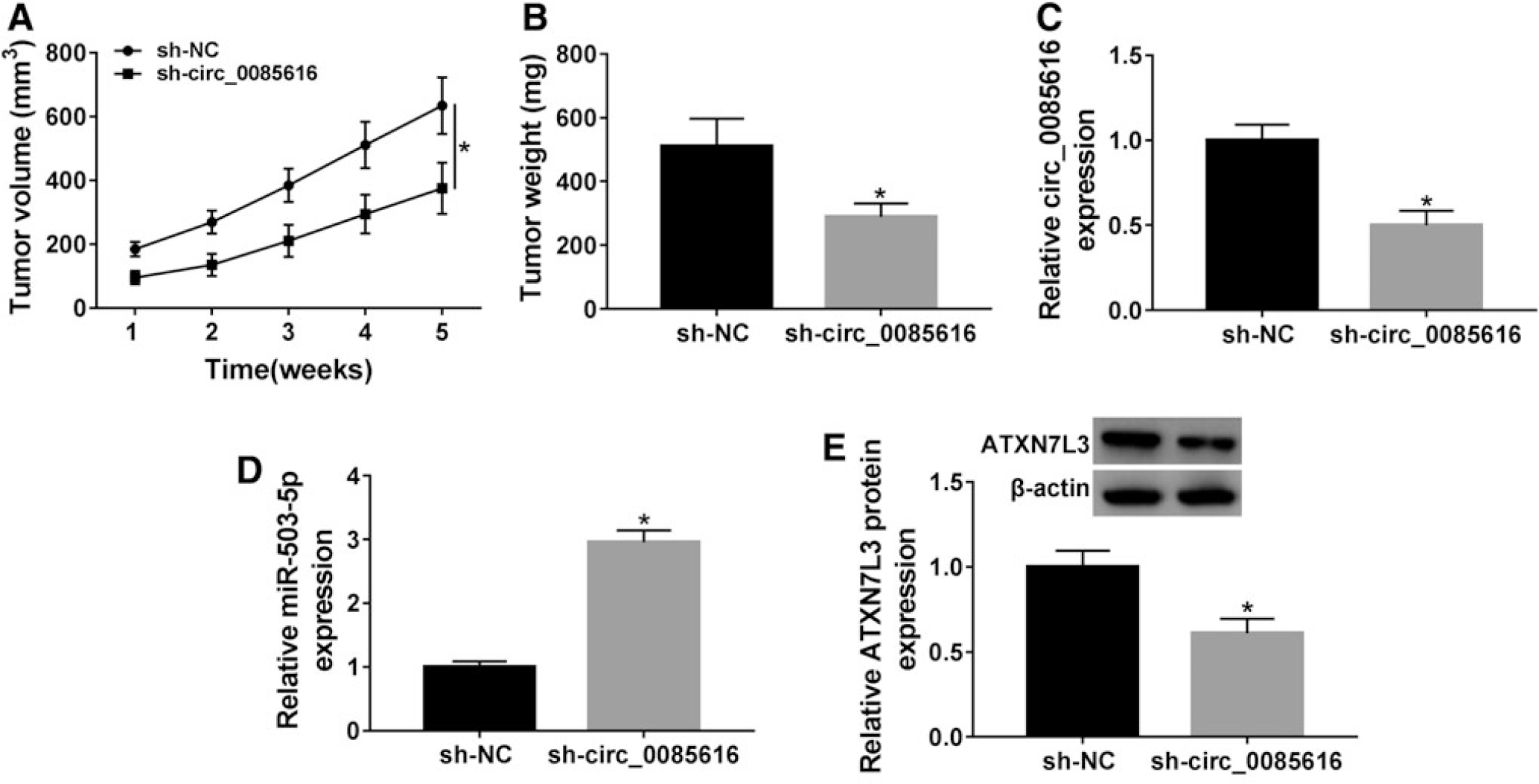
Circ_0085616 promoted tumorigenesis of CC in vivo by the mediation of miR-503-5p/ATXN7L3 axis.
Discussion
With the exploration of the roles of circRNAs in CC, a multitude of novel biomarkers related to carcinogenesis have been exposed in recent years. 19 In this chapter, circ_0085616 was confirmed to be upregulated in CC tissues and different CC cell lines. Then, we further studied the function of circ_0085616 via knocking down its expression, and we illuminated its action mechanism in detail.
As a product of nonclassical backsplicing in pre-mRNA, circRNAs are shaped as circular single-stranded RNA molecules that do not possess the 5′ caps and 3′ poly (A) tails. 20 Various biological properties of circRNAs have been recognized according to the research evidence, including widespread distribution, high resistance to RNase R digestion, high conservation, etc. 21 CircRNAs can function as either oncogenic stimuli or tumor inhibitors in multiple neoplasms. 22 Similar to procarcinomatous function, overexpression of circFARSA was shown to generate significant motivation on cell migration and invasion in nonsmall cell lung cancer 23 ; circRHOT1 was upregulated and accelerated the abilities of proliferation and metastasis in pancreatic cancer cells 24 ; and circBANP was affirmed as an upregulated circRNA that heightened the proliferative potential in colorectal cancer cells. 25 Regarding the antitumorous effect, circRNA profiling has identified circADAMTS13 as a proliferation repressor in hepatocellular carcinoma 26 ; circ_0004491 was proved to be a metastatic factor in oral squamous cell carcinoma cells 27 ; and circ_0001368 played a tumor-inhibitory role in gastric cancer to weaken cell viability and invasion. 28
As for circ_0085616 in CC, the qRT-PCR analysis in CC tissues and cells indicated its ectopic high expression. Li et al. have suggested the stimulatory role of circ_0085616 in cell proliferation, migration, and invasion of hepatocellular carcinoma. 29 Our results were in accordance with the earlier study in exhibiting the inhibitory phenomena in CC cell proliferation, migration, and invasion after the downregulation of circ_0085616. Moreover, aerobic glycolysis usually happens in tumors as one of the earliest known evidences in metabolic changes. 30 Dysregulated circRNAs can affect glycolysis in most cancers, such as melanoma, 31 breast cancer, 32 hepatocellular carcinoma, 33 and so on. After our detection of glycolytic indicators and enzymes, we validated si-circ_0085616 as a glycolytic holdback in CC cells to decelerate the cellular metabolism. Thus, circ_0085616 was a tumorigenic molecule in CC and its suppression may be another optional therapeutic approach against the deterioration of CC.
In participating in the carcinogenic action and malignant behavior of cancers, the “miRNA sponge” function is embodied in circRNA biology to affect the transcription and expression of genes. 34 Circ_LARP4 suppressed cell proliferative and invasive abilities in gastric cancer as a sponge of miR-424-5p to upregulate the level of LATS1. 35 Cao et al. announced that circRNF20 elevated the tumorigenesis and glycolytic effect in breast cancer cell by regulating HIF-1α and HK2 via sponging miR-487a. 36 Based on the target regulation of miR-503-5p by circ_0085616 and ATXN7L3 by miR-503-5p here, we subsequently found the modulation of circ_0085616 on ATXN7L3 via the sponge action on miR-503-5p. Our experiments suggested that circ_0085616 exerted the oncogenic properties on CC cellular progression in vitro and tumor growth in vivo via activating ATXN7L3 as a miR-503-5p sponge.
Conclusion
Overall, we first identified circ_0085616 as a cancerogenic molecule in regulating CC cell proliferation, metastasis, and glycolysis depending on the regulation of miR-503-5p and ATXN7L3, which exhibited an innovative signal network of circ_0085616/miR-503-5p/ATXN7L3 in CC progression. This finding may lay the advantageous foundation for circ_0085616 as an effective target in the diagnosis and treatment of CC.
Footnotes
Funding Information
No funding was received for this article.
Disclosure Statement
No competing financial interests exist.